
Non-Additive Effects of Mixing Eucalyptus and Castanopsis hystrix Trees on Carbon Stocks under an Eco-Silviculture Regime in Southern China

Abstract
:1. Introduction
2. Materials and Methods
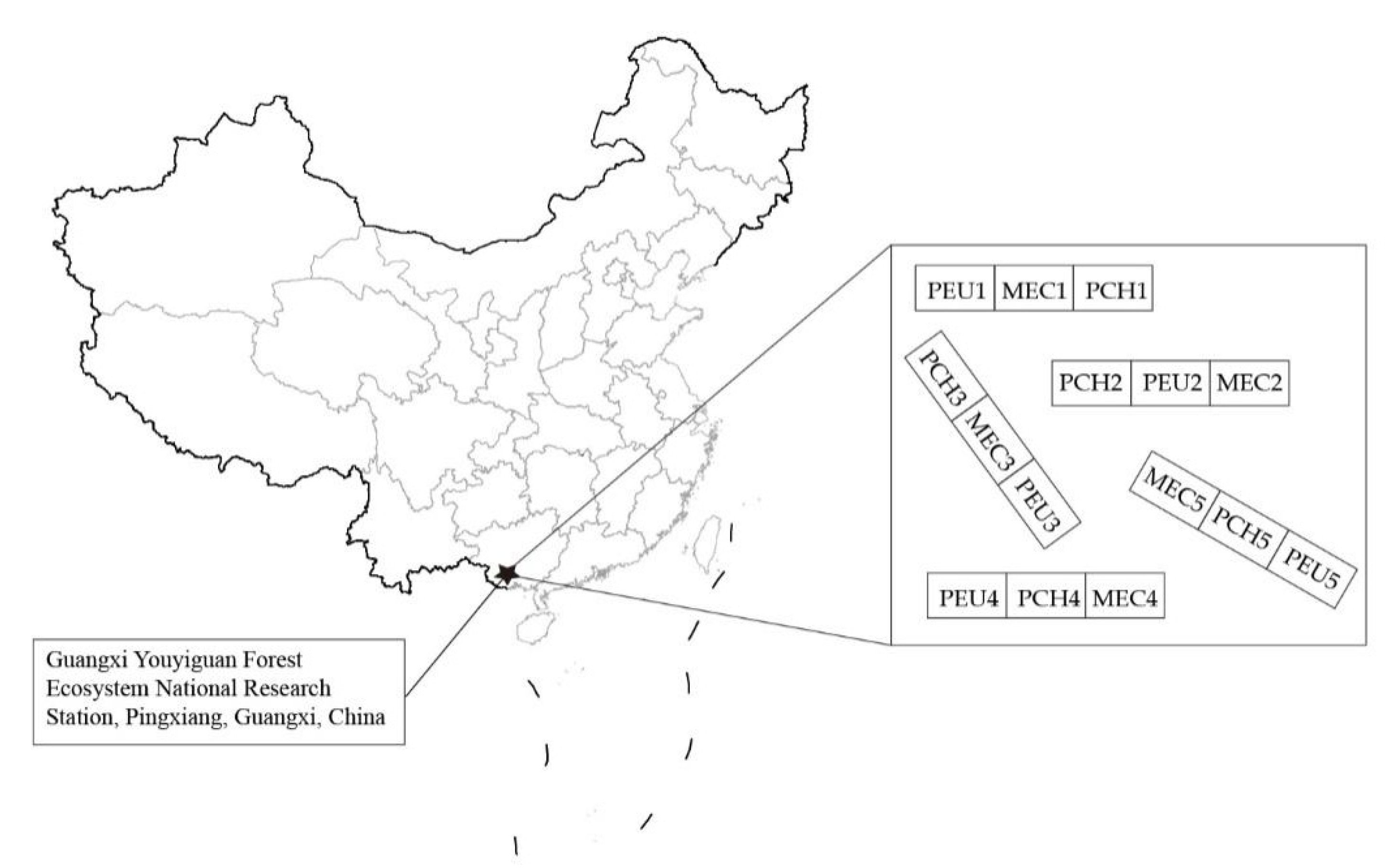
2.1. Study Area and Experimental Design
2.2. Overstory Survey and Timber Production Estimates
2.3. Overstory and Understory Biomass Measurements
2.4. Measuring the Carbon Content of Plant and Soil Components
2.5. Carbon Storage Measurements
2.6. Statistical Analysis
3. Results
3.1. Tree Growth
3.2. Carbon Content of Tree, Understory, Litter, and Soil
3.3. Carbon Storage
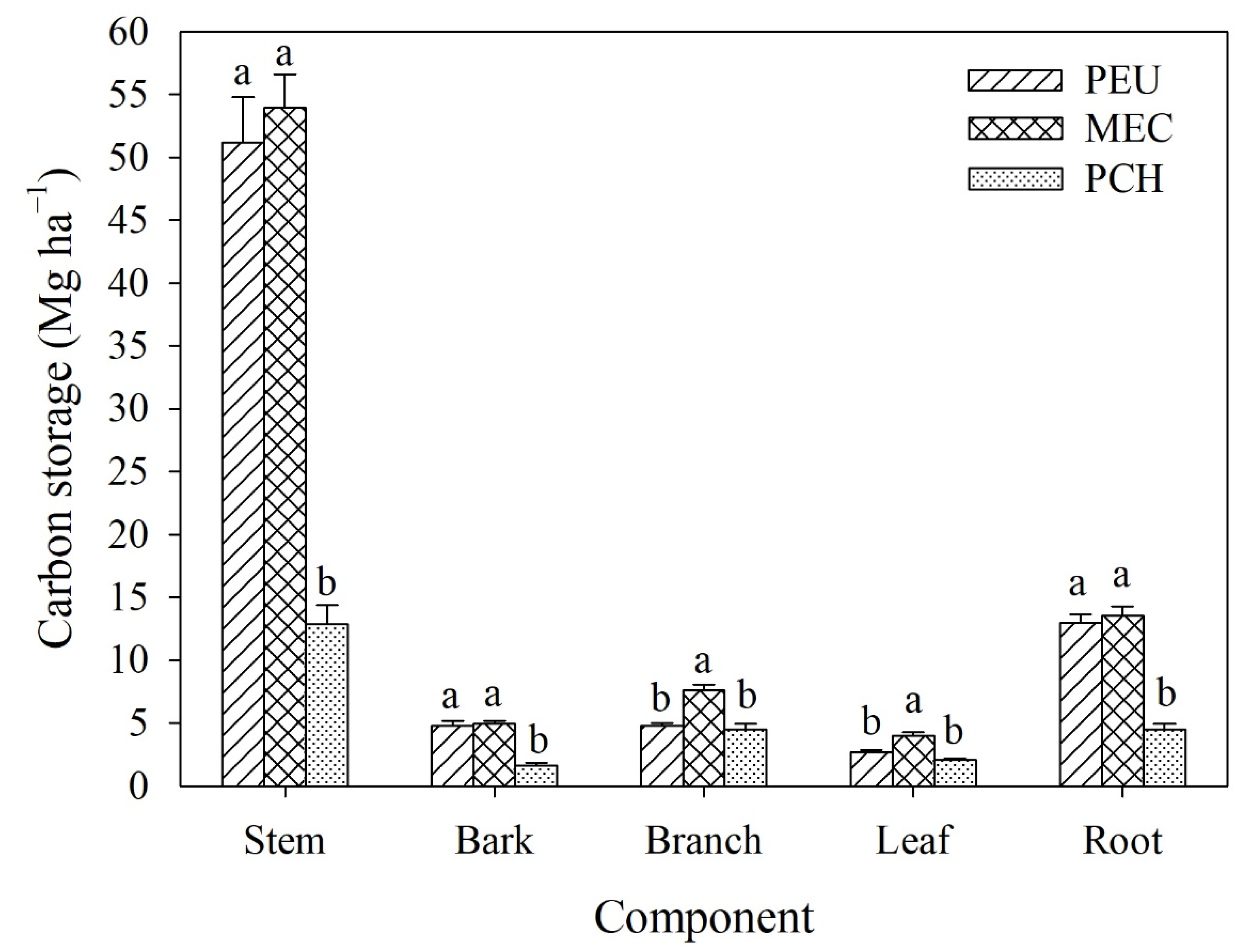
3.3.1. Tree Carbon Storage
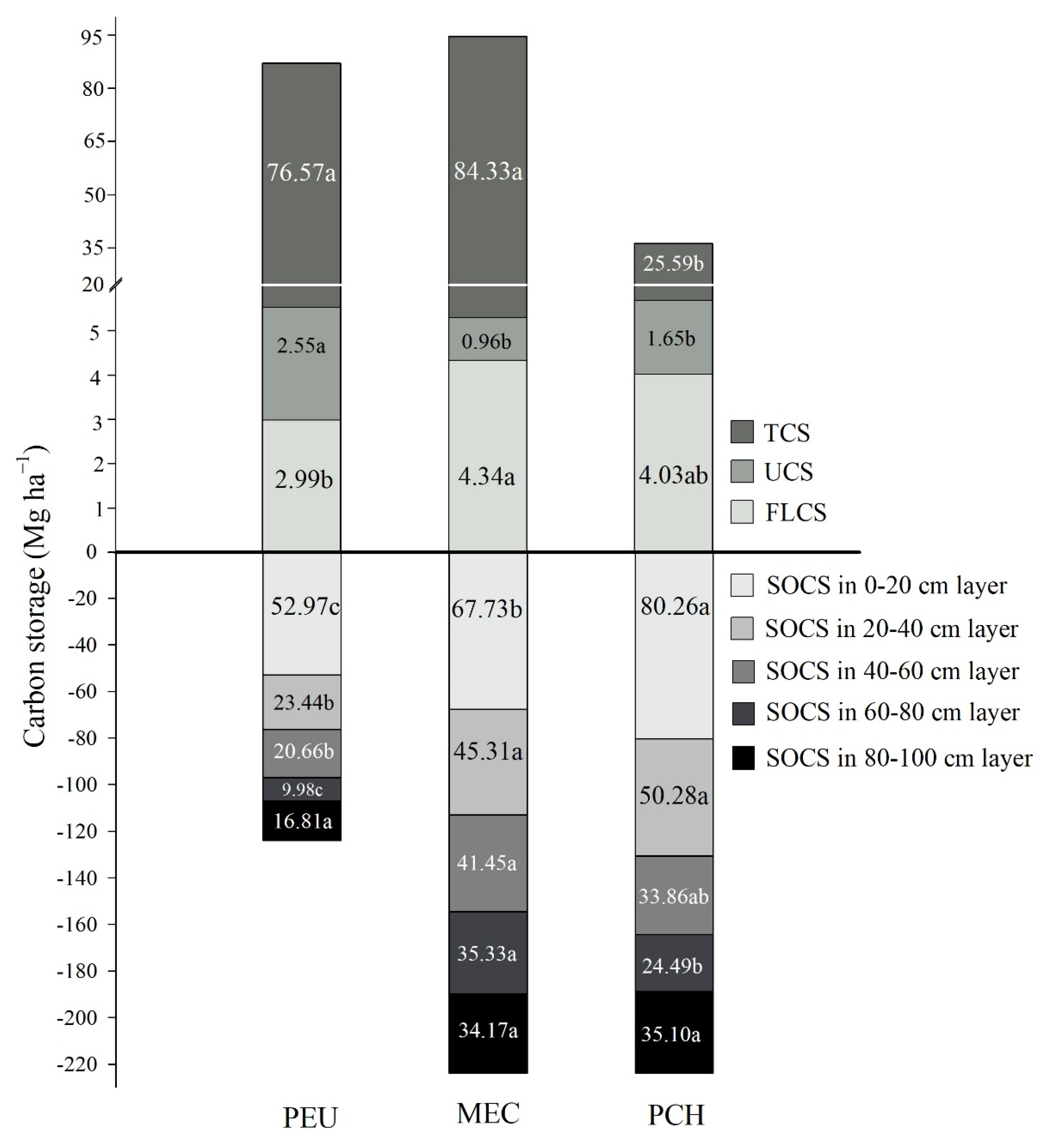
3.3.2. Understory and Floor Litter Carbon Storage
3.3.3. Soil Carbon Storage
3.3.4. Whole Ecosystem Carbon Storage
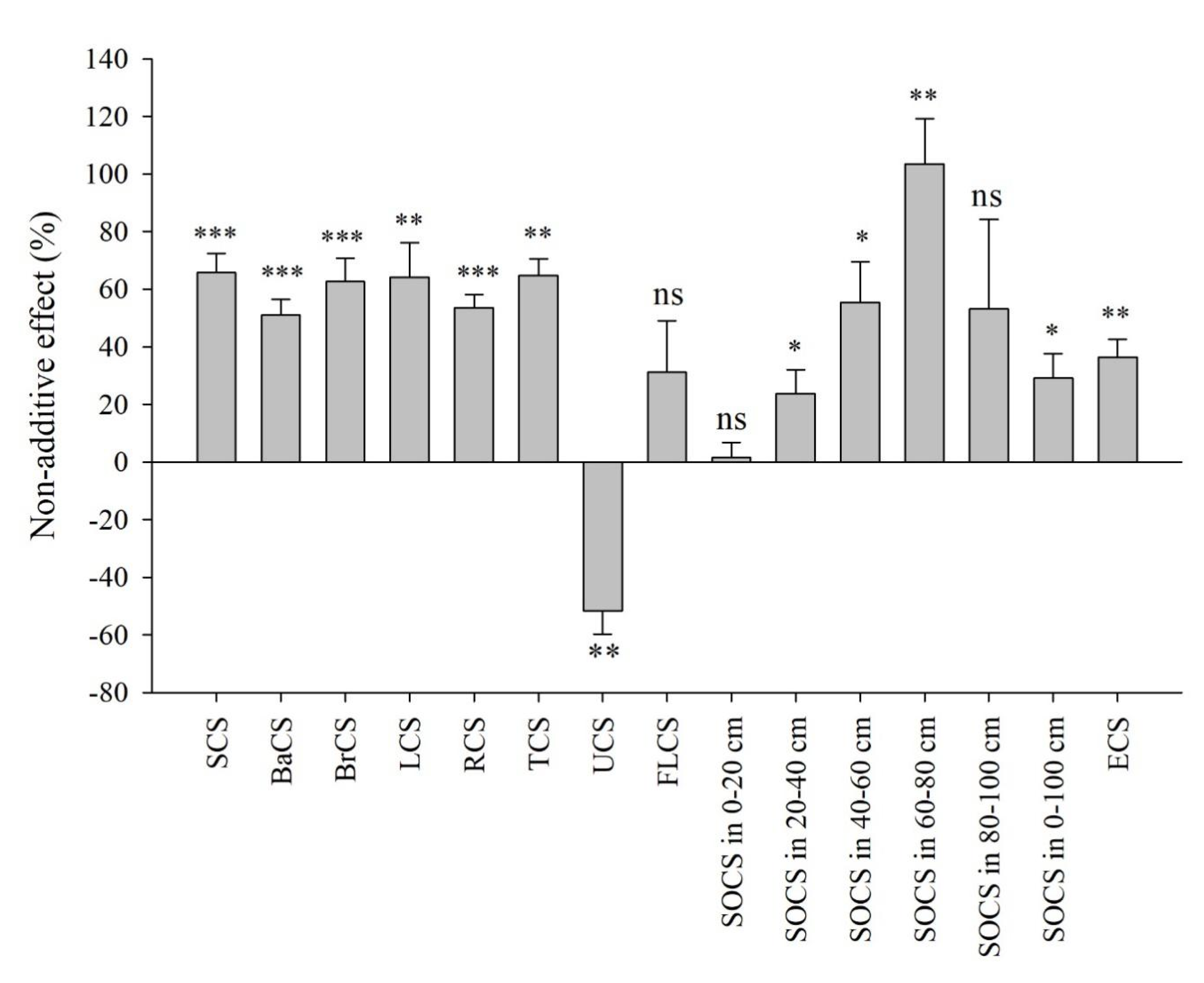
3.4. Admixing Effects on Whole Ecosystem Carbon Storage
4. Discussion
4.1. Effects of Mixing Species on Tree Growth and Productivity
4.2. Effect of Plantation Types on Whole Ecosystem Carbon Stocks
4.3. Tree Species Mixture Effects on Whole Ecosystem Carbon Stocks
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Global Forest Resources Assessment; Main Report; FAO: Rome, Italy, 2020. [Google Scholar]
- Huang, X.M.; Liu, S.R.; Wang, H.; Hu, Z.D.; Li, Z.G.; You, Y.M. Changes of soil microbial biomass carbon and community composition through mixing nitrogen-fixing species with Eucalyptus Urophylla in subtropical China. Soil Biol. Biochem. 2014, 73, 42–48. [Google Scholar] [CrossRef]
- Wen, Y.G.; Zhou, X.G.; Zhu, H.G. Theory, Technology and Practices in Eucalyptus Plantations under Eco-Silviculture Regime; Science Press: Beijing, China, 2020; pp. 1–254. (In Chinese) [Google Scholar]
- Zhou, X.G.; Zhu, H.G.; Wen, Y.G.; Goodale, U.M.; Zhu, Y.L.; Yu, S.F.; Li, C.T.; Li, X.Q. Intensive management and declines in soil nutrients lead to serious exotic plant invasion in Eucalyptus plantations under successive short–rotation regimes. Land Degrad. Dev. 2020, 31, 297–310. [Google Scholar] [CrossRef]
- Wu, J.P.; Fan, H.B.; Liu, W.F.; Huang, G.M.; Tang, J.F.; Zeng, R.J.; Huang, J.; Liu, Z.F. Should exotic Eucalyptus be planted in Subtropical China: Insights from understory plant diversity in two contrasting Eucalyptus chronosequences. Environ. Manag. 2015, 56, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- FAO. Global Forest Resources Assessment; Main Report; FAO: Rome, Italy, 2010. [Google Scholar]
- Williams, R.A. Mitigating biodiversity concerns in Eucalyptus plantations located in South China. J. Biosci. Med. 2015, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, J.W. Eucalypt plantations. New For. 1999, 17, 37–52. [Google Scholar] [CrossRef]
- Zaiton, S.; Sheriza, M.R.; Ainishifaa, R.; Alfred, K.; Norfaryant, K. Eucalyptus in Malaysia: Review on environmental impacts. J. Landsc. Ecol. 2020, 13, 79–94. [Google Scholar] [CrossRef]
- Zhang, K.; Zheng, H.; Chen, F.L.; Ouyang, Z.Y.; Wang, Y.; Wu, Y.F.; Lan, L.; Fu, M.; Xiang, X.W. Changes in soil quality after converting Pinus to Eucalyptus plantations in southern China. Solid Earth 2015, 6, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.G.; Ye, D.; Chen, F.; Liu, S.R.; Liang, H.W. The changes of understory plant diversity in continuous cropping system of Eucalyptus plantations, South China. J. For. Res. 2010, 15, 252–258. [Google Scholar] [CrossRef]
- Zhou, X.G.; Ye, D.; Zhu, H.G.; Li, X.Q.; Su, Y.; Lan, J.; Wen, Y.G. Effects of second rotation seedlings and coppice on understorey vegetation and timber production of Eucalyptus plantations. J. Trop. For. Sci. 2017, 29, 54–68. [Google Scholar]
- Jin, D.; Huang, Y.; Zhou, X.L.; Chen, B.; Ma, J.; Yan, Y.H. High risk of plant invasion in the understory of eucalypt plantations in South China. Sci. Rep. 2015, 5, 18492. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.G.; Zhu, H.G.; Wen, Y.G.; Goodale, U.M.; Li, X.Q.; You, Y.M.; Ye, D.; Liang, H.W. Effects of understory management on trade-offs and synergies between biomass carbon stock, plant diversity and timber production in eucalyptus plantations. For. Ecol. Manag. 2018, 410, 164–173. [Google Scholar] [CrossRef]
- Li, X.Q.; Ye, D.; Liang, H.W.; Zhu, H.G.; Qin, L.; Zhu, Y.L.; Wen, Y.G. Effects of successive rotation regimes on carbon stocks in Eucalyptus plantations in subtropical China measured over a full rotation. PLoS ONE 2015, 10, e0132858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelty, M.J. Comparative productivity of monocultures and mixed-species stands. In The Ecology and Silviculture of Mixed-Species Forests; Kelty, M.J., Ed.; Kluwer Academic: Dordrecht, The Netherlands, 1992; pp. 25–141. [Google Scholar]
- Man, R.Z.; Lieffers, V.J. Are mixtures of aspen and white spruce more productive than single species stands? For. Chron. 1999, 75, 505–513. [Google Scholar] [CrossRef]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Guo, L.B.; Sims, R.E.H. Eucalypt litter decomposition and nutrient release under a short rotation forest regime and effluent irrigation treatments in New Zealand. I. External effects. Soil Biol. Biochem. 2001, 33, 1381–1388. [Google Scholar] [CrossRef]
- McTiernan, K.B.; Ineson, P.; Coward, P.A. Respiration and nutrient release from tree leaf litter mixtures. Oikos 1997, 78, 527–538. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.R.; Wang, J.X.; You, Y.M.; Yang, J.Y.; Shi, Z.M.; Huang, X.M.; Zheng, L.; Li, Z.Y.; Ming, A.G.; et al. Mixed-species plantation with Pinus massoniana and Castanopsis hystrix accelerates C loss in recalcitrant coniferous litter but slows C loss in labile broadleaf litter in southern China. Forest Ecol. Manag. 2018, 422, 207–213. [Google Scholar] [CrossRef]
- Wen, Y.G.; Zhang, Z.F.; Zhou, X.G.; Zhu, H.G.; Wang, L.; Cai, D.X.; Jia, H.Y.; Ming, A.G.; Lu, L.H. Effects of mixing precious indigenous tree species and Eucalyptus on ecosystem biomass and carbon stocks. Guangxi Sci. 2020, 27, 111–119. (In Chinese) [Google Scholar]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L. Carbon allocation in a mixed-species plantation of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 2006, 233, 275–284. [Google Scholar] [CrossRef]
- He, Y.J.; Qin, L.; Li, Z.Y.; Liang, X.Y.; Shao, M.X.; Tan, L. Carbon storage capacity of monoculture and mixed-species plantations in subtropical China. For. Ecol. Manag. 2013, 295, 193–198. [Google Scholar] [CrossRef]
- Amazonasa, N.T.; Forrester, D.I.; Silva, C.C.; Almeida, D.R.A.; Rodrigues, R.R.; Brancalion, P.H.S. High diversity mixed plantations of Eucalyptus and native trees: An interface between production and restoration for the tropics. For. Ecol. Manag. 2018, 417, 247–256. [Google Scholar] [CrossRef]
- McCracken, A.R.; Dawson, W.M. Growing clonal mixtures of willow to reduce effect of Melampsora epitea var. epitea. Eur. J. For. Pathol. 1997, 27, 319–329. [Google Scholar] [CrossRef]
- Burdon, R.D. Genetic diversity and disease resistance: Some considerations for research, breeding, and deployment. Can. J. For. Res. 2001, 31, 596–606. [Google Scholar] [CrossRef]
- Rothe, A.; Binkley, D. Nutritional interactions in mixed species forests: A synthesis. Can. J. For. Res. 2001, 31, 1855–1870. [Google Scholar] [CrossRef]
- Piotto, D. A meta-analysis comparing tree growth in monocultures and mixed plantations. For. Ecol. Manag. 2008, 255, 781–786. [Google Scholar] [CrossRef]
- Binkley, D.; Stape, J.L.; Ryan, M.G. Thinking about efficiency of resource use in forests. For. Ecol. Manag. 2004, 193, 5–16. [Google Scholar] [CrossRef]
- Chomel, M.; DesRochers, A.; Baldy, V.; Larchevêque, M.; Gauquelin, T. Non-additive effects of mixing hybrid poplar and white spruce on aboveground and soil carbon storage in boreal plantations. For. Ecol. Manag. 2014, 328, 292–299. [Google Scholar] [CrossRef]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, S.R.; Schindlbacher, A.; Wang, J.X.; Yang, Y.J.; Song, Z.C.; You, Y.M.; Si, Z.M.; Chen, L.; Ming, A.G.; et al. Experimental warming reduced topsoil carbon content and increased soil bacterial diversity in a subtropical planted forest. Soil Biol. Biochem. 2019, 133, 155–164. [Google Scholar] [CrossRef]
- Meng, X.Y. Forest Mensuration; China Forestry Publishing House: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Liu, F.; Jiang, Y.; Dai, L.; Huang, R.L.; Wei, L.X.; Wang, Y. Growth rhythm and fitting effect of six kinds of growth model of Castanopsis hystrix. Guangxi For. Sci. 2014, 43, 264–270. (In Chinese) [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis: Part 2, Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 539–580. [Google Scholar]
- Wardle, D.A.; Bonner, K.I.; Nicholson, K.S. Biodiversity and plant litter: Experimental evidence which does not support the view that enhanced species richness improves ecosystem function. Oikos 1997, 79, 247–258. [Google Scholar] [CrossRef]
- Ming, A.G.; Liu, S.R.; Mo, H.H.; Cai, D.X.; Nong, Y.; Zeng, J.; Li, H.; Tao, Y. Comparison of carbon storage in pure and mixed stands of Castanopsis hystrix and Cunninghamia lanceolata in subtropical China. Acta Ecol. Sin. 2016, 36, 244–251. (In Chinese) [Google Scholar]
- Zhou, X.G.; Wen, Y.G.; Goodale, U.M.; Zuo, H.; Zhu, H.G.; Li, X.Q.; You, Y.M.; Yan, L.; Su, Y.; Huang, X.M. Optimal rotation length for carbon sequestration in Eucalyptus plantations in subtropical China. New For. 2017, 48, 609–627. [Google Scholar] [CrossRef]
- Kara, F.; Topaçoğlu, O. Influence of stand density and canopy structure on the germination and growth of Scots pine (Pinus sylvestris L.) seedlings. Environ. Monit. Assess. 2018, 190, 749. [Google Scholar] [CrossRef] [PubMed]
- Assenac, G. Interactions between forest stands and microclimate: Ecophysiological aspects and consequences for silviculture. Ann. Forest Sci. 2000, 57, 287–301. [Google Scholar] [CrossRef]
- Collet, C.; Lanter, O.; Pardos, M. Effects of canopy opening on height and diameter growth in naturally regenerated beech seedlings. Ann. For. Sci. 2001, 58, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Pardos, M.; Montes, F.; Aranda, I.; Cañellas, I. Influence of environmental conditions on germinant survival and diversity of Scots pine (Pinus sylvestris L.) in central Spain. Eur. J. For. Res. 2007, 126, 37–47. [Google Scholar] [CrossRef]
- Grams, T.E.; Andersen, C. Competition for resources in trees: Physiological versus morphological plasticity. In Progress in Botany; Esser, K., Löttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 356–381. [Google Scholar]
- Bauhus, J.; Aubin, I.; Messier, C.; Connell, M. Composition, structure, light attenuation and nutrient content of the understorey vegetation in a Eucalyptus sieberi regrowth stand 6 years after thinning and fertilisation. For. Ecol. Manag. 2001, 144, 275–286. [Google Scholar] [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Vandermeer, J. The Ecology of Intercropping; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Liu, L.L.; King, J.S.; Giardina, C.P. Effects of elevated concentrations of atmospheric CO2 and tropospheric O3 on leaf litter production and chemistry in trembling aspen and paper birch communities. Tree Physiol. 2005, 25, 1511–1522. [Google Scholar] [CrossRef]
- Sayer, E.J.; Heard, M.S.; Grant, H.K.; Marthews, T.R.; Tanner, E.V.J. Soil carbon release enhanced by increased tropical forest litterfall. Nat. Clim. Change 2011, 1, 304–307. [Google Scholar] [CrossRef]
- Leff, J.W.; Wieder, W.R.; Taylor, P.G.; Townsend, A.R.; Nemergut, D.R.; Grandy, A.S.; Cleveland, C.C. Experimental litterfall manipulation drives large and rapid changes in soil carbon cycling in a wet tropical forest. Glob. Change Biol. 2012, 18, 2969–2979. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.Z.; Li, Y.; Luo, X.Q.; Liu, Y.; Huang, P.; Bin, Y.B.; Zhang, L.Y.; Li, W.L.; Xue, J.M.; Gao, H.J.; et al. Mixed plantations enhance more soil organic carbon stocks than monocultures across China: Implication for optimizing afforestation/reforestation strategies. Sci. Total Environ. 2022, 821, 153449. [Google Scholar] [CrossRef] [PubMed]
- Benomar, L.; DesRochers, A.; Larocque, G.R. Comparing growth and fine root distribution in monocultures and mixed plantations of hybrid poplar and spruce. J. For. Res. 2013, 24, 247–254. [Google Scholar] [CrossRef]
- Wu, W.X.; Zhou, X.G.; Wen, Y.G.; Zhu, H.G.; You, Y.M.; Qin, Z.W.; Li, Y.C.; Huang, X.M.; Yan, L.; Li, X.Q. Coniferous-broadleaf mixture increases soil microbial biomass and functions accompanied by improved stand biomass and litter production in Subtropical China. Forests 2019, 10, 879. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.H.; Popadiouk, R.V. Dynamics of North American boreal mixedwoods. Environ. Rev. 2002, 10, 137–166. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Kirschbaum, M.U.; Hou, Z.H.; Guo, Z.H. Carbon stock changes in successive rotations of Chinese fir (Cunninghamia lanceolata (lamb) hook) plantations. For. Ecol. Manag. 2004, 202, 131–147. [Google Scholar] [CrossRef]
- Jouquet, P.; Huchet, G.; Bottinelli, N.; Thu, T.D.; Duc, T.T. Does the influence of earthworms on water infiltration, nitrogen leaching and soil respiration depend on the initial soil bulk density? A mesocosm experiment with the endogeic species Metaphire posthuma. Biol. Fertil. Soils 2012, 48, 561–567. [Google Scholar] [CrossRef]
- Lu, L.H.; Jia, H.J.; He, R.M.; Li, J.L.; Qin, S.Y. A preliminary study on litter falls of six kinds of plantations in the tropical South Asia. For. Res. 2008, 21, 346–352. [Google Scholar]
- Bonanomi, G.; Incerti, G.; Antignani, V.; Capodilupo, M.; Mazzoleni, S. Decomposition and nutrient dynamics in mixed litter of Mediterranean species. Plant Soil. 2010, 331, 481–496. [Google Scholar] [CrossRef]
- Barantal, S.; Roy, J.; Fromin, N.; Schimann, H.; Hättenschwiler, S. Long-term presence of tree species but not chemical diversity affect litter mixture effects on decomposition in a neotropical rainforest. Oecologia 2011, 167, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Cornelissen, J.H.C.; Zhang, Q.Q.; Yan, E.R. Functional evenness of N-to-P ratios of evergreen-deciduous mixtures predicts positive non-additive effect on leaf litter decomposition. Plant Soil 2019, 436, 299–309. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, G.Y.; Cao, G.M.; Zhang, X.X.; Liu, Z.W. Non-additive effects of leaf litter mixtures from Robinia pseudoacacia and ten tree species on soil properties. J. Sustain. For. 2020, 39, 1–14. [Google Scholar] [CrossRef]
- MacPherson, D.M.; Lieffers, V.J.; Blenis, P.V. Productivity of aspen stands with and without a spruce understory in Alberta’s boreal mixedwood forests. For. Chronicle. 2001, 77, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.L.; Gower, S.T.; Plaut, J.; Holmes, B. Carbon pools in a boreal mixedwood logging chronosequence. Glob. Change Biol. 2005, 11, 1883–1894. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Abbreviation | Full Name | Unit |
|---|---|---|---|
| Plantation type | PEU | pure Eucalyptus plantation | |
| PCH | pure C. hystrix plantation | ||
| MEC | mixed Eucalyptus and C. hystrix plantation | ||
| Soil property | SBD | soil bulk density | g cm−3 |
| SWC | soil water content | % | |
| SOC | soil organic carbon | g kg−1 | |
| TN | soil total nitrogen | g kg−1 | |
| TP | soil total phosphorus | g kg−1 | |
| AN | soil available nitrogen | mg kg−1 | |
| AP | soil available phosphorus | mg kg−1 | |
| AK | soil available potassium | mg kg−1 | |
| DOC | soil dissolved organic carbon | mg kg−1 | |
| DON | soil dissolved organic nitrogen | mg kg−1 | |
| Tree characteristic | DBH | diameter at breast height | cm |
| H | tree height | m | |
| Carbon storage | SCS | stem carbon storage | Mg ha−1 |
| BaCS | bark carbon storage | Mg ha−1 | |
| BrCS | branch carbon storage | Mg ha−1 | |
| LCS | leaf carbon storage | Mg ha−1 | |
| RCS | root carbon storage | Mg ha−1 | |
| TCS | tree carbon storage | Mg ha−1 | |
| UCS | understory carbon storage | Mg ha−1 | |
| FLCS | floor litter carbon storage | Mg ha−1 | |
| SOCS | soil organic carbon storage | Mg ha−1 | |
| ECS | ecosystem carbon storage | Mg ha−1 |
| Content | Factor | PEU | PCH | MEC | F(4,12) | p |
|---|---|---|---|---|---|---|
| Topographic features | Longitude | 22°10′4.62″ N | 22°9′59.7″ N | 22°9′46.8″ N | ||
| Latitude | 106°41′39.8″ E | 106°41′41.7″ E | 106°41′48.5″ E | |||
| Elevation (m) | 253 | 242 | 257 | |||
| Slope aspect | Northwest | North | Northwest | |||
| Slope gradient (°) | 19 | 20 | 20 | |||
| Soil properties | Soil type | Latosol | Latosol | Latosol | ||
| pH | 4.50 ± 0.17 | 4.47 ± 0.11 | 4.51 ± 0.09 | 0.19 | 0.831 | |
| SBD (g cm−3) | 1.21 ± 0.02 | 1.19 ± 0.06 | 1.19 ± 0.01 | 0.93 | 0.422 | |
| SWC (%) | 32.16 ± 3.52 | 32.93 ± 3.07 | 32.83 ± 3.12 | 0.29 | 0.756 | |
| SOC (g kg−1) | 21.57 ± 1.42 | 22.03 ± 2.54 | 21.96 ± 1.78 | 1.99 | 0.179 | |
| TN (g kg−1) | 1.23 ± 0.15 | 1.24 ± 0.08 | 1.25 ± 0.05 | 0.76 | 0.490 | |
| TP (g kg−1) | 0.36 ± 0.08 | 0.34 ± 0.11 | 0.35 ± 0.07 | 3.33 | 0.071 | |
| AN (mg kg−1) | 15.27 ± 1.23 | 14.90 ± 1.14 | 14.64 ± 1.25 | 1.48 | 0.266 | |
| AP (mg kg−1) | 35.62 ± 3.53 | 37.23 ± 3.36 | 36.52 ± 3.88 | 0.47 | 0.636 | |
| AK (mg kg−1) | 64.18 ± 6.55 | 63.78 ± 5.21 | 64.93 ± 3.42 | 1.86 | 0.197 | |
| DOC (mg kg−1) | 348.56 ± 15.24 | 340.21 ± 13.53 | 342.35 ± 11.62 | 0.90 | 0.431 | |
| DON (mg kg−1) | 42.48 ± 3.04 | 44.65 ± 3.19 | 43.16 ± 2.11 | 1.77 | 0.212 | |
| Microbial community PLFAs | Total PLFAs (nmol g−1) | 50.42 ± 3.86 | 51.65 ± 3.21 | 50.89 ± 3.34 | 2.75 | 0.104 |
| Bacteria PLFAs (nmol g−1) | 39.25 ± 2.71 | 40.16 ± 2.63 | 38.87 ± 2.85 | 2.66 | 0.110 | |
| Fungal PLFAs (nmol g−1) | 3.62 ± 0.25 | 4.05 ± 0.32 | 3.75 ± 0.33 | 4.23 | 0.041 |
| Plantation | Species | Component | Allometric Equation | Correlation Coefficient (R) | F-Value |
|---|---|---|---|---|---|
| PEU | Eucalyptus | Stem | W = 0.03040 × (DBH2H)0.9320 | 0.968 | 709.10 ** |
| Bark | W = 0.00503 × (DBH2H)0.8887 | 0.961 | 577.04 ** | ||
| Branches | W = 0.00196 × (DBH2H)0.9836 | 0.826 | 102.78 ** | ||
| Leaves | W = 0.000223 × (DBH2H)1.1627 | 0.767 | 68.73 ** | ||
| Roots | W = 0.00445 × (DBH2H)1.0069 | 0.951 | 452.86 ** | ||
| PCH | C. hystrix | Stem | W = 0.04350 × (DBH2H)0.8886 | 0.996 | 1767.57 ** |
| Bark | W = 0.01005 × (DBH2H)0.8102 | 0.986 | 526.73 ** | ||
| Branches | W = 0.02211 × (DBH2H)0.8333 | 0.958 | 168.53 ** | ||
| Leaves | W = 0.14336 × (DBH2H)0.4479 | 0.894 | 59.97 ** | ||
| Roots | W = 0.03544 × (DBH2H)0.7768 | 0.977 | 316.78 ** | ||
| MEC | Eucalyptus | Stem | W = 0.02529 × (DBH2H)0.9576 | 0.960 | 624.13 ** |
| Bark | W = 0.00950 × (DBH2H)0.8079 | 0.953 | 522.27 ** | ||
| Branches | W = 0.00350 × (DBH2H)0.9167 | 0.871 | 166.05 ** | ||
| Leaves | W = 0.00292 × (DBH2H)0.8382 | 0.654 | 39.59 ** | ||
| Roots | W = 0.00877 × (DBH2H)0.9219 | 0.947 | 456.27 ** | ||
| C. hystrix | Stem | W = 0.05568 × (DBH2H)0.8618 | 0.979 | 233.62 ** | |
| Bark | W = 0.00606 × (DBH2H)0.8607 | 0.978 | 217.75 ** | ||
| Branches | W = 0.12859 × (DBH2H)0.6434 | 0.869 | 30.89 ** | ||
| Leaves | W = 0.01528 × (DBH2H)0.8817 | 0.873 | 25.63 ** | ||
| Roots | W = 0.05587 × (DBH2H)0.7277 | 0.946 | 84.99 ** |
| Factor | Eucalyptus | C. hystrix | ||
|---|---|---|---|---|
| PEU | MEC | PCH | MEC | |
| Tree height (m) | 19.52 ± 0.15 | 20.99 ± 0.20 ** | 9.93 ± 0.13 | 11.69 ± 0.23 ** |
| DBH (cm) | 13.62 ± 0.16 | 14.58 ± 0.17 ** | 7.89 ± 0.15 | 9.81 ± 0.23 ** |
| Volume (m3 tree−1) | 0.142 ± 0.004 | 0.170 ± 0.005 ** | 0.031 ± 0.002 | 0.050 ± 0.003 ** |
| Biomass (kg tree−1) | 104.62 ± 2.65 | 125.75 ± 3.46 ** | 31.75 ± 1.33 | 59.07 ± 2.80 ** |
| Component | Eucalyptus | C. hystrix | ||
|---|---|---|---|---|
| PEU | MEC | PCH | MEC | |
| Stem | 47.06 ± 2.58 | 51.12 ± 1.18 ns | 50.92 ± 0.74 | 50.84 ± 1.22 ns |
| Bark | 38.39 ± 2.34 | 44.07 ± 1.90 ns | 48.56 ± 0.61 | 47.54 ± 0.70 ns |
| Branch | 44.15 ± 1.62 | 48.46 ± 1.34 ns | 51.41 ± 0.29 | 50.49 ± 1.76 ns |
| Leaf | 48.53 ± 2.26 | 51.36 ± 2.53 ns | 49.96 ± 0.42 | 46.03 ± 2.48 ns |
| Root | 44.12 ± 1.43 | 48.11 ± 1.11 ns | 48.11 ± 0.76 | 49.35 ± 2.20 ns |
| Layer | Component | PEU | PCH | MEC | F(2,12) | p |
|---|---|---|---|---|---|---|
| Understory | Aboveground | 42.29 ± 0.30 b | 46.54 ± 0.30 a | 46.83 ± 0.37 a | 61.29 | 0.000 |
| Belowground | 37.62 ± 0.45 b | 44.15 ± 0.91 a | 43.04 ± 0.39 a | 30.85 | 0.000 | |
| Litter | 37.63 ± 1.93 b | 40.29 ± 2.32 b | 47.16 ± 1.07 a | 7.08 | 0.009 | |
| Soil | 0–20 cm | 2.00 ± 0.09 c | 3.40 ± 0.15 a | 2.70 ± 0.17 b | 25.95 | 0.000 |
| 20–40 cm | 0.75 ± 0.09 b | 1.89 ± 0.14 a | 1.58 ± 0.11 a | 26.95 | 0.000 | |
| 40–60 cm | 0.66 ± 0.06 b | 1.21 ± 0.20 a | 1.39 ± 0.17 a | 6.02 | 0.015 | |
| 60–80 cm | 0.30 ± 0.08 b | 0.90 ± 0.12 a | 1.17 ± 0.16 a | 13.33 | 0.001 | |
| 80–100 cm | 0.47 ± 0.06 b | 1.19 ± 0.35 a | 1.10 ± 0.10 a | 3.42 | 0.067 | |
| Mean | 0.83 ± 0.01 b | 1.72 ± 0.20 a | 1.59 ± 0.13 a | 31.64 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhou, X.; Wen, Y.; Sun, D. Non-Additive Effects of Mixing Eucalyptus and Castanopsis hystrix Trees on Carbon Stocks under an Eco-Silviculture Regime in Southern China. Forests 2022, 13, 733. https://doi.org/10.3390/f13050733
Wang L, Zhou X, Wen Y, Sun D. Non-Additive Effects of Mixing Eucalyptus and Castanopsis hystrix Trees on Carbon Stocks under an Eco-Silviculture Regime in Southern China. Forests. 2022; 13(5):733. https://doi.org/10.3390/f13050733
Chicago/Turabian StyleWang, Lei, Xiaoguo Zhou, Yuanguang Wen, and Dongjing Sun. 2022. "Non-Additive Effects of Mixing Eucalyptus and Castanopsis hystrix Trees on Carbon Stocks under an Eco-Silviculture Regime in Southern China" Forests 13, no. 5: 733. https://doi.org/10.3390/f13050733