
Effects of Controlled Mycorrhization and Deficit Irrigation in the Nursery on Post-Transplant Growth and Physiology of Acer campestre L. and Tilia cordata Mill.

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
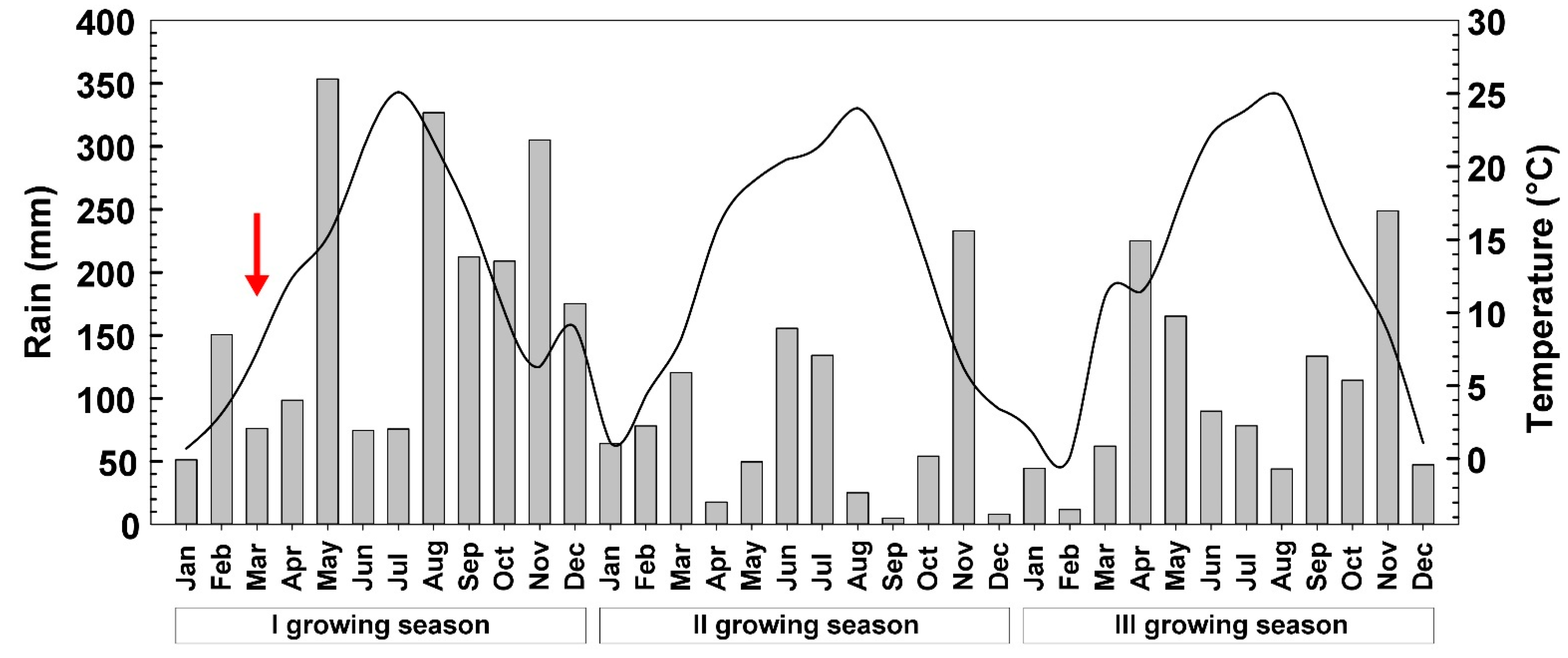
2.1. Plant Material and Experimental Site
2.2. Measurements
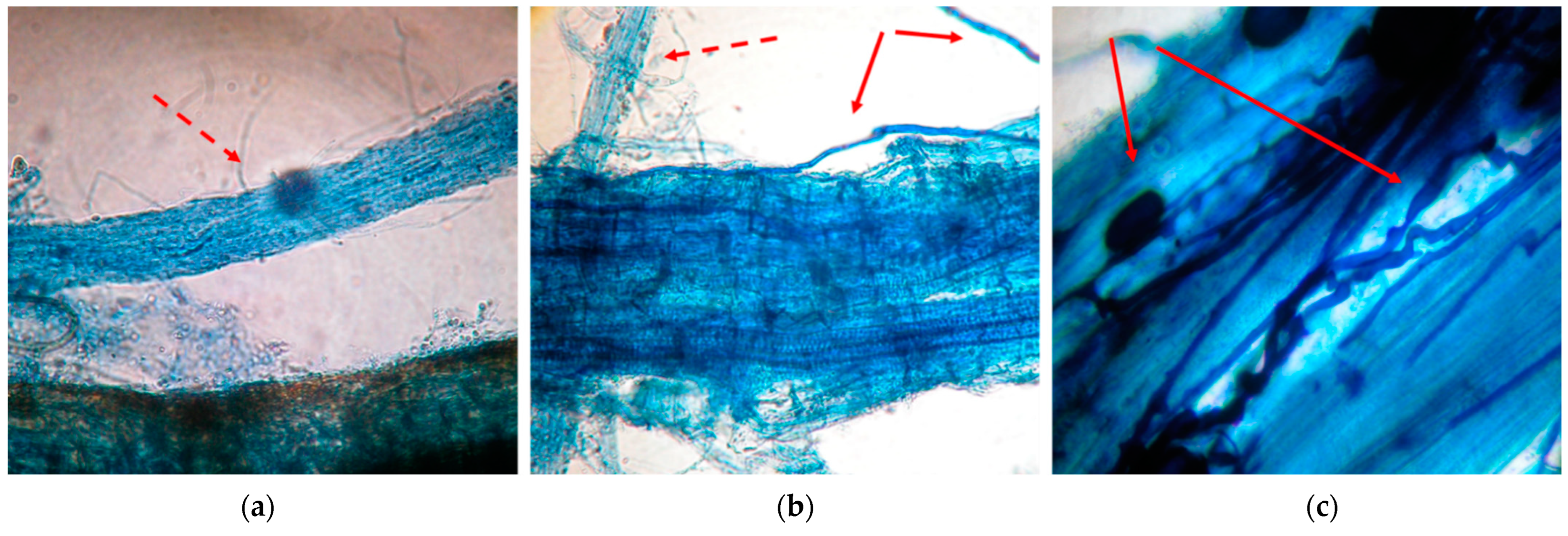
2.2.1. Root Colonization
2.2.2. Plant Mortality and Growth
2.2.3. Leaf Gas Exchange and Chlorophyll Fluorescence
2.2.4. Water Relations and Soluble Carbohydrates
2.3. Statistical Analysis
3. Results
3.1. Root Colonization
3.2. Plant Mortality and Growth
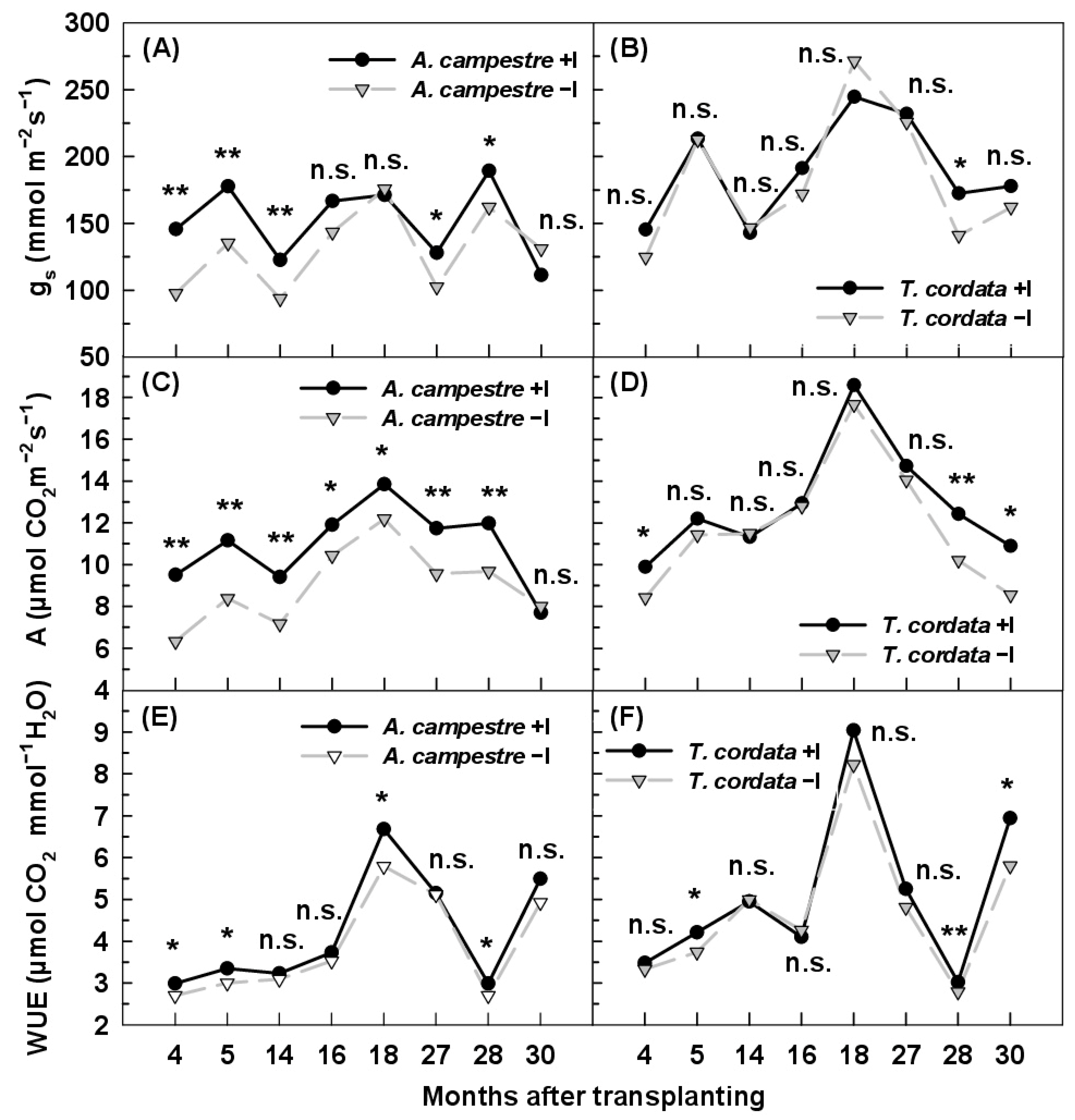
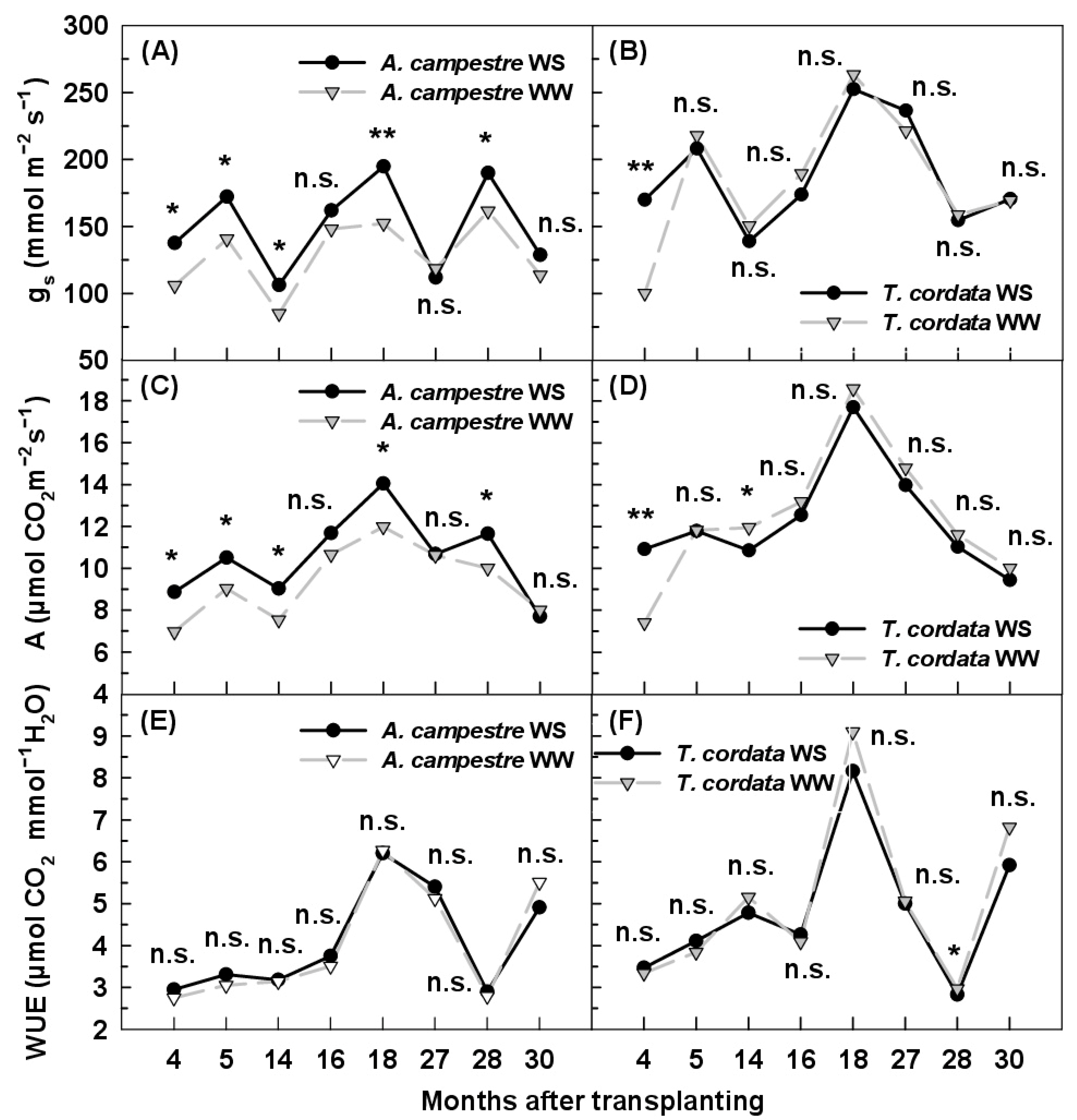
3.3. Leaf Gas Exchange
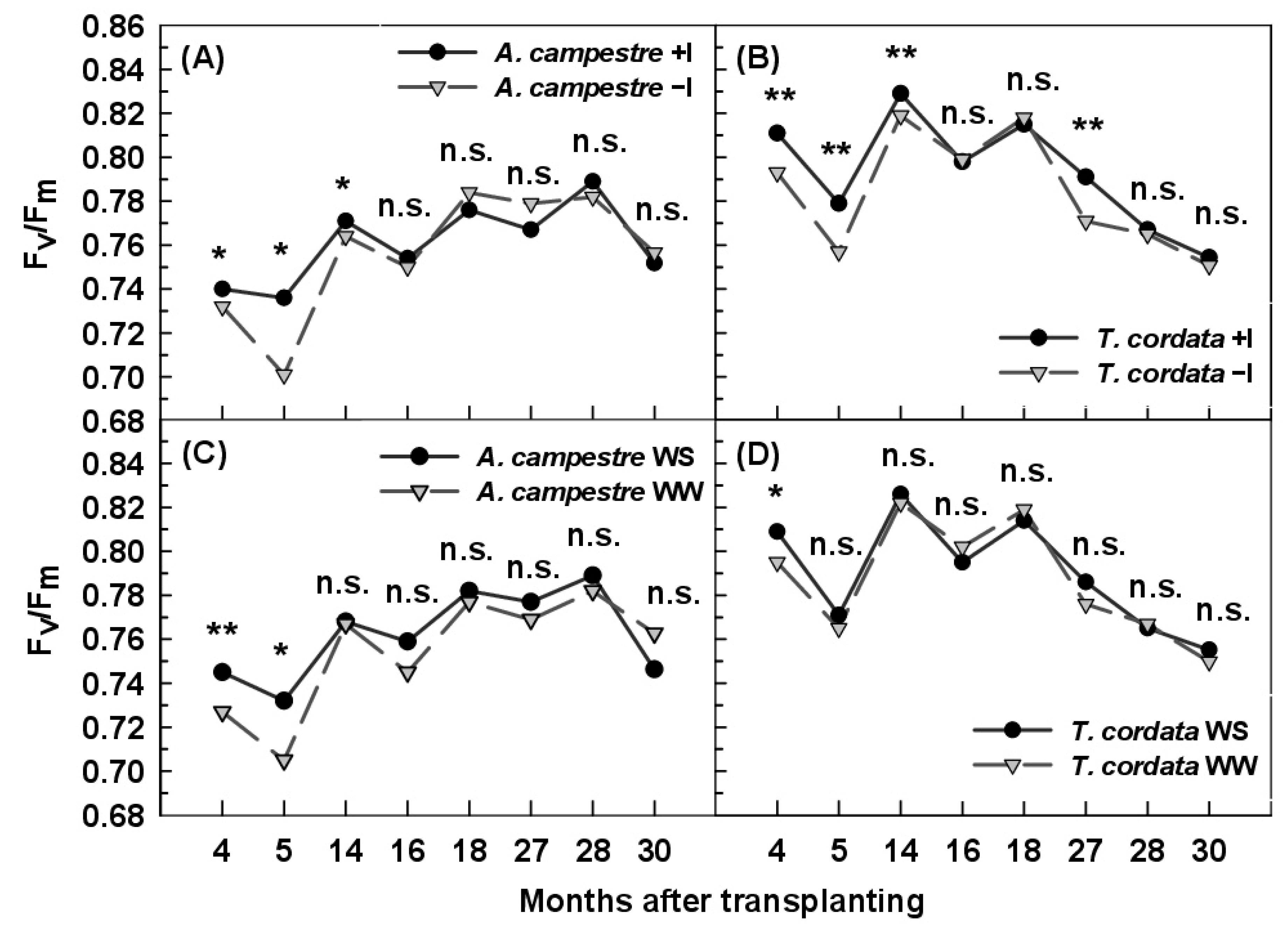
3.4. Chlorophyll Fluorescence
3.5. Water Relations
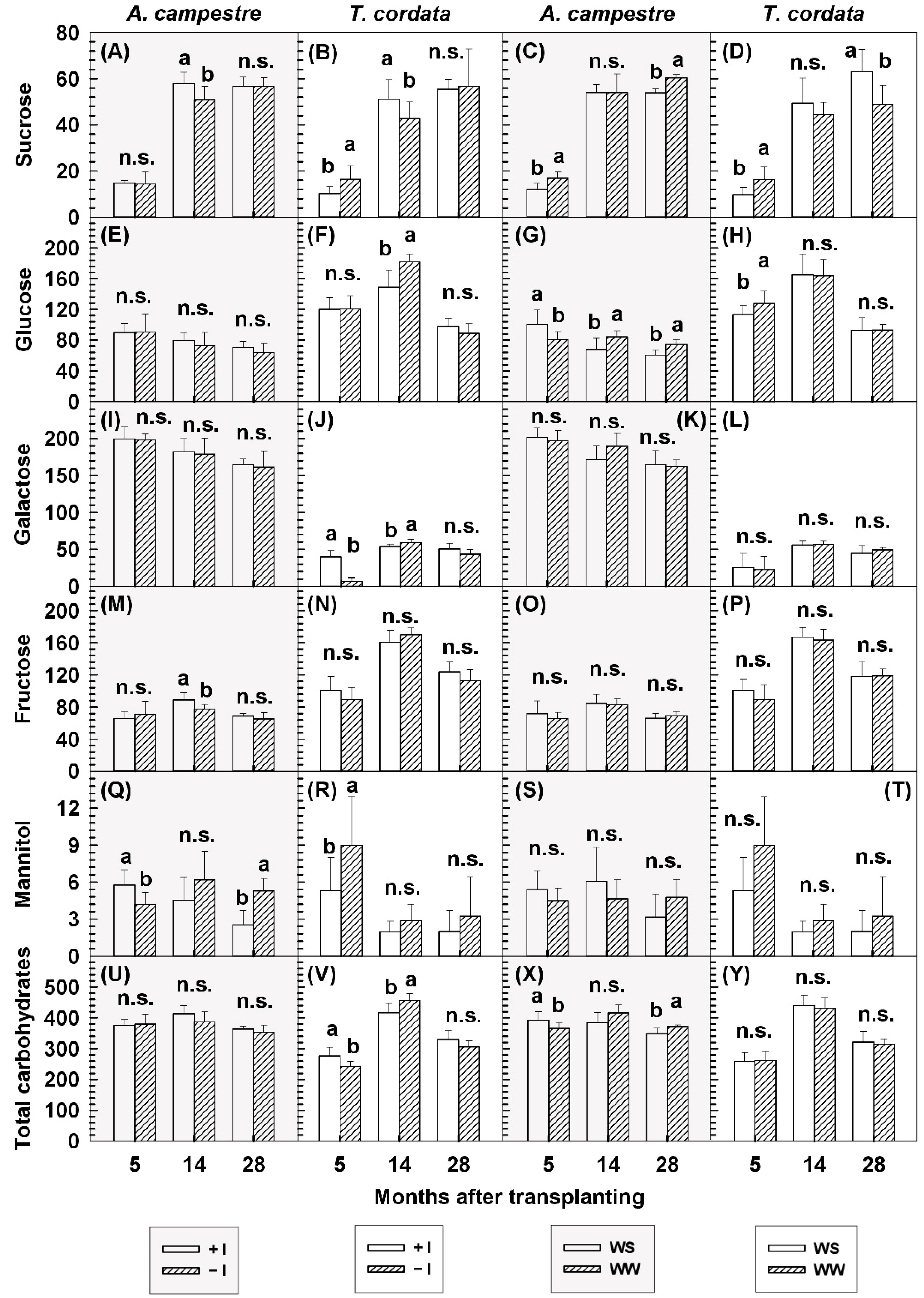
3.6. Soluble Carbohydrates
4. Discussion
4.1. Effects of Mycorrhizal Inoculation in the Nursery on Post-Transplant Growth and Physiology
4.2. Effects of Deficit Irrigation in the Nursery on Post-Transplant Growth and Physiology
4.3. Effects of Deficit Irrigation on Post-Transplant Root Mycorrhization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


References
- Szmigiera, M. Degree of Urbanization (Percentage of Urban Population in Total Population) by Continent in 2021. Available online: https://www.statista.com/statistics/270860/urbanization-by-continent/#statisticContainer (accessed on 22 June 2021).
- Ferrini, F.; Fini, A.; Mori, J.; Gori, A. Role of Vegetation as a Mitigating Factor in the Urban Context. Sustainability 2020, 12, 4247. [Google Scholar] [CrossRef]
- Vigevani, I.; Corsini, D.; Mori, J.; Pasquinelli, A.; Gibin, M.; Comin, S.; Szwałko, P.; Cagnolati, E.; Ferrini, F.; Fini, A. Particulate Pollution Capture by Seventeen Woody Species Growing in Parks or along Roads in Two European Cities. Sustainability 2022, 14, 1113. [Google Scholar] [CrossRef]
- Sicard, P.; Agathokleous, E.; Araminiene, V.; Carrari, E.; Hoshika, Y.; De Marco, A.; Paoletti, E. Should We See Urban Trees as Effective Solutions to Reduce Increasing Ozone Levels in Cities? Environ. Pollut. 2018, 243, 163–176. [Google Scholar] [PubMed]
- Weissert, L.F.; Salmond, J.A.; Schwendenmann, L. Photosynthetic CO2 uptake and carbon sequestration potential of deciduous and evergreen tree species in an urban environment. Urban. Ecosyst. 2016, 20, 663–674. [Google Scholar] [CrossRef]
- Sanusi, R.; Johnstone, D.; May, P.; Livesley, S. Microclimate benefits that different street tree species provide to sidewalk pedestrians relate to differences in Plant Area Index. Landsc. Urban Plan. 2017, 157, 502–511. [Google Scholar] [CrossRef]
- Speak, A.; Montagnani, L.; Wellstein, C.; Zerbe, S. The influence of tree traits on urban ground surface shade cooling. Landsc. Urban Plan. 2020, 197, 103748. [Google Scholar] [CrossRef]
- Tan, P.Y.; Wong, N.H.; Tan, C.L.; Jusuf, S.K.; Schmiele, K.; Chiam, Z.Q. Transpiration and cooling potential of tropical urban trees from different native habitats. Sci. Total Environ. 2020, 705, 135764. [Google Scholar] [CrossRef]
- Fini, A.; Frangi, P.; Mori, J.; Donzelli, D.; Ferrini, F. Nature based solutions to mitigate soil sealing in urban areas: Results from a 4-year study comparing permeable, porous, and impermeable pavements. Environ. Res. 2017, 156, 443–454. [Google Scholar] [CrossRef]
- Koeser, A.K.; Gilman, E.F.; Paz, M.; Harchick, C. Factors influencing urban tree planting program growth and survival in Florida, United States. Urban For. Urban Green. 2014, 13, 655–661. [Google Scholar] [CrossRef]
- Gilman, E.; Masters, F. Effect of Tree Size, Root Pruning, and Production Method on Root Growth and Lateral Stability of Quercus virginiana. Arboric. Urban For. 2010, 36, 281–291. [Google Scholar] [CrossRef]
- Khan, R.; Ma, X.; Shah, S.; Wu, X.; Shaheen, A.; Xiao, L.; Wu, Y.; Wang, S. Drought-hardening improves drought tolerance in Nicotiana tabacum at physiological, biochemical, and molecular levels. BMC Plant Biol. 2020, 20, 486. [Google Scholar] [CrossRef] [PubMed]
- Roloff, A.; Korn, S.; Gillner, S. The Climate-Species-Matrix to select tree species for urban habitats considering climate change. Urban For. Urban Green. 2009, 8, 295–308. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Bussotti, F.; Ferrini, F.; Pollastrini, M.; Fini, A. The challenge of Mediterranean sclerophyllous vegetation under climate change: From acclimation to adaptation. Environ. Exp. Bot. 2014, 103, 80–98. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef]
- Fini, A.; Ferrini, F.; Di Ferdinando, M.; Brunetti, C.; Giordano, C.; Gerini, F.; Tattini, M. Acclimation to partial shading or full sunlight determines the performance of container-grown Fraxinus ornus to subsequent drought stress. Urban For. Urban Green. 2014, 13, 63–70. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Franco, J.A.; Martínez-Sánchez, J.J.; Fernández, J.A.; Bañón, S. Selection and nursery production of ornamental plants for landscaping and xerogardening in semi-arid environments. J. Hortic. Sci. Biotechnol. 2006, 81, 3–17. [Google Scholar] [CrossRef]
- Vilagrosa, A.; Cortina, J.; Gil-Pelegrin, E.; Bellot, J. Suitability of Drought-Preconditioning Techniques in Mediterranean Climate. Restor. Ecol. 2003, 11, 208–216. [Google Scholar] [CrossRef]
- Rincón, A.; Parladé, J.; Pera, J. Influence of the fertilisation method in controlled ectomycorrhizal inoculation of two Mediterranean pines. Ann. For. Sci. 2007, 64, 577–583. [Google Scholar] [CrossRef] [Green Version]
- Augé, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Itoo, Z.A.; Reshi, Z.A. The Multifunctional Role of Ectomycorrhizal Associations in Forest Ecosystem Processes. Bot. Rev. 2013, 79, 371–400. [Google Scholar] [CrossRef]
- Giovannini, L.; Palla, M.; Agnolucci, M.; Avio, L.; Sbrana, C.; Turrini, A.; Giovannetti, M. Arbuscular Mycorrhizal Fungi and Associated Microbiota as Plant Biostimulants: Research Strategies for the Selection of the Best Performing Inocula. Agronomy 2020, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Augé, R.M.; Toler, H.D.; Sams, C.E.; Nasim, G. Hydraulic conductance and water potential gradients in squash leaves showing mycorrhiza-induced increases in stomatal conductance. Mycorrhiza 2008, 18, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Lehto, T.; Zwiazek, J.J. Ectomycorrhizas and water relations of trees: A review. Mycorrhiza 2010, 21, 71–90. [Google Scholar] [CrossRef]
- Augé, R.M.; Toler, H.D.; Saxton, A. Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: A meta-analysis. Mycorrhiza 2015, 25, 13–24. [Google Scholar] [CrossRef]
- Porcel, R.; Ruiz-Lozano, J.M. Arbuscular Mycorrhizal Influence on Leaf Water Potential, Solute Accumulation, and Oxidative Stress in Soybean Plants Subjected to Drought Stress. J. Exp. Bot. 2004, 55, 1743–1750. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, N.; Armada, E.; Duque, E.; Roldan, A.; Azcón, R. Contribution of arbuscular mycorrhizal fungi and/or bacteria to enhancing plant drought tolerance under natural soil conditions: Effectiveness of autochthonous or allochthonous strains. J. Plant Physiol. 2015, 174, 87–96. [Google Scholar] [CrossRef]
- Schellenbaum, L.; Sprenger, N.; Schüepp, H.; Wiemken, A.; Boller, T. Effects of drought, transgenic expression of a fructan synthesizing enzyme and of mycorrhizal symbiosis on growth and soluble carbohydrate pools in tobacco plants. New Phytol. 1999, 142, 67–77. [Google Scholar] [CrossRef]
- Ferrini, F.; Fini, A.; Marasco, P.L.; Pennati, L.; Sani, L. How to Select Trees That Will Thrive in the Urban Environments, given Differences in Urban Sites, Species Attribute, Management Requirements and Global Change. Proc. ISAAC 2008, May, 9–14. [Google Scholar]
- Szabo, K.; Lovas, V.; Eros-Honti, Z. The First Extensive Case-Study on the Effects of Artificial Mycorrhization of Acer Buergerianum Miq. in Hungary. Univers. J. Plant Sci. 2015, 3, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Le Tacon, F.; Alvarez, I.F.; Bouchard, D.; Henrion, B.; Jackson, R.M.; Luff, S.; Parlade, J.I.; Pera, J.; Stenström, E.; Villeneuve, N. Variations in Field Response of Forest Trees to Nursery Ectomycorrhizal Inoculation in Europe. In 3. European Symposium on Mycorrhizas; CAB International: Wallingford, UK, 1991. [Google Scholar]
- Garbaye, J.; Churin, J. Effect of Ectomycorrhizal Inoculation at Planting on Growth and Foliage Quality of Tilia Tomentosa. Arboric. Urban For. 1996, 22, 29–34. [Google Scholar] [CrossRef]
- Fini, A.; Frangi, P.; Amoroso, G.; Piatti, R.; Faoro, M.; Bellasio, C.; Ferrini, F. Effect of controlled inoculation with specific mycorrhizal fungi from the urban environment on growth and physiology of containerized shade tree species growing under different water regimes. Mycorrhiza 2011, 21, 703–719. [Google Scholar] [CrossRef]
- Johnson, N.C.; Pfleger, F. Vesicular-Arbuscular Mycorrhizae and Cultural Stresses. Anim. Manure 2015, 54, 71–99. [Google Scholar] [CrossRef]
- Fini, A.; Ferrini, F.; Seri, M.; Amoroso, G.; Piatti, R.; Robbiani, E.; Frangi, P. Effect of fertilization and mycorrhizal inoculation in the nursery on post-transplant growth and physiology in three ornamental woody species. Acta Hortic. 2016, 1108, 47–54. [Google Scholar] [CrossRef]
- Ruiz-Sánchez, M.; Domingo, R.; Torrecillas, A.; Pérez-Pastor, A. Water stress preconditioning to improve drought resistance in young apricot plants. Plant Sci. 2000, 156, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Blanco, M.; Ortuño, M.; Bañon, S.; Álvarez, S. Deficit irrigation as a strategy to control growth in ornamental plants and enhance their ability to adapt to drought conditions. J. Hortic. Sci. Biotechnol. 2019, 94, 137–150. [Google Scholar] [CrossRef]
- Bañon, S.; Fernandez, J.; Franco, J.; Torrecillas, A.; Alarcón, J.; Sánchez-Blanco, M. Effects of water stress and night temperature preconditioning on water relations and morphological and anatomical changes of Lotus creticus plants. Sci. Hortic. 2004, 101, 333–342. [Google Scholar] [CrossRef]
- Franco, J.A.; Cros, V.; Bañón, S.; González, A.; Abrisqueta, J.M. Effects of Nursery Irrigation on Postplanting Root Dynamics of Lotus creticus in Semiarid Field Conditions. HortScience 2002, 37, 525–528. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Álvarez, S.; Ortuño, M.F.; Ruiz-Sánchez, M.C. Root System Response to Drought and Salinity: Root Distribution and Water Transport. Soil Enzymol. 2014, 40, 325–352. [Google Scholar] [CrossRef]
- Navarro, A.; Álvarez, S.; Castillo, M.; Bañón, S.; Sánchez-Blanco, M.J. Changes in tissue-water relations, photosynthetic activity, and growth of Myrtus communis plants in response to different conditions of water availability. J. Hortic. Sci. Biotechnol. 2009, 84, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding Plant Responses to Drought—From Genes to the Whole Plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Xia, R.-X. Arbuscular mycorrhizal fungi influence growth, osmotic adjustment and photosynthesis of citrus under well-watered and water stress conditions. J. Plant Physiol. 2006, 163, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Dichio, B.; Xiloyannis, C.; Sofo, A.; Montanaro, G. Osmotic regulation in leaves and roots of olive trees during a water deficit and rewatering. Tree Physiol. 2006, 26, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; Volume 29, pp. 153–188. [Google Scholar] [CrossRef] [Green Version]
- García, A.N.; Árias, S.D.P.B.; Morte, A.; Sánchez-Blanco, M.J. Effects of nursery preconditioning through mycorrhizal inoculation and drought in Arbutus unedo L. plants. Mycorrhiza 2010, 21, 53–64. [Google Scholar] [CrossRef]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161, IN16–IN18. [Google Scholar] [CrossRef]
- McGONIGLE, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Jifon, J.; Syvertsen, J.P. Moderate shade can increase net gas exchange and reduce photoinhibition in citrus leaves. Tree Physiol. 2003, 23, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Fini, A.; Ferrini, F.; Frangi, P.; Amoroso, G.; Piatti, R. Withholding Irrigation During the Establishment Phase Affected Growth and Physiology of Norway Maple (Acer platanoides) and Linden (Tilia spp.). Arboric. Urban For. 2009, 35, 241–251. [Google Scholar] [CrossRef]
- Fini, A.; Frangi, P.; Faoro, M.; Piatti, R.; Amoroso, G.; Ferrini, F. Effects of different pruning methods on an urban tree species: A four-year-experiment scaling down from the whole tree to the chloroplasts. Urban For. Urban Green. 2015, 14, 664–674. [Google Scholar] [CrossRef]
- Lawlor, D.W. Limitation to Photosynthesis in Water-stressed Leaves: Stomata vs. Metabolism and the Role of ATP. Ann. Bot. 2002, 89, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Bernacchi, C.J. Gas Exchange Measurements, What Can They Tell Us about the Underlying Limitations to Photosynthesis? Procedures and Sources of Error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [Green Version]
- Grassi, G.; Magnani, F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Bernacchi, C.J.; Farquhar, G.D.; Singsaas, E.L. Fitting photosynthetic carbon dioxide response curves for C3 leaves. Plant Cell Environ. 2007, 30, 1035–1040. [Google Scholar] [CrossRef]
- Harley, P.C.; Loreto, F.; Di Marco, G.; Sharkey, T.D. Theoretical Considerations when Estimating the Mesophyll Conductance to CO2 Flux by Analysis of the Response of Photosynthesis to CO2. Plant Physiol. 1992, 98, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Pinior, A.; Grunewaldt-Stöcker, G.; Von Alten, H.; Strasser, R.J. Mycorrhizal impact on drought stress tolerance of rose plants probed by chlorophyll a fluorescence, proline content and visual scoring. Mycorrhiza 2005, 15, 596–605. [Google Scholar] [CrossRef]
- Scholander, P.F.; Hammel, H.T.; Hemmingsen, E.A.; Bradstreet, E.D. Hydrostatic pressure and osmotic potential in leaves of mangroves and some other plants. Proc. Natl. Acad. Sci. USA 1964, 52, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Gucci, R.; Xiloyannis, C.; Flore, J.A. Gas Exchange Parameters, Water Relations and Carbohydrate Partitioning in Leaves of Field-grown Prunus Domestica Following Fruit Removal. Physiol. Plant. 1991, 83, 497–505. [Google Scholar] [CrossRef]
- Fini, A.; Guidi, L.; Giordano, C.; Baratto, M.C.; Ferrini, F.; Brunetti, C.; Calamai, L.; Tattini, M. Salinity stress constrains photosynthesis in Fraxinus ornus more when growing in partial shading than in full sunlight: Consequences for the antioxidant defence system. Ann. Bot. 2014, 114, 525–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tattini, M.; Montagni, G.; Traversi, M.L. Gas exchange, water relations and osmotic adjustment in Phillyrea latifolia grown at various salinity concentrations. Tree Physiol. 2002, 22, 403–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tattini, M.; Gucci, R.; Romani, A.; Baldi, A.; Everard, J.D. Changes in Non-structural Carbohydrates in Olive (Olea Europaea) Leaves during Root Zone Salinity Stress. Physiol. Plant. 1996, 98, 117–124. [Google Scholar] [CrossRef]
- El Mrabet, S.; Ouahmane, L.; Mousadik, E.A.; Msanda, F.; Abbas, Y. The Effectiveness of Arbuscular Mycorrhizal Inoculation and Bio-Compost Addition for Enhancing Reforestation with Argania Spinosa in Morocco. Open J. For. 2014, 4, 14–23. [Google Scholar]
- Davidson, B.E.; Novak, S.J.; Serpe, M.D. Consequences of inoculation with native arbuscular mycorrhizal fungi for root colonization and survival of Artemisia tridentata ssp. wyomingensis seedlings after transplanting. Mycorrhiza 2016, 26, 595–608. [Google Scholar] [CrossRef]
- Prematuri, R.; Turjaman, M.; Tawaraya, K. Effect of Arbuscular Mycorrhiza Fungal Inoculation on Growth of Tropical Tree Species under Nursery and Post-Opencast Bauxite Mining Field in Bintan Island, Indonesia. Int. J. Plant Soil Sci. 2020, 1–13. [Google Scholar] [CrossRef]
- Sebastiana, M.; Pereira, V.T.; Alcântara, A.; Pais, M.S.; da Silva, A.B. Ectomycorrhizal inoculation with Pisolithus tinctorius increases the performance of Quercus suber L. (Cork Oak) nursery and field seedlings. New For. 2013, 44, 937–949. [Google Scholar] [CrossRef]
- Rennenberg, H.; Loreto, F.; Polle, A.; Brilli, F.; Fares, S.; Beniwal, R.S.; Gessler, A. Physiological Responses of Forest Trees to Heat and Drought. Plant Biol. 2006, 8, 556–571. [Google Scholar] [CrossRef]
- Fini, A.; Loreto, F.; Tattini, M.; Giordano, C.; Ferrini, F.; Brunetti, C.; Centritto, M. Mesophyll conductance plays a central role in leaf functioning of Oleaceae species exposed to contrasting sunlight irradiance. Physiol. Plant. 2016, 157, 54–68. [Google Scholar] [CrossRef]
- Birhane, E.; Kuyper, T.W.; Sterck, F.J.; Gebrehiwot, K.; Bongers, F. Arbuscular mycorrhiza and water and nutrient supply differently impact seedling performance of dry woodland species with different acquisition strategies. Plant Ecol. Divers. 2015, 8, 387–399. [Google Scholar] [CrossRef]
- Wiseman, P.E.; Wells, C.E. Arbuscular Mycorrhizal Inoculation Affects Root Development of Acer and Magnolia Species. J. Environ. Hortic. 2009, 27, 70–79. [Google Scholar] [CrossRef]
- Vemmos, S.N.; Petri, E.; Stournaras, V. Seasonal changes in photosynthetic activity and carbohydrate content in leaves and fruit of three fig cultivars (Ficus carica L.). Sci. Hortic. 2013, 160, 198–207. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Cui, L.; Mu, L.-Q. Ectomycorrhizal communities associated with Tilia amurensis trees in natural versus urban forests of Heilongjiang in northeast China. J. For. Res. 2016, 27, 401–406. [Google Scholar] [CrossRef]
- Rudawska, M.; Kujawska, M.; Leski, T.; Janowski, D.; Karliński, L.; Wilgan, R. Ectomycorrhizal community structure of the admixture tree species Betula pendula, Carpinus betulus, and Tilia cordata grown in bare-root forest nurseries. For. Ecol. Manag. 2019, 437, 113–125. [Google Scholar] [CrossRef]
- Sýkorová, Z.; Rydlová, J.; Slavíková, R.; Ness, T.; Kohout, P.; Püschel, D. Forest reclamation of fly ash deposit: A field study on appraisal of mycorrhizal inoculation. Restor. Ecol. 2016, 24, 184–193. [Google Scholar] [CrossRef]
- Chen, Y.L.; Liu, R.J.; Bi, Y.L.; Feng, G. Use of Mycorrhizal Fungi for Forest Plantations and Minesite Rehabilitation. In Soil Enzymology; Springer Science and Business Media LLC: Berlin, Germany, 2014; pp. 325–355. [Google Scholar]
- Schellenbaum, L.; Muller, J.; Boller, T.; Wiemken, A.; Schuepp, H. Effects of drought on non-mycorrhizal and mycorrhizal maize: Changes in the pools of non-structural carbohydrates, in the activities of invertase and trehalase, and in the pools of amino acids and imino acids. New Phytol. 1998, 138, 59–66. [Google Scholar] [CrossRef]
- Nowak, J. Effects of Arbuscular Mycorrhizal Fungi and Organic Fertilization on Growth, Flowering, Nutrient Uptake, Photosynthesis and Transpiration of Geranium (Pelargonium Hortorum LH Bailey’Tango Orange. Symbiosis 2004, 37, 259–266. [Google Scholar]
- Ortuño, M.F.; Lorente, B.; Hernández, J.A.; Sánchez-Blanco, M.J. Mycorrhizal inoculation on compost substrate affects nutritional balance, water uptake and photosynthetic efficiency in Cistus albidus plants submitted to water stress. Braz. J. Bot. 2018, 41, 299–310. [Google Scholar] [CrossRef]
- Bañon, S.; Ochoa, J.; Franco, J.A.; Sánchez-Blanco, M.J.; Alarcon, J.J. Influence of Water Deficit and Low Air Humidity in the Nursery on Survival of Rhamnus Alaternus Seedlings Following Planting. J. Hortic. Sci. Biotechnol. 2003, 78, 518–522. [Google Scholar] [CrossRef]
- Ranney, T.; Bassuk, N.L.; Whitlow, T.H. Osmotic Adjustment and Solute Constituents in Leaves and Roots of Water-stressed Cherry (Prunus) Trees. J. Am. Soc. Hortic. Sci. 1991, 116, 684–688. [Google Scholar] [CrossRef]
- Bañon, S.; Ochoa, J.; Franco, J.; Alarcón, J.; Sánchez-Blanco, M. Hardening of oleander seedlings by deficit irrigation and low air humidity. Environ. Exp. Bot. 2006, 56, 36–43. [Google Scholar] [CrossRef]
- Martinez, P.F.; Hernández, M.; Franco, J.A.; Banón, S. Adaptación a Condiciones de Sequıa y Endurecimiento Previo Al Trasplante de Plantas de Lotus Creticus Cytidoides En Vivero. IV. Jornadas Del Grupo de Ornamentales. Actas Hortic. 2000, 31, 77–84. [Google Scholar]
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and Adaptive Responses of Woody Plants to Environmental Stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Cameron, R.W.F.; Harrison-Murray, R.S.; Atkinson, C.J.; Judd, H.L. Regulated deficit irrigation—A means to control growth in woody ornamentals. J. Hortic. Sci. Biotechnol. 2006, 81, 435–443. [Google Scholar] [CrossRef]
- Selig, M.; Bohne, H. Drought stress reactions of different populations of Quercus robur L. and Tilia cordata Mill.1. J. Environ. Hortic. 2017, 35, 6–12. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Poyatos, R.; Aguadé, D.; Retana, J.; Mencuccini, M. A new look at water transport regulation in plants. New Phytol. 2014, 204, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Feifel, M.; Karimi, Z.; Schuldt, B.; Choat, B.; Jansen, S. Leaf gas exchange performance and the lethal water potential of five European species during drought. Tree Physiol. 2015, 36, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Leuschner, C.; Wedde, P.; Lübbe, T. The relation between pressure–volume curve traits and stomatal regulation of water potential in five temperate broadleaf tree species. Ann. For. Sci. 2019, 76, 60. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Inoculation (I) | Water Regime (W) | Significance | ||||
|---|---|---|---|---|---|---|---|
| +I (%) | −I (%) | WW (%) | WS (%) | I | W | I × W | |
| Acer campestre (AMF) | 70.6 a | 63.0 b | 66.0 a | 67.6 a | * | n.s. | n.s. |
| Tilia cordata (AMF) | 38.6 a | 33.5 a | 33.8 a | 38.3 a | n.s. | n.s. | n.s. |
| Tilia cordata (ECM) | 87.7 a | 91.7 a | 90.0 a | 89.3 a | n.s. | n.s. | n.s. |
| Species | Parameter | Months after Transplanting | Inoculation (I) | Water Regime (W) | Significance | ||||
|---|---|---|---|---|---|---|---|---|---|
| +I (%) | −I (%) | WW (%) | WS (%) | I | W | I × W | |||
| A. campestre | Plant DW (g) | 34 | 3519 a | 3360 a | 3168 b | 3711 a | n.s. | * | n.s. |
| Leaf DW (g) | 34 | 535.5 a | 574.1 a | 515.3 a | 594.2 a | n.s. | n.s. | n.s. | |
| Shoot DW (g) | 34 | 2115 a | 2017 a | 1897 b | 2235 a | n.s. | * | n.s. | |
| Root DW (g) | 34 | 867.9 a | 768.6 a | 881.5 a | 755.0 a | n.s. | n.s. | n.s. | |
| Root:shoot (%) | 34 | 0.41 a | 0.38 a | 0.46 a | 0.34 a | n.s. | n.s. | n.s. | |
| Shoot length (cm) | 11 | 16.2 a | 19.1 a | 14.4 b | 20.9 a | n.s. | ** | n.s. | |
| Shoot length (cm) | 23 | 64.8 a | 56.8 b | 52.4 b | 69.3 a | * | ** | * | |
| Shoot length (cm) | 34 | 54.4 a | 45.6 b | 43.0 b | 56.9 a | * | ** | n.s. | |
| Stem diameter (cm) | 11 | 2.27 a | 2.24 a | 2.30 a | 2.20 a | n.s. | n.s. | n.s. | |
| Stem diameter (cm) | 23 | 3.64 a | 3.70 a | 3.61 a | 3.73 a | n.s. | n.s. | n.s. | |
| Stem diameter (cm) | 34 | 5.75 a | 5.68 a | 5.43 a | 6.00 a | n.s. | n.s. | n.s. | |
| T. cordata | Plant DW (g) | 34 | 2684 a | 3077 a | 3394 a | 2367 b | n.s. | ** | n.s. |
| Leaf DW (g) | 34 | 385.4 a | 486.8 a | 488.8 a | 383.4 a | n.s. | n.s. | n.s. | |
| Shoot DW (g) | 34 | 1702 a | 1962 a | 2170 a | 1495 b | n.s. | * | n.s. | |
| Root DW (g) | 34 | 596.9 a | 627.7 a | 735.7 a | 488.8 b | n.s. | ** | n.s. | |
| Root:shoot (%) | 34 | 0.35 a | 0.32 a | 0.34 a | 0.33 a | n.s. | n.s. | n.s. | |
| Shoot length (cm) | 11 | 16.4 a | 15.8 a | 19.1 a | 13.1 b | n.s. | ** | n.s. | |
| Shoot length (cm) | 23 | 78.5 a | 69.4 b | 78.4 a | 69.5 b | * | * | n.s. | |
| Shoot length (cm) | 34 | 64.3 a | 65.5 a | 64.6 a | 63.2 a | n.s. | n.s. | n.s. | |
| Stem diameter (cm) | 11 | 2.15 b | 2.32 a | 2.35 a | 2.12 b | * | ** | n.s. | |
| Stem diameter (cm) | 23 | 4.01 a | 4.03 a | 4.27 a | 3.78 b | n.s. | ** | n.s. | |
| Stem diameter (cm) | 34 | 6.01 a | 6.04 a | 6.47 a | 5.58 b | n.s. | * | n.s. | |
| Species | Parameter | Months after Transplanting | −I WW | +I WW | −I WS | +I WS | Significance | ||
|---|---|---|---|---|---|---|---|---|---|
| I | W | I × W | |||||||
| A. campestre | Vcmax (μmol m−2 s−1) | 4 | 10.7 d | 27.0 c | 46.8 b | 72.7 a | ** | ** | n.s. |
| Vcmax (μmol m−2 s−1) | 27 | 70.3 b | 80.2 ab | 70.3 b | 87.6 a | * | n.s. | n.s. | |
| Jmax (μmol m−2 s−1) | 4 | 27.8 c | 47.6 b | 79.5 a | 84.9 a | * | ** | * | |
| Jmax (μmol m−2 s−1) | 27 | 114.2 b | 133.9 a | 120.9 b | 140.1 a | ** | n.s. | n.s. | |
| Ls (%) | 4 | 0.06 c | 0.12 b | 0.12 b | 0.22 a | * | * | n.s. | |
| Ls (%) | 27 | 0.25 a | 0.26 a | 0.19 a | 0.25 a | n.s. | n.s. | n.s. | |
| Lns (%) | 4 | 0.67 a | 0.53 b | 0.01 c | 0.00 c | * | * | * | |
| Lns (%) | 27 | 0.19 a | 0.01 b | 0.07 ab | 0.00 b | ** | n.s. | n.s. | |
| gm (mmol m−2 s−1) | 27 | 62.0 b | 78.0 ab | 65.0 b | 92.0 a | * | n.s. | n.s. | |
| gs/gm | 27 | 2.29 a | 1.81 b | 2.23 a | 1.69 b | ** | n.s. | n.s. | |
| T. cordata | Vcmax (μmol m−2 s−1) | 4 | 39.4 b | 40.9 b | 45.7 b | 81.1 a | ** | ** | ** |
| Vcmax (μmol m−2 s−1) | 27 | 77.2 b | 89.9 a | 97.9 a | 79.5 b | n.s. | n.s. | ** | |
| Jmax (μmol m−2 s−1) | 4 | 58.1 b | 57.9 b | 80.9 ab | 121.9 a | * | ** | n.s. | |
| Jmax (μmol m−2 s−1) | 27 | 109.9 c | 154.2 a | 131.4 b | 125.0 b | ** | n.s. | ** | |
| Ls (%) | 4 | 0.19 a | 0.17 a | 0.19 a | 0.07 b | * | * | * | |
| Ls (%) | 27 | 0.24 a | 0.22 a | 0.24 a | 0.24 a | n.s. | n.s. | n.s. | |
| Lns (%) | 4 | 0.59 a | 0.55 a | 0.43 a | 0.00 b | * | * | * | |
| Lns (%) | 27 | 0.30 a | 0.00 b | 0.32 a | 0.12 ab | * | * | n.s. | |
| gm (mmol m−2 s−1) | 27 | 75.0 c | 111.0 a | 98.0 b | 65.0 c | n.s. | * | ** | |
| gs/gm | 27 | 1.75 a | 1.20 a | 0.87 a | 2.16 a | n.s. | n.s. | n.s. | |
| Species | Parameter | Months after Transplanting | Inoculation (I) | Water Regime (W) | Significance | ||||
|---|---|---|---|---|---|---|---|---|---|
| +I (%) | −I (%) | WW (%) | WS (%) | I | W | I × W | |||
| A. campestre | ΨW | 5 | −0.30 a | −0.44 b | −0.42 b | −0.32 a | ** | ** | n.s. |
| ΨW | 14 | −0.35 a | −0.40 b | −0.41 b | −0.34 a | * | ** | n.s. | |
| ΨW | 28 | −0.30 a | −0.40 b | −0.38 b | −0.32 a | ** | ** | n.s. | |
| ΨΠ | 5 | −1.74 a | −1.71 a | −1.76 a | −1.69 a | n.s. | n.s. | n.s. | |
| ΨΠ | 14 | −1.77 b | −1.66 a | −1.71 a | −1.73 a | * | n.s. | n.s. | |
| ΨΠ | 28 | −1.91 a | −1.90 a | −1.94 b | −1.87 a | n.s. | * | n.s. | |
| ΨP | 5 | 1.44 a | 1.27 b | 1.34 a | 1.37 a | ** | n.s. | n.s. | |
| ΨP | 14 | 1.42 a | 1.26 b | 1.30 a | 1.39 a | * | n.s. | n.s. | |
| ΨP | 28 | 1.61 a | 1.50 b | 1.56 a | 1.55 a | ** | n.s. | n.s. | |
| T. cordata | ΨW | 5 | −0.25 a | −0.36 b | −0.33 b | −0.28 a | ** | * | n.s. |
| ΨW | 14 | −0.22 a | −0.28 b | −0.25 a | −0.25 a | ** | n.s. | n.s. | |
| ΨW | 28 | −0.25 a | −0.31 b | −0.27 a | −0.29 a | ** | n.s. | n.s. | |
| ΨΠ | 5 | −1.47 a | −1.46 a | −1.43 a | −1.50 a | n.s. | n.s. | * | |
| ΨΠ | 14 | −1.74 b | −1.59 a | −1.64 a | −1.69 a | * | n.s. | n.s. | |
| ΨΠ | 28 | −1.76 a | −1.79 a | −1.69 a | −1.86 b | n.s. | ** | n.s. | |
| ΨP | 5 | 1.22 a | 1.10 a | 1.10 a | 1.22 a | n.s. | n.s. | * | |
| ΨP | 14 | 1.52 a | 1.31 b | 1.39 a | 1.44 a | ** | n.s. | n.s. | |
| ΨP | 28 | 1.51 a | 1.48 a | 1.42 b | 1.57 a | n.s. | ** | n.s. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corsini, D.; Vigevani, I.; Oggioni, S.D.; Frangi, P.; Brunetti, C.; Mori, J.; Viti, C.; Ferrini, F.; Fini, A. Effects of Controlled Mycorrhization and Deficit Irrigation in the Nursery on Post-Transplant Growth and Physiology of Acer campestre L. and Tilia cordata Mill. Forests 2022, 13, 658. https://doi.org/10.3390/f13050658
Corsini D, Vigevani I, Oggioni SD, Frangi P, Brunetti C, Mori J, Viti C, Ferrini F, Fini A. Effects of Controlled Mycorrhization and Deficit Irrigation in the Nursery on Post-Transplant Growth and Physiology of Acer campestre L. and Tilia cordata Mill. Forests. 2022; 13(5):658. https://doi.org/10.3390/f13050658
Chicago/Turabian StyleCorsini, Denise, Irene Vigevani, Silvio Daniele Oggioni, Piero Frangi, Cecilia Brunetti, Jacopo Mori, Carlo Viti, Francesco Ferrini, and Alessio Fini. 2022. "Effects of Controlled Mycorrhization and Deficit Irrigation in the Nursery on Post-Transplant Growth and Physiology of Acer campestre L. and Tilia cordata Mill." Forests 13, no. 5: 658. https://doi.org/10.3390/f13050658