
Loss of Gramicidin Biosynthesis in Gram-Positive Biocontrol Bacterium Aneurinibacillus migulanus (Takagi et al., 1993) Shida et al. 1996 Emend Heyndrickx et al., 1997 Nagano Impairs Its Biological Control Ability of Phytophthora


 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions for Bacteria
2.2. Growth Conditions for Phytophthora Species
2.3. Screening for Biosurfactant Activity Using Surface Tension Method
2.4. Co-Cultivation of A. migulanus and Plant Pathogens
2.5. Effects of A. migulanus Treatment on Phytophthora Disease Severity on Apple Fruits
2.6. Preparation of Gramicidin S Standard Curve
2.7. Extraction of Gramicidin S from A. migulanus Cultures
2.8. LC-MS Analysis
2.9. Statistical Analysis
3. Results
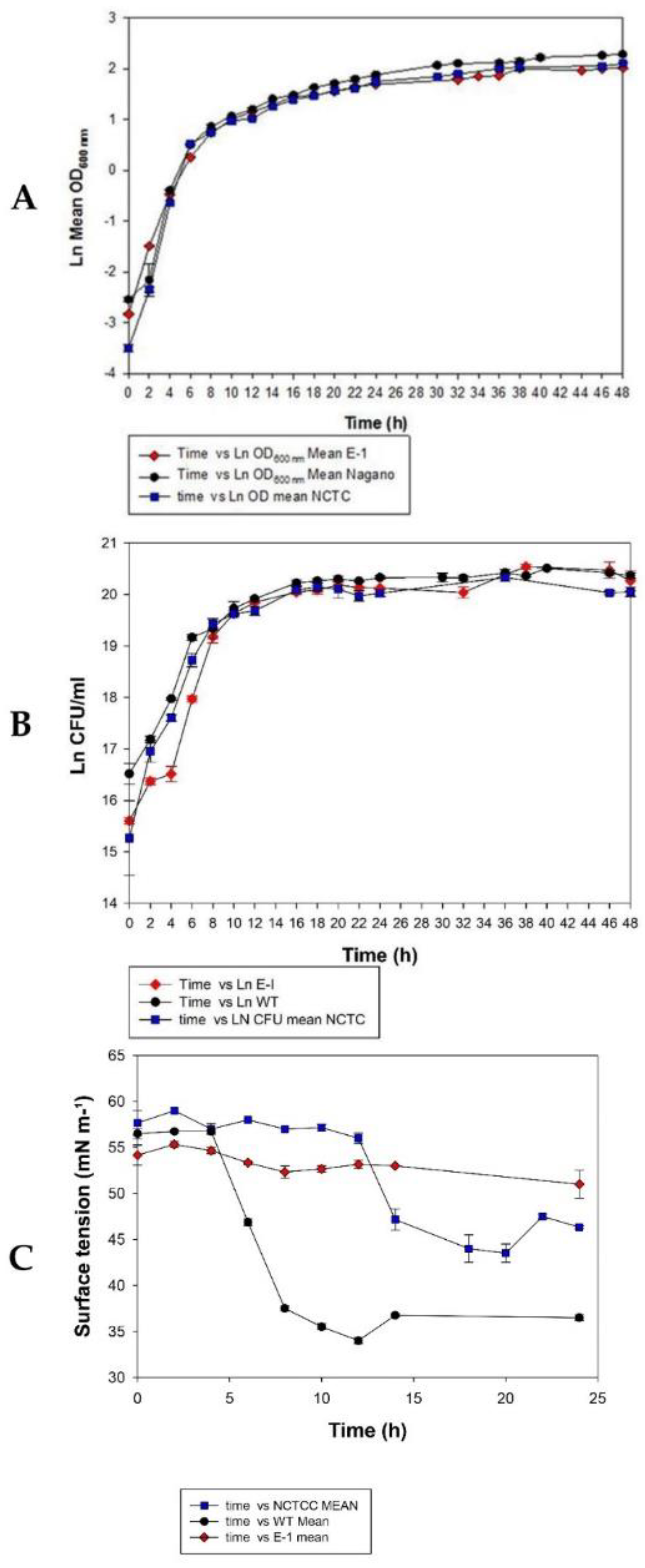
3.1. Growth Characteristics of A. migulanus Nagano, NCTC 7096, and E1 Mutant
3.2. Comparison of Biosurfactant Activity of A. migulanus Nagano, NCTC 7096, and E1 Mutant Strains Using Surface Tension Method
3.3. Gramicidin Production by A. migulanus Strains
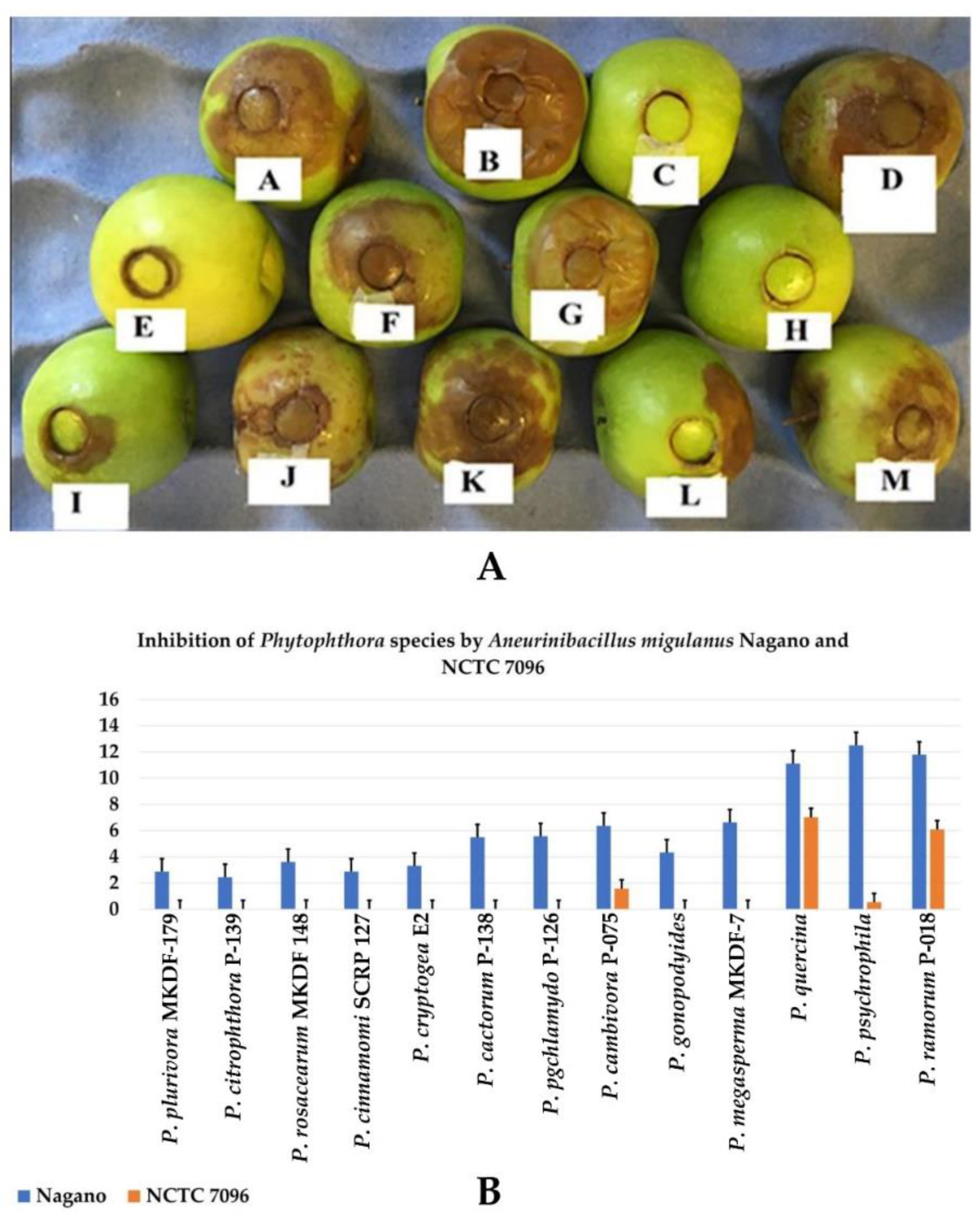
3.4. Effect of A. migulanus Nagano and NCTC 7096 Strains on Apple Infection of Phytophthora Species
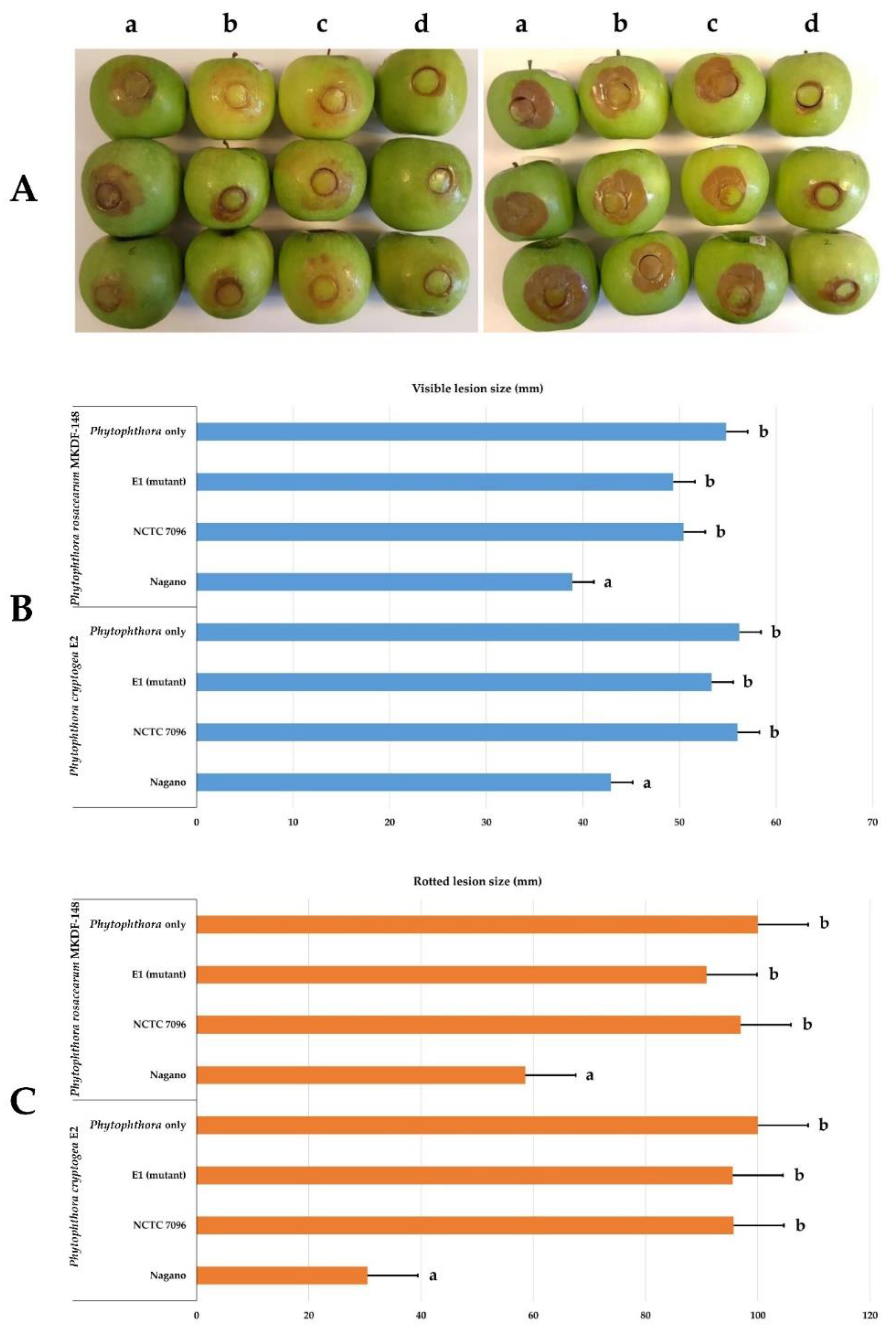
3.5. Effect of A. migulanus Nagano, NCTC 7096, and E1 Mutant Strains on Apple Infection of Phytophthora Rosacearum and Phytophthora Cryptogea Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Belbahri, L.; Lara, E. SSU rRNA reveals major trends in oomycete evolution. Fungal Divers. 2011, 49, 93–100. [Google Scholar] [CrossRef]
- Mefteh, F.B.; Daoud, A.; Chenari Bouket, A.; Alenezi, F.N.; Luptakova, L.; Rateb, M.E.; Kadri, A.; Gharsallah, N.; Belbahri, L. Fungal Root Microbiome from Healthy and Brittle Leaf Diseased Date Palm Trees (Phoenix dactylifera L.) Reveals a Hidden Untapped Arsenal of Antibacterial and Broad Spectrum Antifungal Secondary Metabolites. Front. Microbiol. 2017, 8, 307. [Google Scholar] [CrossRef]
- Cherrad, S.; Charnay, A.; Hernandez, C.; Steva, H.; Belbahri, L.; Vacher, S. Emergence of boscalid-resistant strains of Erysiphe necator in French vineyards. Microbiol. Res. 2018, 216, 79–84. [Google Scholar] [CrossRef]
- Slama, H.; Cherif-Silini, H.; Chenari Bouket, A.; Qader, M.; Silini, A.; Yahiaoui, B.; Alenezi, F.; Luptakova, L.; Triki, M.; Vallat, A.; et al. Screening for Fusarium antagonistic bacteria from contrasting niches designated the endophyte Bacillus halotolerans as plant warden against Fusarium. Front. Microbiol. 2019, 9, 3236. [Google Scholar] [CrossRef] [Green Version]
- Slama, H.B.; Triki, M.A.; Chenari Bouket, A.; Ben Mefteh, F.; Alenezi, F.N.; Luptakova, L.; Cherif-Silini, H.; Vallat, A.; Oszako, T.; Gharsallah, N.; et al. Screening of the High-Rhizosphere Competent Limoniastrum monopetalum’ Culturable Endophyte Mi-crobiota Allows the Recovery of Multifaceted and Versatile Biocontrol Agents. Microorganisms 2019, 7, 249. [Google Scholar] [CrossRef] [Green Version]
- Silini, H.C.; Silini, A.; Chenari Bouket, A.; Alenezi, F.N.; Luptakova, L.; Bouremani, N.; Nowakowska, J.A.; Oszako, T.; Belbahri, L. Tailoring Next Generation Plant Growth Promoting Microorganisms as Versatile Tools beyond Soil Desalinization: A Road Map towards Field Application. Sustainability 2021, 13, 4422. [Google Scholar] [CrossRef]
- Solla, A.; Moreno, G.; Malewski, T.; Jung, T.; Klisz, M.; Tkaczyk, M.; Siebyla, M.; Pérez, A.; Cubera, E.; Hrynyk, H.; et al. Phosphite Spray for the Control of Oak Decline Induced by Phytophthora in Europe. For. Ecol. Manag. 2021, 485, 118938. [Google Scholar] [CrossRef]
- Balla, A.; Silini, A.; Cherif-Silini, H.; Chenari Bouket, A.; Moser, W.K.; Nowakowska, J.A.; Oszako, T.; Benia, F.; Belbahri, L. The Threat of Pests and Pathogens and the Potential for Biological Control in Forest Ecosystems. Forests 2021, 12, 1579. [Google Scholar] [CrossRef]
- Silini, H.C.; Thissera, B.; Chenari Bouket, A.; Saadaoui, N.; Silini, A.; Eshelli, M.; Alenezi, F.N.; Vallat, A.; Luptakova, L.; Yahiaoui, B.; et al. Durum Wheat Stress Tolerance Induced by Endophyte Pantoea agglomerans with Genes Contributing to Plant Functions and Secondary Metabolite Arsenal. Int. J. Mol. Sci. 2019, 20, 3989. [Google Scholar] [CrossRef] [Green Version]
- Cheffi, M.; Chenari Bouket, A.; Alenezi, F.N.; Luptakova, L.; Belka, M.; Vallat, A.; Rateb, M.E.; Tounsi, S.; Triki, M.A.; Belbahri, L. Olea europaea L. Root Endophyte Bacillus velezensis OEE1 Counteracts Oomycete and Fungal Harmful Pathogens and Harbours a Large Repertoire of Secreted and Volatile Metabolites and Beneficial Functional Genes. Microorganisms 2019, 7, 314. [Google Scholar] [CrossRef] [Green Version]
- Dhouib, H.; Zouari, I.; Abdallah, D.B.; Belbahri, L.; Taktak, W.; Triki, M.A.; Tounsi, S. Potential of a novel endophytic Bacillus velezensis in tomato growth promotion and protection against Verticillium wilt disease. Biol. Control 2019, 139, 104092. [Google Scholar] [CrossRef]
- Kerbab, S.; Silini, A.; Chenari Bouket, A.; Cherif-Silini, H.; Eshelli, M.; El Houda Rabhi, N.; Belbahri, L. Mitigation of NaCl Stressin Wheat by Rhizosphere Engineering Using Salt Habitat Adapted PGPR Halotolerant Bacteria. Appl. Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Oszako, T.; Voitka, D.; Stocki, M.; Stocka, N.; Nowakowska, J.A.; Linkiewicz, A.; Hsiang, T.; Belbahri, L.; Berezovska, D.; Malewski, T. Trichoderma asperellum efficiently protects Quercus robur leaves against Erysiphe alphitoides. Eur. J. Plant Pathol. 2021, 159, 295–308. [Google Scholar] [CrossRef]
- Roylawar, P.; Khandagale, K.; Randive, P.; Shinde, B.; Murumkar, C.; Ade, A.; Singh, M.; Gawande, S.; Morelli, M. Piriformospora indica Primes Onion Response against Stemphylium Leaf Blight Disease. Pathogens 2021, 10, 1085. [Google Scholar] [CrossRef]
- Vick, S.H.W.; Fabian, B.K.; Dawson, C.J.; Foster, C.; Asher, A.; Hassan, K.A.; Midgley, D.J.; Paulsen, I.T.; Tetu, S.G. Delving into defence: Identifying the Pseudomonas protegens Pf-5 gene suite involved in defense against secreted products of fungal, oomycete and bacterial rhizosphere competitors. Microb. Genom. 2021, 7, 000671. [Google Scholar] [CrossRef]
- Sandhu, S.K.; Morozov, A.Y.; Holt, R.D.; Barfield, M. Revisiting the Role of Hyperparasitism in the Evolution of Virulence. Am. Nat. 2021, 197, 216–235. [Google Scholar] [CrossRef]
- Alenezi, F.N.; Rekik, I.; Chenari Bouket, A.; Luptakova, L.; Weitz, H.J.; Rateb, M.E.; Jaspars, M.; Woodward, S.; Belbahri, L. Increased Biological Activity of Aneurinibacillus migulanus Strains Correlates with the Production of New Gramicidin Secondary Metabolites. Front. Microbiol. 2017, 8, 517. [Google Scholar] [CrossRef] [Green Version]
- Thissera, B.; Alhadrami, H.A.; Hassan, M.H.A.; Hassan, H.M.; Behery, F.A.; Bawazeer, M.; Yaseen, M.; Belbahri, L.; Rateb, M.E. Induction of Cryptic Antifungal Pulicatin Derivatives from Pantoea Agglomerans by Microbial Co-Culture. Biomolecules 2020, 10, 268. [Google Scholar] [CrossRef] [Green Version]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Belbahri, L.; Alenezi, F.N.; Luptakova, L.; Rateb, M.E.; Woodward, S. Complete genome sequence of Aneurinibacillus migulanus E1, a gramicidin S- and D-phenylalanyl-L-propyl diketopiperazine-deficient mutant. Genome Announc. 2015, 3, e01441-15. [Google Scholar] [CrossRef] [Green Version]
- Slama, H.; Cherif-Silini, H.; Chenari Bouket, A.; Silini, A.; Alenezi, F.N.; Luptakova, L.; Vallat, A.; Belbahri, L. Biotechnology and bioinformatics of endophytes in biocontrol, bioremediation, and plant growth promotion. In Endophytes: Mineral Nutrient Management, Volume 3. Sustainable Development and Biodiversity; Maheshwari, D.K., Dheeman, S., Eds.; Springer: Cham, Switzerland, 2021; Volume 26, pp. 181–205. [Google Scholar]
- Alenezi, F.N.; Fraser, S.; Bełka, M.; Doğmubbbs, T.H.; Hečkova, Z.; Oskay, F.; Belbahri, L.; Woodward, S. Biological control of Dothistroma needle blight on pine with Aneurinibacillus migulanus. For. Pathol. 2016, 46, 555–558. [Google Scholar] [CrossRef]
- Alenezi, F.N.; Rekik, I.; Belka, M.; Ibrahim, A.F.; Luptakova, L.; Woodward, S.; Belbahri, L. Strain-level diversity of secondary metabolism in the biocontrol species Aneurinibacillus migulanus. Microbiol. Res. 2016, 182, 116–124. [Google Scholar] [CrossRef]
- Sellami, M.; Khlifi, A.; Frikha, F.; Miled, N.; Belbahri, L.; Ben Rebah, F. Agro-industrial waste-based growth media optimization for biosurfactant production by Aneurinibacillus migulanus. J. Microbiol. Biotechnol. Food Sci. 2016, 5, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Alenezi, F.N.; Weitz, H.J.; Belbahri, L.; Nidhal, J.; Luptakova, L.; Jaspars, M.; Woodward, S. Draft genome sequence of Aneurinibacillus migulanus NCTC 7096. Genome Announc. 2015, 3, e00234-15. [Google Scholar] [CrossRef] [Green Version]
- Alenezi, F.N.; Weitz, H.J.; Belbahri, L.; Ben Rebah, H.; Luptakova, L.; Jaspars, M.; Woodward, S. Draft genome sequence of Aneurinibacillus migulanus strain nagano. Genome Announc. 2015, 3, e00232-15. [Google Scholar] [CrossRef] [Green Version]
- Shimura, K.; Iwaki, M.; Kanda, M.; Hori, K.; Kaji, E.; Hasegawa, S.; Saito, Y. On the enzyme system obtained from some mutants of Bacillus brevis deficient in gramicidin S formation. Biochim. Biophys. Acta 1974, 338, 577–587. [Google Scholar] [CrossRef]
- Belbahri, L.; Calmin, G.; Sanchez-Hernandez, E.; Oszako, T.; Lefort, F. Pythium sterilum sp. nov. isolated from Poland, Spain and France: Its morphology and molecular phylogenetic position. FEMS Microbiol. Lett. 2006, 255, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Prospero, S.; Vercauteren, A.; Heungens, K.; Belbahri, L.; Rigling, D. Phytophthora diversity and the population structure of Phytophthora ramorum in Swiss ornamental nurseries. Plant Pathol. 2013, 62, 1063–1071. [Google Scholar] [CrossRef]
- Naseri, B.; Younesi, H. Beneficial microbes in biocontrol of root rots in bean crops: A meta-analysis (1990–2020). Physiol. Mol. Plant Pathol. 2021, 116, 101712. [Google Scholar] [CrossRef]
- Belbahri, L.; Chenari Bouket, A.; Rekik, I.; Alenezi, F.N.; Vallat, A.; Luptakova, L.; Petrovova, E.; Oszako, T.; Cherrad, S.; Vacher, S.; et al. Comparative Genomics of Bacillus amyloliquefaciens Strains Reveals a Core Genome with Traits for Habitat Adaptation and a Secondary Metabolites Rich Accessory Genome. Front. Microbiol. 2017, 8, 1438. [Google Scholar] [CrossRef]
- Mefteh, F.B.; Daoud, A.; Chenari Bouket, A.; Thissera, B.; Kadri, Y.; Cherif-Silini, H.; Eshelli, M.; Alenezi, F.N.; Vallat, A.; Oszako, T.; et al. Date Palm Trees Root-Derived Endophytes as Fungal Cell Factories for Diverse Bioactive Metabolites. Int. J. Mol. Sci. 2018, 19, 1986. [Google Scholar] [CrossRef] [Green Version]
- Thissera, B.; Hallyburton, I.; Ngwa, C.J.; Cherif-Silini, H.; Hassane, A.S.I.; Anderson, M.; Campbell, L.A.; Mutter, N.; Eshelli, M.; Abdelmohsen, U.R.; et al. Potent antiplasmodial alkaloids from the rhizobacterium Pantoea agglomerans as hemozoin modulators. Bioorg. Chem. 2021, 115, 105215. [Google Scholar] [CrossRef]
- Biessy, A.; Filion, M. Biological control of potato common scab by plant-beneficial bacteria. Biol. Control 2022, 165, 104808. [Google Scholar] [CrossRef]
- Qiu, Y.; Yan, H.-H.; Sun, S.-M.; Wang, Y.-Q.; Zhao, X.-R.; Wang, H.-Y. Use of Bacillus velezensis SDTB022 against tobacco black shank (TBS) and the biochemical mechanism involved. Biol. Control 2022, 165, 104785. [Google Scholar] [CrossRef]
- Dimkic, I.; Janakiev, T.; Petrovic, M.; Degrassi, G.; Fira, D. Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms—A review. Physiol. Mol. Plant Pathol. 2022, 117, 101754. [Google Scholar] [CrossRef]
- Schuster, C.; Schmitt, A. Efficacy of a bacterial preparation of Aneurinibacillus migulanus against downy mildew of cucumber (Pseudoperonospora cubensis). Eur. J. Plant Pathol. 2017, 151, 439–450. [Google Scholar] [CrossRef]
- Seddon, B.; Edwards, S.; Markellou, E.; Malathrakis, N. Bacterial antagonist-fungal pathogen interactions on the plant aerial surface. Multitrophic Interact. Terr. Syst. 1997, 1, 5–25. [Google Scholar]
- Seddon, B.; McHugh, R.; Schmitt, A. Brevibacillus brevis—A novel candidate biocontrol agent with broad-spectrum antifungal activity. In Proceedings of the BCPC Conference: Pests and Diseases. Proceedings of the International Conference Held at the Brighton Hilton Metropole Hotel, Brighton, UK, 13–16 November 2000; Volume 2. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Clade | Phytophthora Species | Strain | Origin |
|---|---|---|---|
| 1 | P. cactorum | P-138 | Kent |
| 2 | P. plurivora P. citrophthora | MKDF-179 P-139 | Macedonia Kent |
| 3 | P. psychrophila | — | Spain |
| 4 | P. quercina | — | Spain |
| 6 | P. taxonPgchlamydo P.gonapodyides P. megasperma P. rosacearum | P-126 — MKDF-7 MKDF-148 | Kent Spain Macedonia Macedonia |
| 7 | P. cinnamomi P. cambivora | SCRP 127 P-075 | Dundee Unknown |
| 8 | P. cryptogea P. ramorum | E2 P-018 | Aberdeen Kent |
| Title | Nterm_mod | aa1 | aa2 | aa3 | aa4 | aa5 | aa6 | aa7 | aa8 | aa9 | aa10 | adduct |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Term | L | F | P | V | K | L | F | P | V | K | [M + H]+ | |
| Mass | 1.007825 | 113.0841 | 147.0684 | 97.05276 | 99.06841 | 128.095 | 113.0841 | 147.0684 | 97.05276 | 99.06841 | 128.09496 | 1169.745 |
| b-series | 1.007825 | 114.0919 | 261.1603 | 358.2131 | 457.2815 | 585.3764 | 698.4605 | 845.5289 | 942.5817 | 1041.65 | 1169.745 | |
| Actual mass | - | 261.1606 | 358.2188 | 457.2803 | 585.3751 | 698.4605 | 845.5264 | 942.5762 | 1041.6481 | |||
| y-series | 1.007825 | 1169.745 | 1056.661 | 909.5926 | 812.5398 | 713.4714 | 585.3764 | 472.2924 | 325.224 | 228.1712 | 129.102785 | |
| Actual mass | - | 1056.6727 | 909.5892 | 812.5347 | 713.4695 | 585.3751 | 472.2917 | 325.2228 | 228.1687 | |||
| Nterm_mod | aa1 | aa2 | aa3 | aa4 | Orn | aa6 | aa7 | aa8 | aa9 | aa10 | adduct | |
| Term | L | F | P | V | K | L | F | P | V | K | [M + H]+ | |
| Mass | 1.007825 | 113.0841 | 147.0684 | 97.05276 | 99.06841 | 114.0793 | 113.0841 | 147.0684 | 97.05276 | 99.06841 | 128.09496 | 1155.729 |
| b-series | 1.007825 | 114.0919 | 261.1603 | 358.2131 | 457.2815 | 571.3608 | 684.4448 | 831.5132 | 928.566 | 1027.634 | 1155.7294 | |
| Actual mass | - | 261.1598 | 358.2122 | 457.2795 | 571.3612 | 684.4433 | 831.5117 | 928.5595 | 1027.6339 | |||
| y-series | 1.007825 | 1155.729 | 1042.645 | 895.5769 | 798.5241 | 699.4557 | 585.3764 | 472.2924 | 325.224 | 228.1712 | 129.102785 | |
| Actual mass | 1042.6502 | 895.5762 | 798.5214 | 699.4553 | 585.3752 | 472.2917 | 325.2232 | 228.1713 | ||||
| Nterm_mod | aa1 | aa2 | aa3 | aa4 | Orn | aa6 | aa7 | aa8 | aa9 | Orn | adduct | |
| Term | L | F | P | V | K | L | F | P | V | K | [M + H]+ | |
| Mass | 1.007825 | 113.0841 | 147.0684 | 97.05276 | 99.06841 | 114.0793 | 113.0841 | 147.0684 | 97.05276 | 99.06841 | 113.93846 | 1141.573 |
| b-series | 1.007825 | 114.0919 | 261.1603 | 358.2131 | 457.2815 | 571.3608 | 684.4448 | 831.5132 | 928.566 | 1027.634 | 1141.5729 | |
| Actual mass | 261.1598 | 358.2108 | 457.2784 | 571.3607 | 684.4442 | 831.5147 | 928.5653 | 1027.6261 | ||||
| y-series | 1.007825 | 1141.714 | 1028.63 | 881.5613 | 784.5085 | 685.4401 | 571.3608 | 458.2767 | 311.2083 | 214.1555 | 115.087135 | |
| Actual mass | 1028.6317 | 881.5618 | 784.5082 | 685.4402 | 571.3607 | 458.2810 | 311.2080 | 214.1554 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alenezi, F.N.; Bouket, A.C.; Cherif-Silini, H.; Silini, A.; Jaspars, M.; Oszako, T.; Belbahri, L. Loss of Gramicidin Biosynthesis in Gram-Positive Biocontrol Bacterium Aneurinibacillus migulanus (Takagi et al., 1993) Shida et al. 1996 Emend Heyndrickx et al., 1997 Nagano Impairs Its Biological Control Ability of Phytophthora. Forests 2022, 13, 535. https://doi.org/10.3390/f13040535
Alenezi FN, Bouket AC, Cherif-Silini H, Silini A, Jaspars M, Oszako T, Belbahri L. Loss of Gramicidin Biosynthesis in Gram-Positive Biocontrol Bacterium Aneurinibacillus migulanus (Takagi et al., 1993) Shida et al. 1996 Emend Heyndrickx et al., 1997 Nagano Impairs Its Biological Control Ability of Phytophthora. Forests. 2022; 13(4):535. https://doi.org/10.3390/f13040535
Chicago/Turabian StyleAlenezi, Faizah N., Ali Chenari Bouket, Hafsa Cherif-Silini, Allaoua Silini, Marcel Jaspars, Tomasz Oszako, and Lassaȃd Belbahri. 2022. "Loss of Gramicidin Biosynthesis in Gram-Positive Biocontrol Bacterium Aneurinibacillus migulanus (Takagi et al., 1993) Shida et al. 1996 Emend Heyndrickx et al., 1997 Nagano Impairs Its Biological Control Ability of Phytophthora" Forests 13, no. 4: 535. https://doi.org/10.3390/f13040535