
Thinning Effects on Stand Structure and Carbon Content of Secondary Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
2.3.1. DBH Structure Analysis
2.3.2. Spatial Structure Analysis
2.3.3. Carbon Analysis
3. Results
3.1. Effects of Thinning on DBH Structure
3.2. Effects of Thinning on Spatial Structure
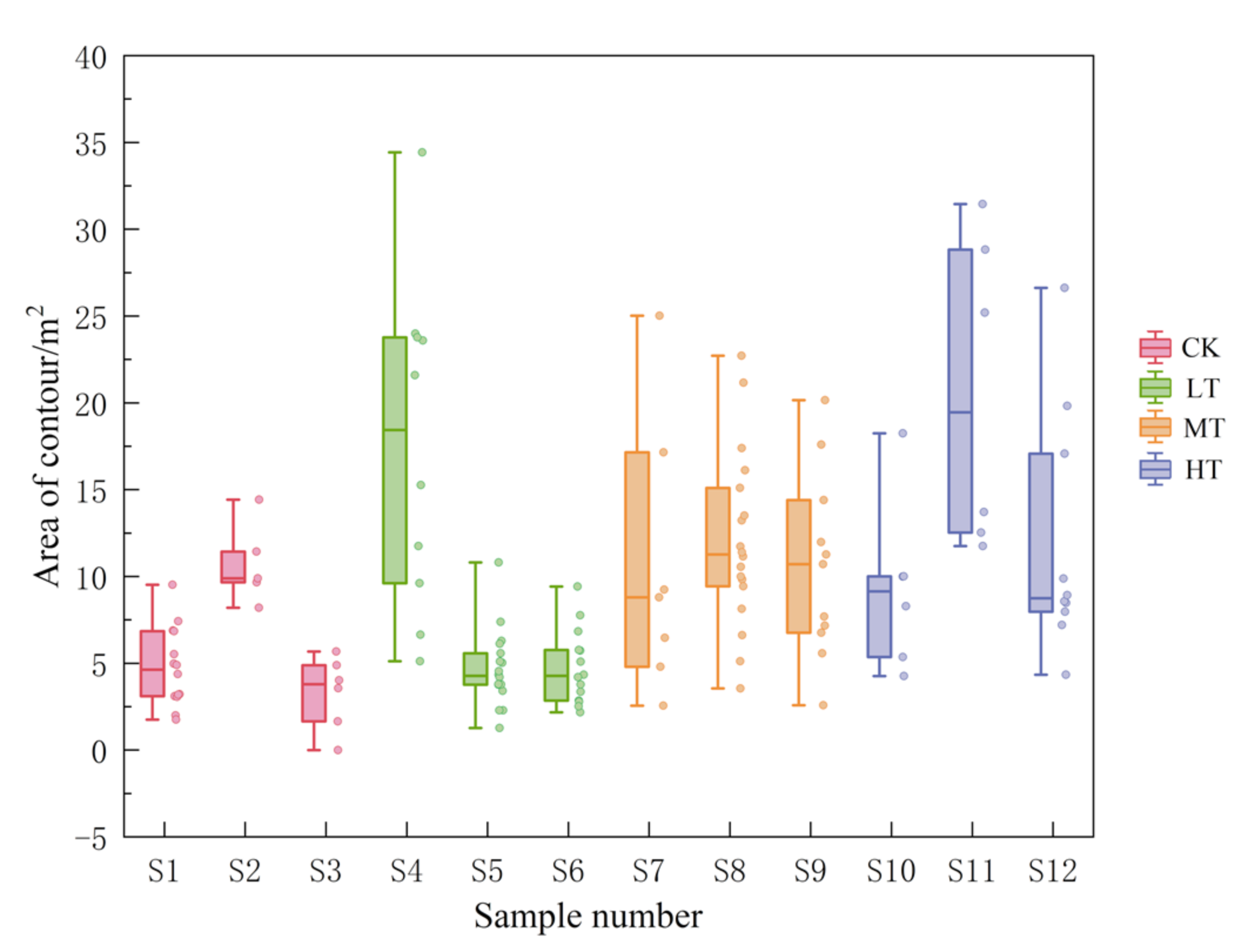
3.2.1. Effects of Thinning on Crown Area
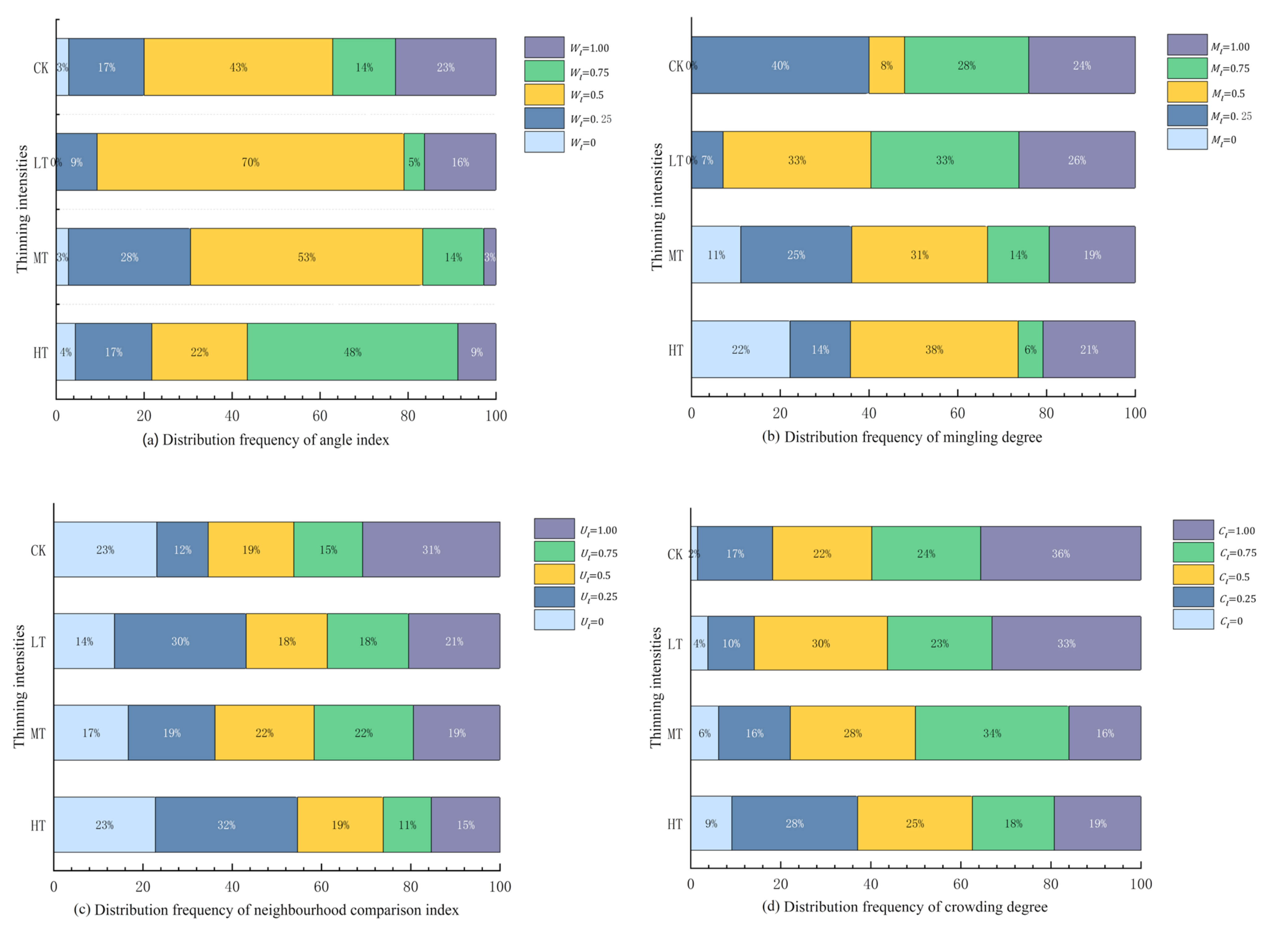
3.2.2. Effects of Thinning on Spatial Structure Index
3.3. Relationship between Spatial Structure Parameters and Carbon Stock under Different Treatments
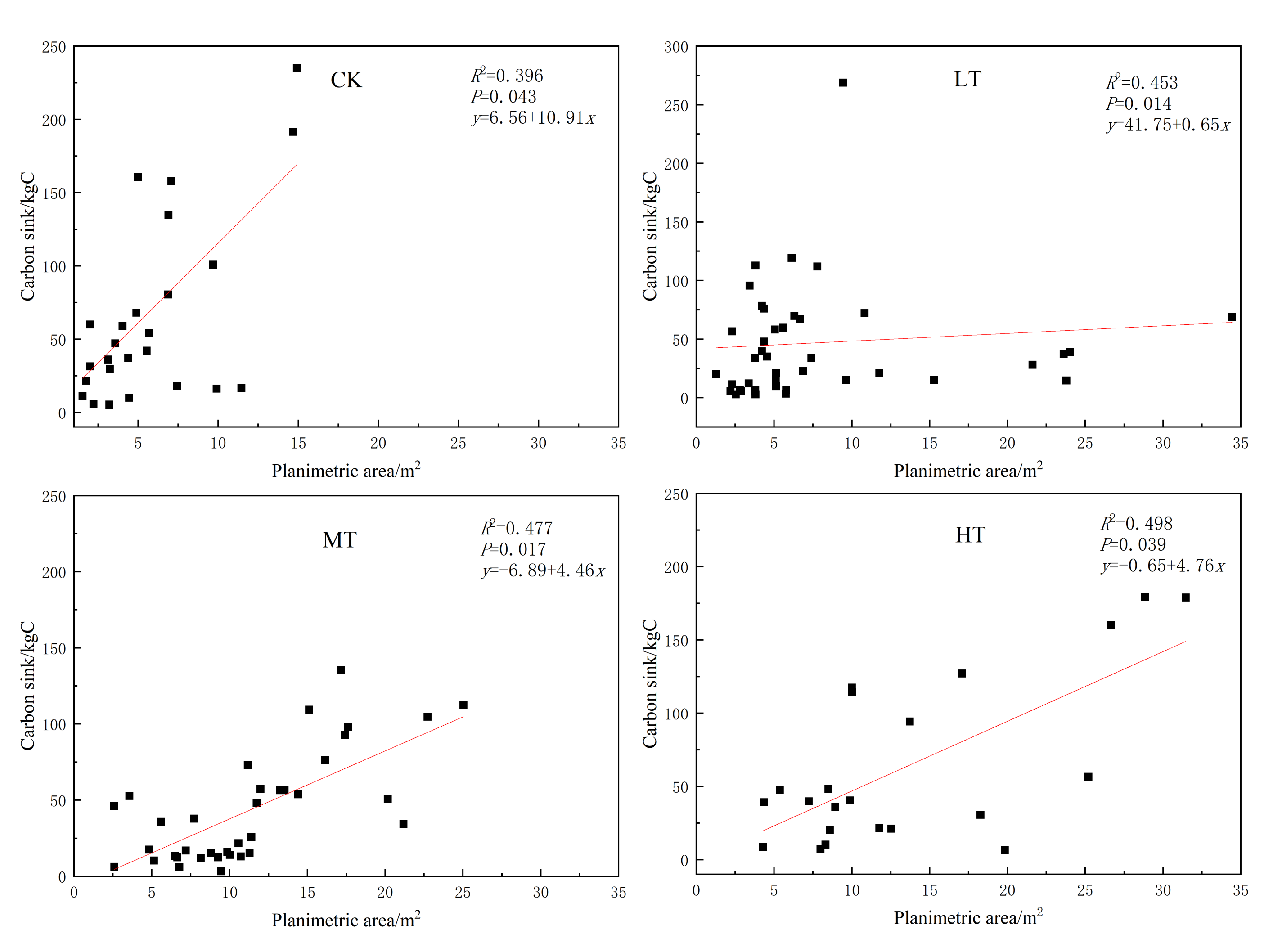
3.3.1. Relationship between Crown Area and Carbon Stock
3.3.2. Relationship between Spatial Structure Index and Carbon Stock
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pastorella, F.; Paletto, A. Stand structure indices as tools to support forest management: An application in Trentino forests (Italy). J. For. Sci. 2013, 59, 159–168. [Google Scholar] [CrossRef] [Green Version]
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and woodland stand structural complexity: Its definition and measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H. Stand Structural Dynamics of North American Boreal Forests. Crit. Rev. Plant Sci. 2006, 25, 115–137. [Google Scholar] [CrossRef]
- DeWalt, S.J.; Maliakal, S.K.; Denslow, J.S. Changes in vegetation structure and composition along a tropical forest chronosequence: Implications for wildlife. For. Ecol. Manag. 2003, 182, 139–151. [Google Scholar] [CrossRef]
- Rahman, M.M.; Nishat, A.; Rahman, G.M.M.; Ruprecht, H.; Vacik, H. Analysis of spatial diversity of sal (Shorea robusta Gaertn.f) forests using neighbourhood-based measures. Community Ecol. 2008, 9, 193–199. [Google Scholar] [CrossRef]
- Zhang, M.M.; Fan, S.H.; Yan, X.R.; Zhou, Y.Q.; Guan, F.Y. Relationships between stand spatial structure characteristics and influencing factors of bamboo and broad-leaved mixed forest. J. For. Res. 2020, 25, 83–91. [Google Scholar] [CrossRef]
- Kohyama, T.; Aiba, S. Dynamics of primary and secondary warm-temperate rain forests in Yakushima Island. Tropics 1997, 6, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zeng, F.P.; Song, T.Q.; Wang, K.L.; Du, H. Stand Structure and Abiotic Factors Modulate Karst Forest Biomass in Southwest China. Forests 2020, 11, 443. [Google Scholar] [CrossRef] [Green Version]
- Neil, G.; Williams, M.D.P. Carbon storage implications of active management in mature Pseudotsuga menziesii forests of western Oregon. For. Ecol. Manag. 2019, 432, 761–775. [Google Scholar] [CrossRef]
- Huang, X.M.; Liu, S.R.; Wang, H.; Hu, Z.D.; Li, Z.G.; You, Y.M. Changes of soil microbial biomass carbon and community composition through mixing nitrogen-fixing species with Eucalyptus urophylla in subtropical China. Soil Biol. Biochem. 2014, 73, 42–48. [Google Scholar] [CrossRef]
- Ali, A.; Lin, S.L.; He, J.K.; Kong, F.M.; Yu, J.H.; Jiang, H.S. Climate and soils determine aboveground biomass indirectly via species diversity and stand structural complexity in tropical forests. For. Ecol. Manag. 2019, 432, 823–831. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J. A Review of Processes Behind Diversity—Productivity Relationships in Forests. Curr. For. Rep. 2016, 2, 45–61. [Google Scholar] [CrossRef] [Green Version]
- Montoro, G.M.; Morin, H.; Lussier, J.M.; Walsh, D. Radial growth response of black spruce stands ten years after experimental shelterwoods and seed-tree cuttings in boreal forest. Forests 2016, 7, 240. [Google Scholar] [CrossRef] [Green Version]
- Moussaoui, L.; Leduc, A.; Girona, M.M.; Bélisle, A.C.; Lafleur, B.; Fenton, N.J.; Bergeron, Y. Success factors for experimental partial harvesting in unmanaged boreal forest: 10-year stand yield results. Forests 2020, 11, 1199. [Google Scholar] [CrossRef]
- Montoro, G.M.; Lussier, J.M.; Morin, H.; Thiffault, N. Conifer regeneration after experimental shelterwood and seed-tree treatments in boreal forests: Finding silvicultural alternatives. Front. Plant Sci. 2018, 9, 1145. [Google Scholar] [CrossRef]
- Montoro, G.M.; Morin, H.; Lussier, J.M.; Ruel, J.C. Post-cutting mortality following experimental silvicultural treatments in unmanaged boreal forest stands. Front. For. Glob. Change 2018, 9, 1145. [Google Scholar] [CrossRef] [Green Version]
- Bose, A.K.; Weiskittel, A.; Kuehne, C.; Wagner, R.G.; Turnblom, E.; Burkhart, H.E. Tree-level growth and survival following commercial thinning of four major softwood species in North America. For. Ecol. Manag. 2018, 427, 355–364. [Google Scholar] [CrossRef]
- Kim, S.; Axelsson, E.P.; Girona, M.M.; Senior, J.K. Continuous-cover forestry maintains soil fungal communities in Norway spruce dominated boreal forests. For. Ecol. Manag. 2021, 480, 118659. [Google Scholar] [CrossRef]
- Liu, C.X.; Wang, Y.J.; Ma, C.; Wang, Y.Q.; Zhang, H.L.; Hu, B. Quantifying the effect of non-spatial and spatial forest stand structure on rainfall partitioning in mountain forests, Southern China. For. Chron. 2018, 94, 162–172. [Google Scholar] [CrossRef]
- Zhao, Q.Y.; Yu, K.Y.; Xiang, J.; Lin, L.L.; Huang, H.; Xie, Q.Y.; Liu, J. Influence of forest park stand structure characteristics on spatial thermal environment. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2019, 48, 48–54. (In Chinese) [Google Scholar]
- Wang, Z.C.; Li, Y.X.; Meng, Y.B.; Wang, C. Effect of tending and thinning on spatial and carbon distribution patterns of natural mixed broadlesf-conifer secondary forest in Xiaoxing’an Mountains, PR China. Appl. Ecol. Environ. Res. 2021, 19, 4751–4764. [Google Scholar] [CrossRef]
- Wang, Z.C.; Li, Y.X.; Meng, Y.B.; Wang, C. Responses of spatial distribution patterns and associations of Larix gmelinii and Populus davidiana mixed forests in Daxing’an Mountains to different tending thinning intensities. J. Cent. South Univ. For. Technol. 2022, 42, 75–83. (In Chinese) [Google Scholar]
- Zhang, T.; Zhu, Y.J.; Dong, X.B. Effects of thinning on the habitat of natural mixed broadleaf-conifer secondary forest in Xiaoxing’an Mountains of northeastern China. J. Beijing For. Univ. 2017, 39, 1–12. (In Chinese) [Google Scholar]
- Saka, M.G. Characterization of diameter distribution and prediction of Weibull parameters equation for plantation-grown Eucalyptus species. Asian J. Res. Agric. For. 2021, 7, 1–13. [Google Scholar] [CrossRef]
- Ganbaatar, B.; Jamsran, T.; Gradel, A.; Sukhbaatar, G. Assessment of the effects of thinnings in scots pine plantations in Mongolia: A comparative analysis of tree growth and crown development based on dominant trees. For. Sci. Technol. 2021, 17, 135–143. [Google Scholar] [CrossRef]
- Hui, G.Y. The neighbourhood pattern—A new structure parameter describing distribution of forest tree position. Sci. Silvae Sin. 1999, 35, 37–42. (In Chinese) [Google Scholar]
- Hui, G.Y.; Hu, Y.B. Measuring species spatial isolation in mixed forests. For. Res. 2001, 14, 23–27. (In Chinese) [Google Scholar]
- Hui, G.Y.; Hu, Y.B.; Zhao, Z.H. Evaluating tree species segregation based on neighborhood spatial relationships. J. Beijing For. Univ. 2008, 30, 131–134. (In Chinese) [Google Scholar]
- Hui, G.Y.; von Gadow, K.; Albert, M. A new parameter for stand spatial structure neighbourhood comparison. For. Res. 1999, 12, 1–6. (In Chinese) [Google Scholar]
- Hu, Y.B.; Hui, G.Y. How to describe the crowding degree of trees based on the relationship of neighboring trees. J. Beijing For. Univ. 2015, 37, 1–8. (In Chinese) [Google Scholar]
- Hu, H.Q.; Luo, B.Z.; Wei, S.J.; Wei, S.W.; Sun, L.; Luo, S.S.; Ma, H.B. Biomass carbon density and carbon sequestration capacity in seven typical forest types of the Xiaoxing’an Mountains, China. Chin. J. Plant Ecol. 2015, 39, 140–158. (In Chinese) [Google Scholar]
- Wang, C.K. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For. Ecol. Manag. 2006, 222, 9–16. [Google Scholar] [CrossRef]
- Zhang, L.J.; Liu, C.M. Fitting irregular diameter distributions of forest stands by Weibull, modified Weibull, and mixture Weibull models. J. For. Res. 2006, 11, 369–372. [Google Scholar] [CrossRef]
- Zhang, M.T.; Kang, X.G.; Guo, W.W.; Meng, J.H.; Yang, Y.J. Diameter structural distribution of spruce-fir mixed forest in Changbai Mountain. J. Northwest A F Univ. Nat. Sci. Ed. 2015, 43, 65–72. (In Chinese) [Google Scholar]
- Poudel, K.P.; Cao, Q.V. Evaluation of Methods to Predict Weibull Parameters for Characterizing Diameter Distributions. For. Sci. 2013, 59, 243–252. [Google Scholar] [CrossRef]
- Philip, G.C. Effects of Aspen and spruce density on size and number of lower branches 20 years after thinning of two boreal mixedwood stands. Forests 2021, 12, 211. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Hui, G.Y.; Zhao, Z.H.; Hu, Y.B.; Wang, H.X.; Liu, W.Z.; Zang, R.G. Composition of basal area in natural forests based on the uniform angle index. Ecol. Inform. 2018, 45, 1–8. [Google Scholar] [CrossRef]
- Zhang, T.; Dong, X.B.; Guan, H.W.; Meng, Y.; Ruan, J.F.; Wang, Z.Y. Effect of Thinning on the Spatial Structure of a Larix gmelinii Rupr. Secondary Forest in the Greater Khingan Mountains . Forests 2018, 9, 720. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.X.; Zheng, Z.R.; Diao, Z.Y.; Ding, G.D.; Bao, Y.F.; Liu, Y.D.; Gao, G.L. Effects of thinning on the spatial structure of Larixprincipis-rupprechtii plantation. Sustainability 2018, 10, 1250. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H. Mixing degree, stand density, and water supply can increase the overyielding of mixed versus mono specific stands in Central Europe. For. Ecol. Manag. 2021, 503, 119741. [Google Scholar] [CrossRef]
- Fotis, A.T.; Murphy, S.J.; Ricart, R.D.; Krishnadas, M.; Whitacre, J.; Wenzel, J.W.; Queenborough, S.A.; Comita, L.S. Above-ground biomass is driven by mass-ratio effects and stand structural attributes in a temperate deciduous forest. J. Ecol. 2018, 106, 561–570. [Google Scholar] [CrossRef]
- Capellesso, E.S.; Scrovonski, K.L.; Zanin, E.M.; Hepp, L.U.; Bayer, C.; Sausen, T.L. Effects of forest structure on litter production, soil chemical composition and litter-soil interactions. Acta Bot. Bras. 2016, 30, 329–335. [Google Scholar] [CrossRef]
- Martin, M.; Raymond, P. Assessing tree-related microhabitat retention according to a harvest gradient using tree-defect surveys as proxies in Eastern Canadian mixedwood forests. For. Chron. 2019, 95, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Cui, S.; Xiao, R.; Wang, W.F.; Liu, B.F. The study on the structural characteristics of the mixed-wood carbon sink in Xiaoxinganling Area. For. Eng. 2020, 36, 30–35. (In Chinese) [Google Scholar]
- Ameray, A.; Bergeron, Y.; Valeria, O.; Montoro, G.M.; Cavard, X. Forest Carbon Management: A Review of Silvicultural Practices and Management Strategies Across Boreal, Temperate and Tropical Forests. Curr. For. Rep. 2021, 7, 245–266. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thinning Treatment | Elevation/m | Species | Diameter at Breast Height/cm | Tree Height/m | Stand Density/Plant·hm−2 | Canopy Density/% | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Average | Max | Min | Average | |||||
| CK | 456.8~492.1 | 2A.m:2A.t:1P.a:1A.f:1F:1P.k:1P.L:1P.a-T-U-A.u | 40.7 | 5.1 | 16.4 | 21.9 | 4.3 | 11.6 | 958 | 0.90 |
| LT | 504~509 | 3A.t:2A.f:1P.a:1A.m:1F:2P.k-P.L-T-U | 42.7 | 5.1 | 13.5 | 22.6 | 3.9 | 9.6 | 1183 | 0.92 |
| MT | 487~540 | 3A.m:2A.t:2A.f:1P.a1P.p:1F-P.L-B-T | 35.1 | 5.2 | 16.4 | 20.8 | 4.3 | 11.5 | 895 | 0.81 |
| HT | 468.2~499 | 3A.m:2A.t:2A.f:1P.a:1P.L:1T-U-P.p-A.u | 42.8 | 5.3 | 17.1 | 23.6 | 5.3 | 11.7 | 813 | 0.76 |
| Parameter | Formulations | Specific Meanings | ||||
|---|---|---|---|---|---|---|
| W-index |  = 0 Absolutely uniform |  = 0.25 = 0.25Uniform |  = 0.5 Random |  = 0.75 Nonuniform |  = 1 Clumped | |
| M-index |  = 0 Zero degree |  = 0.25 Weak degree |  = 0.5 Moderate degree |  = 0.75 = 0.75Strong degree |  = 1 Extremely strong degree | |
| U-index |  = 0 Advantage |  = 0.25 Subadvantage |  = 0.5 Moderate |  = 0.75 Disadvantage |  = 1 Absolute disadvantage | |
| C-index |  = 0 Extremely sparse |  = 0.25 Sparse |  = 0.5 Moderately dense |  = 0.75 Relatively dense |  = 1 Extremely dense | |
 stands for different tree species;
stands for different tree species;  represents different DBH size;
represents different DBH size;  stands for basal area and
stands for basal area and  stands for crown area.
stands for crown area.| Spatial Structure Index | Thinning Treatment | Number | Result | |
|---|---|---|---|---|
| r Value | p Value | |||
| W-index | CK | 37 | 0.073 | 0.251 |
| LT | 52 | 0.180 | 0.364 | |
| MT | 46 | 0.441 | 0.000 | |
| HT | 32 | 0.367 | 0.006 | |
| M-index | CK | 37 | 0.096 | 0.268 |
| LT | 52 | 0.379 | 0.000 | |
| MT | 46 | 0.167 | 0.002 | |
| HT | 32 | 0.198 | 0.006 | |
| U-index | CK | 37 | −0.160 | 0.027 |
| LT | 52 | −0.273 | 0.000 | |
| MT | 46 | −0.194 | 0.002 | |
| HT | 32 | −0.072 | 0.000 | |
| C-index | CK | 37 | −0.190 | 0.001 |
| LT | 52 | −0.056 | 0.000 | |
| MT | 46 | −0.244 | 0.000 | |
| HT | 32 | 0.272 | 0.006 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Li, Y.; Meng, Y.; Li, C.; Zhang, Z. Thinning Effects on Stand Structure and Carbon Content of Secondary Forests. Forests 2022, 13, 512. https://doi.org/10.3390/f13040512
Wang Z, Li Y, Meng Y, Li C, Zhang Z. Thinning Effects on Stand Structure and Carbon Content of Secondary Forests. Forests. 2022; 13(4):512. https://doi.org/10.3390/f13040512
Chicago/Turabian StyleWang, Zichun, Yaoxiang Li, Yongbin Meng, Chunxu Li, and Zheyu Zhang. 2022. "Thinning Effects on Stand Structure and Carbon Content of Secondary Forests" Forests 13, no. 4: 512. https://doi.org/10.3390/f13040512