1. Introduction
The terrestrial forest ecosystem provides a vital link to control the water exchange between land surface and atmosphere as well as to sequester the carbon by photosynthesis. In particular, boreal forest biomes cover approximately 11% of the area on the Earth’s surface [
1], which plays an important role in maintaining the stability of the terrestrial forest ecosystem. Thus, the interest in the functions and role of the boreal forest ecosystem in water resources is increasing, especially for the transpiration across the globe [
2,
3,
4,
5].
Canopy transpiration is a crucial process of forest hydrological cycling, which couples the soil moisture and atmosphere interactions [
6,
7,
8,
9]. The results at a global scale showed that canopy transpiration accounts for about 39% of precipitation and more than 60% of evapotranspiration [
10,
11]. In past decades, due to the frangibility and sensitivity of the boreal forest ecosystem to climatic change [
2], canopy transpiration has been increasingly influenced by global climate warming and extreme weather events [
12]. With the significant progress of afforestation projects in China, such as the Natural Forest Protection Project, the boreal forest coverage in China has been greatly improved, which has greatly affected the hydrological cycle of the forest ecosystem [
5,
13]. Thus, determining the canopy transpiration dynamics of boreal forest in a changing environment is greatly important for better understanding the plant survival strategies and the impact of vegetation on related eco-hydrological processes.
Numerous studies analyze the response of canopy transpiration to environmental factors including air temperature, vapor pressure deficit, soil moisture, precipitation, etc. [
14,
15,
16,
17]. For example, Zhang et al. [
18] concluded that increasing vapor pressure deficit promotes canopy transpiration until a certain threshold, while Han et al. [
19] found the precipitation has a negative effect on canopy transpiration. However, most often, case studies have only paid attention to the single impact of meteorological factors on canopy transpiration [
17,
20,
21]. We know that environmental factors occur concurrently under natural conditions and that canopy transpiration is affected by the interaction effects of environmental factors. However, the response mechanism of canopy transpiration to the interaction of environmental factors in the boreal forest of China is not clear.
Canopy transpiration is affected by many environmental factors, which can be divided into three aspects including atmospheric evaporative demand, such as potential evapotranspiration [
9,
22], soil water supply (such as soil moisture) and vegetation phenophase, such as leaf expanding or defoliation [
9,
23]. In previous work, some studies reported the effects of potential evapotranspiration and soil moisture on transpiration [
9,
21,
22]. For instance, Wan et al. [
24] found that soil moisture limits the response of transpiration to potential evapotranspiration. However, numerous studies only consider the atmospheric evaporative demand and water supply and ignore the change in vegetation phenophase during the growing season. Vegetation phenology reflects the dynamic of vegetation growth via leaf area variations and consequently affects the canopy transpiration [
23]. Thus, analyzing how potential evapotranspiration mediates the influence of soil water supply and leaf area variation on canopy transpiration is of great significance in the boreal forest regions of China.
The native larch (
Larix gmelinii (Rupr.) Rupr.) forest represents the typical zonal vegetation and prominent community of boreal forest ecosystems in northeast China, which is the southern margin of the Siberian zone and one of the largest boreal forests underlying wide permafrost in the world [
25,
26]. In the present study, a consecutive measurement of sap flow from May to September in 2021 in the natural forest of
Larix gmelinii was performed: (1) to investigate the dynamics of canopy transpiration of
Larix gmelinii forest in the growing season at different timescales, (2) to clarify the relationships between canopy transpiration and environmental factors, (3) to identify the role of soil moisture in canopy transpiration of
Larix gmelinii forest, and (4) to reveal how potential evapotranspiration mediates the influence of leaf area variation on canopy transpiration of
Larix gmelinii forest. These can be exploited to improve forest-water management and ecologically sustainable development, as well as to predict the effect of climate change on vegetation growth in the future.
4. Discussion
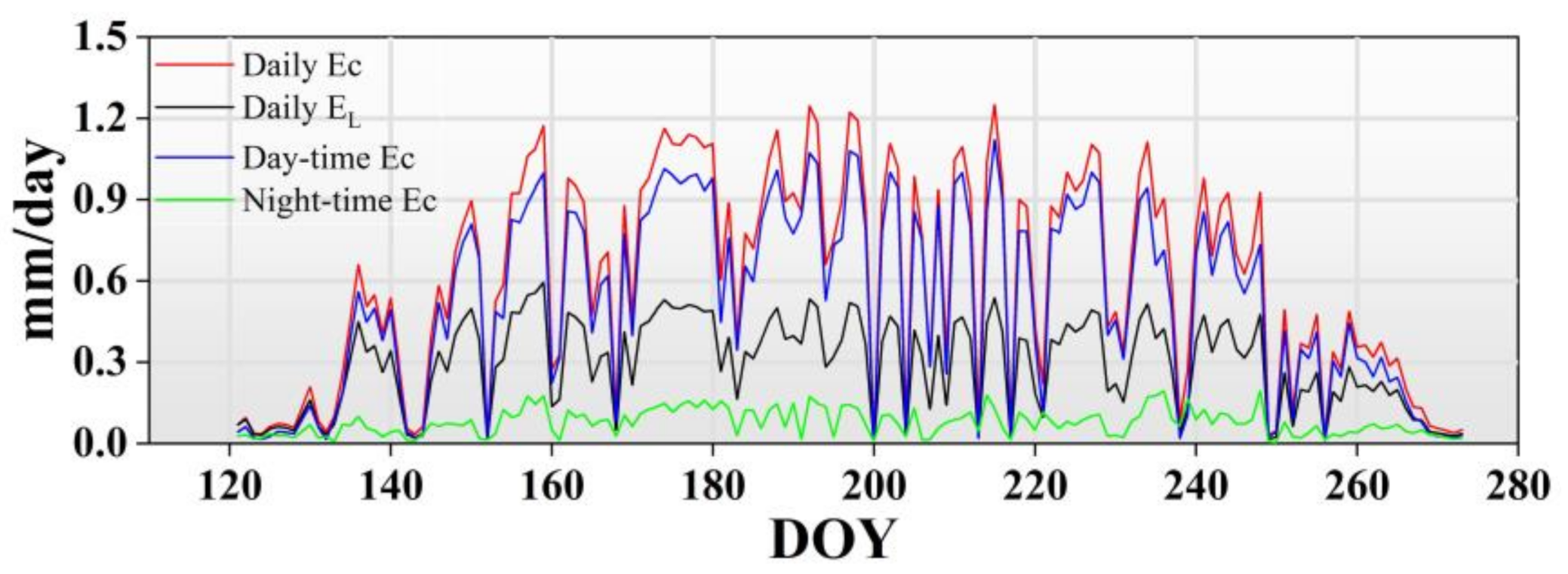
Canopy transpiration is one of the crucial methods of water consumption in the forest ecosystem, which affects the forest hydrological processes [
19,
38]. Knowledge of canopy transpiration is greatly important for the forest–water relationship under the background of carbon neutrality. In the present study, the daily mean Ec was 0.60 mm/day accounting for 16.50% of P during the measurement period, which was in a reasonable range compared with previous studies [
9,
17,
19,
21]. For instance, Wang et al. [
9] reported that the mean daily canopy transpiration of Larch was 0.70 mm/day, ranging from 0.02 to 1.55 mm in the growing season of 2018. However, we also found that the canopy transpiration of the present study was lower than the same tree species in the previous study [
26]. The difference in Ec may be caused by forest structure and climate condition of the measurement period. In the present study, the stand density was 1250 trees ha
−1, which was significantly lower than that for the 2350 trees ha
−1 of Liu et al. [
26]. Wang et al. [
4] demonstrated that the thinning significantly reduced daily transpiration at a stand scale. Moreover, the precipitation in the study period was greatly higher than the mean annual precipitation from 1957 to 2017. Wullschleger and Hanson [
39] indicated that excessive precipitation limited the level of transpiration. These reasons largely account for the lower daily canopy transpiration in the present study.
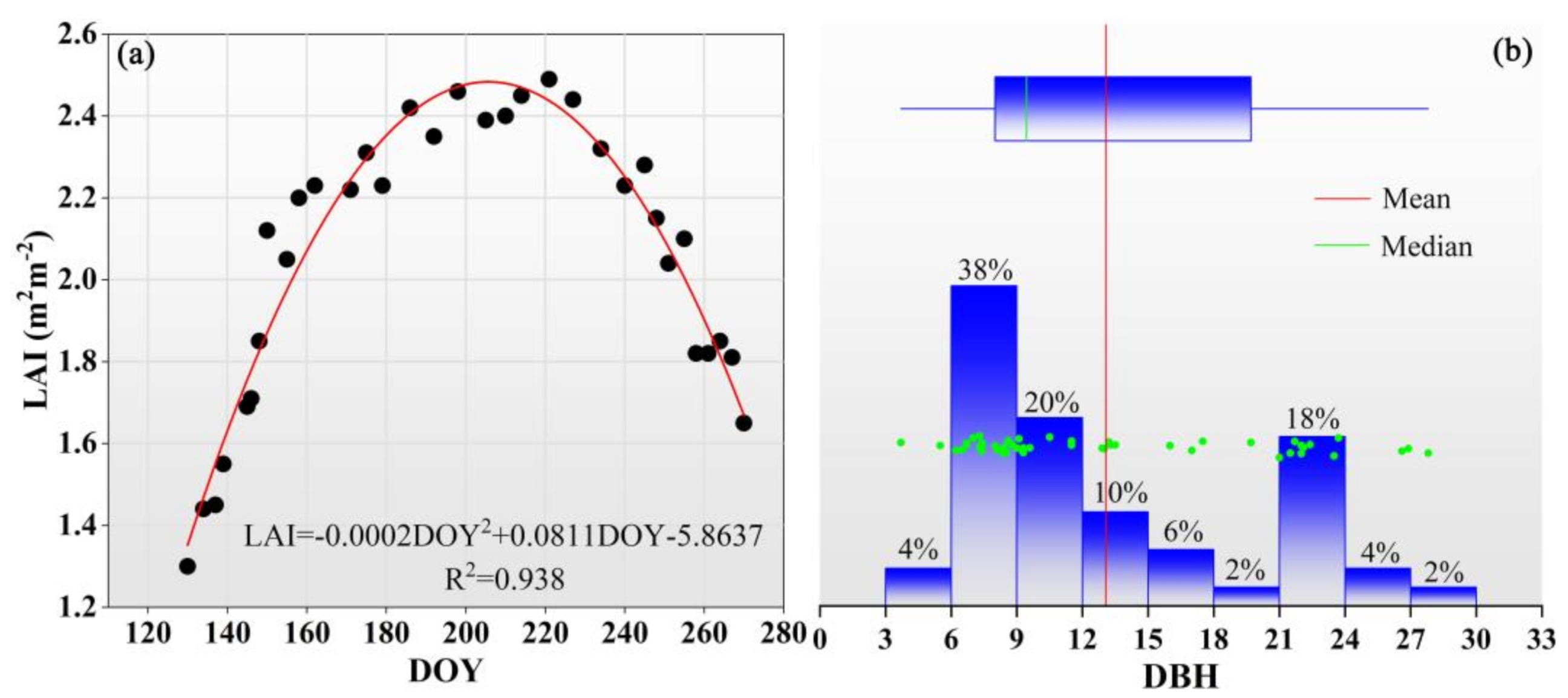
During the growing season, there were significant monthly dynamics in canopy transpiration. The higher canopy transpirations were found in June, July and August, which is consistent with other studies [
19,
38,
40]. However, previous studies also suggested that the canopy transpiration of Mongolian pine trees in May was higher than the other months attributed to the high-growth period [
41]. It is maybe closely related to the monthly dynamic of vegetation growth and climate conditions. It can be seen from
Figure 4 that the LAI and Ta in June, July and August were significantly higher than that of May and September in the present study, which leads to the higher canopy transpiration. Additionally, although the VPD and PAR in August were similar to May and September, the LAI and Ta were higher and subsequently promote canopy transpiration.
In addition, the results of correlation and regression analysis showed a significant relationship between Ec and environmental factors (
Table 3 and
Figure 7), which was consistent with the previous studies [
42,
43,
44,
45]. For instance, the relationship between Ec and PET has exhibited the exponential threshold function (
Figure 7) and indicated that Ec increases rapidly with the increasing of PET and then tended to be saturation when PET continued to increase. Wieser et al. [
42] suggested that the canopy stomatal conductance decreased with the increase in VPD. As shown in
Figure 8, our study also demonstrated that the canopy conductance (G
L) significantly decreased with the increase in VPD. Another possible reason is that the stomata need to be closed through the plant self-protection mechanism to maintain canopy transpiration to avoid excessive water consumption at higher atmospheric evaporative demand [
42,
46,
47]. Furthermore, in the present study, PET as a compound index is the major driver of transpiration rather than a single environmental variable. Similarly, Song et al. [
21] reported that the effects of PET on transpiration in three forest management areas of NE China were stronger than those of the other environmental variables. It is revealed that the compound index characterizing the atmospheric evaporative demand can better explain the change in transpiration. However, we found that the effect of soil moisture on the Ec of the
Larix gmelinii forest was lowest compared with other environment variables. Similar results were observed in the semiarid region and boreal forest [
3,
21,
48,
49]. On the contrary, some studies revealed that canopy transpiration is significantly influenced by soil moisture [
50]. These different results may be closely attributed to the different tree species, root distribution and climate conditions. Chu et al. [
51] reported that the root of
Larix gmelinii forest was mainly distributed in topsoil [
52] combined with the excessive precipitation in the study period, which led to sufficient soil moisture in topsoil to maintain water consumption of canopy transpiration and consequently the soil moisture was not the main limiting environmental factor of canopy transpiration of
Larix gmelinii forest.
Generally, canopy transpiration is influenced by the interaction effects of multiple environmental variables under natural conditions [
53]. In the present study, when soil moisture was more than 0.2 cm
3 cm
−3 (accounting for 94.77% (145 days) of the whole study period (153 days)), we found that the effect of PET on canopy transpiration was not limited by soil moisture. It is indicated that the water supply can meet the water consumption of canopy transpiration of
Larix gmelinii in the current soil moisture conditions. In addition, when PET was higher than the threshold value (PET > 9 mm/day), the response of canopy transpiration to LAI will be limited. For example, Song et al. [
45] reported that the soil moisture limits the response of transpiration to atmospheric water demand in seasonal drought, while the effect was not significant in the well-water soil moisture conditions of subtropical coniferous. Some similar results were also observed in Clausnitzer et al. [
33] and Zha et al. [
54]. Numerous studies have indicated that the limitations of soil moisture on the response of canopy transpiration to atmospheric evaporative demand will occur in drought conditions [
45,
54,
55]. When the soil moisture supply is insufficient or in high atmospheric evaporation demand, the leaf stomata and leaf water content of plants was closed and declined to prevent excessive water consumption [
56,
57,
58], which is consistent with the results in
Figure 8. On the other hand, it is well known that the most important water source of the forest ecosystem is from precipitation input. However, for the above-mentioned result that total canopy transpiration only accounts for 16.50% of P during the measurement period, this indicates that the water input though excessive precipitation can effectively increase soil moisture to maintain canopy transpiration in the present study. Moreover, the age of
Larix gmelinii in the study sample plot ranged from 75 to 90 years, which means it is a relatively mature stand [
59]. Bretfeld et al. [
60] reported that transpiration is more likely to be limited by soil moisture in young forests than that of old forests due to plant root volume difference. Altogether,
Larix gmelinii forest adjust its water consumption through canopy transpiration by responding to changes in the external environment.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}