
Ecological Stoichiometry in Pinus massoniana L. Plantation: Increasing Nutrient Limitation in a 48-Year Chronosequence

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Material and Methods
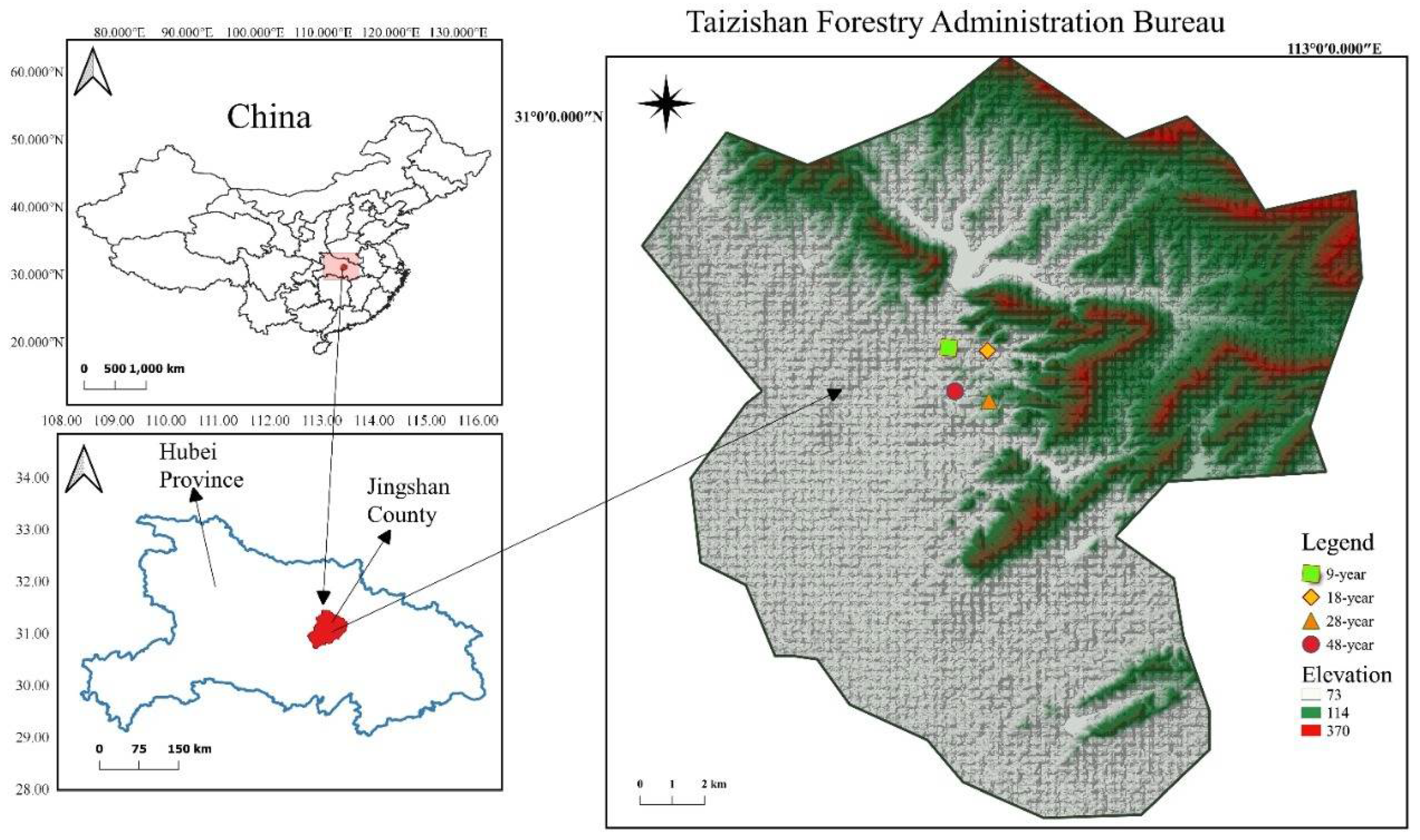
2.1. Study Area and Plantation
2.2. Plant Sampling Design and Data Collection
2.3. Soil Sample and Analysis
2.4. Data Analysis
3. Results
3.1. Changes in Soil Properties among Stand Ages
3.2. Soil C:N:P and Stand Age
3.3. C:N:P and Stand Age
3.4. Nutrient Resorption Efficiency (NuRE) and the Relationship among Stoichiometry Ratios
3.5. Response of CNP Stoichiometry to Different Stand Ages
4. Discussion
4.1. Ecological Stoichiometry and Stand Age
4.2. Stand Age and Plant Component C:N:P Stoichiometry
4.3. Leaf C, N, P Status
4.4. NRE and PRE in Different Age Groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ågren, G.I. Stoichiometry and nutrition of plant growth in natural communities. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 153–170. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Penuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 33–47. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.; Cleland, E.E.; Grunner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güsewell, S. N: P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.Y.; Lin, K.C.; Vadeboncoeur, M.A.; Chen, M.Z.; Huang, M.Y.; Lin, T.C. Understorey plant community and light availability in conifer plantations and natural hardwood forests in Taiwan. Appl. Veg. Sci. 2015, 18, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, W.; Xu, M.; Deng, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Response of forest growth to C: N: P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 2019, 337, 280–289. [Google Scholar] [CrossRef]
- Qiu, X.; Wang, H.; Peng, D.; Liu, X.; Yang, F.; Li, Z.; Cheng, S. Thinning drives C:N:P stoichiometry and nutrient resorption in Larix principis-rupprechtii plantations in North China. For. Ecol. Manag. 2020, 462, 117984. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, Y. Ecosystem C:N:P stoichiometry and carbon storage in plantations and a secondary forest on the Loess Plateau, China. Ecol. Eng. 2017, 105, 125–132. [Google Scholar] [CrossRef]
- Niu, S.; Ren, L.; Song, L.; Duan, Y.; Huang, T.; Han, X.; Hao, W. Plant stoichiometry characteristics and relationships with soil nutrients in Robinia pseudoacacia communities of different planting ages. Acta Ecol. Sin. 2017, 37, 355–362. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Glob. Ecol. Biogeogr. 2009, 18, 11–18. [Google Scholar] [CrossRef]
- Battaglia, M.A.; Mitchell, R.J.; Mou, P.P.; Pecot, S.D. Light Transmittance Estimates in a longleaf pine woodland. For. Sci. 2003, 49, 752–762. [Google Scholar]
- Jiang, P. C:N:P Stoichiometry and Carbon Storage in a Naturally-Regenerated Secondary Quercus variabilis Forest Age Sequence in the Qinling Mountains, China. Forests 2017, 8, 281. [Google Scholar] [CrossRef]
- Dhiedt, E.; Verheyen, K.; De Smedt, P.; Ponette, Q.; Baeten, L. Early Tree Diversity and Composition Effects on Topsoil Chemistry in Young Forest Plantations Depend on Site Context. Ecosystems 2021, 24, 1638–1653. [Google Scholar] [CrossRef]
- Carnus, J.-M.; Parrotta, J.; Brockerhoff, E.; Arbez, M.; Jactel, H.; Kremer, A.; Lamb, D.; O’Hara, K.; Walters, B. Planted forests and biodiversity. J. For. 2006, 104, 65–77. [Google Scholar] [CrossRef]
- Dangal, S.P.; Das, A.K.; Paudel, S.K. Effectiveness of management interventions on forest carbon stock in planted forests in Nepal. J. Environ. Manag. 2017, 196, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Stefańska-Krzaczek, E.; Szymura, T.H. Species diversity of forest floor vegetation in age gradient of managed scots pine stands. Balt. For. 2015, 21, 233–243. [Google Scholar]
- Sitzia, T.; Trentanovi, G.; Dainese, M.; Gobbo, G.; Lingua, E.; Sommacal, M. Stand structure and plant species diversity in managed and abandoned silver fir mature woodlands. For. Ecol. Manag. 2012, 270, 232–238. [Google Scholar] [CrossRef]
- Jiao, J.; Zhang, Z.; Bai, W.; Jia, Y.; Wang, N. Assessing the Ecological Success of Restoration by Afforestation on the Chinese Loess Plateau. Restor. Ecol. 2012, 20, 240–249. [Google Scholar] [CrossRef]
- Szymura, T.H.; Szymura, M. Soil properties and light availability determine species richness and vegetation diversity in an overgrown coppice oak stand. Polish J. Ecol. 2011, 59, 523–533. [Google Scholar]
- Kumar, P.; Chen, H.Y.H.; Thomas, S.C.; Shahi, C. Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forest of Canada. J. Ecol. 2018, 106, 1266–1276. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, L.; Peng, S.; Penuelas, J.; Zeng, H.; Piao, S. Age-Related Modulation of the Nitrogen Resorption Efficiency Response to Growth Requirements and Soil Nitrogen Availability in a Temperate Pine Plantation. Ecosystems 2016, 19, 698–709. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhao, N.; Liu, C.; Yang, H.; Li, M.; Yu, G.; Wilcox, K.; Yu, Q.; He, N. C:N:P stoichiometry in China’ s forests: From organs to ecosystems. Funct. Ecol. 2018, 32, 50–60. [Google Scholar] [CrossRef]
- Noormets, A.; Epron, D.; Domec, J.C.; McNulty, S.G.; Fox, T.; Sun, G.; King, J.S. Effects of forest management on productivity and carbon sequestration: A review and hypothesis. For. Ecol. Manag. 2014, 355, 124–140. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Ahmad, A.; Akhtar, K.; Teng, M.; Zeng, W.; Yan, Z.; Zhou, Z. Patterns of Biomass, carbon, and soil properties in Masson pine (Pinus massoniana Lamb) plantations with different stand ages and management practices. Forests 2019, 10, 645. [Google Scholar] [CrossRef] [Green Version]
- Ray, G.; Ochoa, C.G.; Deboodt, T.; Mata-Gonzalez, R. Overstory–understory vegetation cover and soil water content observations in western juniper woodlands: A paired watershed study in Central Oregon, USA. Forests 2019, 10, 151. [Google Scholar] [CrossRef] [Green Version]
- Dzwonko, Z.; Gawroński, S. Effect of litter removal on species richness and acidification of a mixed oak-pine woodland. Biol. Conserv. 2002, 106, 389–398. [Google Scholar] [CrossRef]
- Zemunik, G.; Turner, B.L.; Lambers, H.; Laliberté, E. Diversity of plant nutrient-acquisition strategies increases during long-term ecosystem development. Nat. Plants 2015, 1, 15050. [Google Scholar] [CrossRef]
- Bai, Y.F.; Shen, Y.Y.; Jin, Y.D.; Hong, Y.; Liu, Y.Y.; Li, Y.Q.; Liu, R.; Zhang, Z.W.; Jiang, C.Q.; Wang, Y.J. Selective thinning and initial planting density management promote biomass and carbon storage in a chronosequence of evergreen conifer plantations in Southeast China. Glob. Ecol. Conserv. 2020, 24, e01216. [Google Scholar] [CrossRef]
- Wen, L.; Lei, P.; Xiang, W.; Yan, W.; Liu, S. Forest Ecology and Management Soil microbial biomass carbon and nitrogen in pure and mixed stands of Pinus massoniana and Cinnamomum camphora differing in stand age. For. Ecol. Manag. 2014, 328, 150–158. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.; Shao, Y.; Xu, G.; Fu, S. Effects of vegetation removal on soil properties and decomposer organisms. Soil Biol. Biochem. 2011, 43, 954–960. [Google Scholar] [CrossRef]
- Liu, C.L.C.; Kuchma, O.; Krutovsky, K.V. Mixed-species versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Glob. Ecol. Conserv. 2018, 15, e00419. [Google Scholar] [CrossRef]
- Wilson, D.S.; Puettmann, K.J. Density management and biodiversity in young Douglas-fir forests: Challenges of managing across scales. For. Ecol. Manag. 2007, 246, 123–134. [Google Scholar] [CrossRef]
- Yang, H.-X.; Wang, S.-L.; Fan, B.; Zhang, W.-D.; Wei, C.-E. Dynamics of nutrients in an age sequence of Pinus massoniana plantation. Chin. J. Appl. Ecol. 2010, 21, 1907–1914. [Google Scholar]
- Cui, N.J.; Liu, X.B.; ZHANG, J. The distribution pattern of carbon, nitrogen and phosphorus and the stoichimotey characteristics of Pinusmassoniana plantion in different ages. Ecol. Environ. Sci 2014, 23, 188. [Google Scholar]
- State Forestry Administration (SFA). Tree Biomass Methods and Related Parameters of Carbon Accumulating in Pinus Massonina; China Forestry Publisher: Beijing, China, 2014. [Google Scholar]
- Peng, S.L.; Li, Q.F.; Li, D.; Wang, Z.F.; Wang, D.P. Genetic diversity of Pinus massoniana revealed by RAPD markers. Silvae Genet. 2003, 52, 60–62. [Google Scholar]
- Song, Y.-J.; Tian, W.-B.; Liu, X.-Y.; Yin, F.; Cheng, J.-Y.; Zhu, D.-N.; Ali, A.; Yan, E.-R. Associations between litterfall dynamics and micro-climate in forests of Putuoshan Island, Zhejiang, China. Chin. J. Plant Ecol. 2016, 40, 1154. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Wang, S.; Ren, C.; Zhang, W.; Zhao, F.; Li, X. Nitrogen and Phosphorus Resorption in Relation to Nutrition Limitation along the Chronosequence of Black Locust (Robinia pseudoacacia L.) Plantation. Forests 2019, 10, 261. [Google Scholar] [CrossRef] [Green Version]
- Saeed, S.; Yujun, S.; Beckline, M.; Chen, L.; Zhang, B. Forest edge effect on biomass carbon along altitudinal gradients in Chinese Forest edge effect on biomass carbon along altitudinal gradients in Chinese Fir (Cunninghamia lanceolata): A study from Southeastern China. Carbon Manag. 2019, 10, 11–22. [Google Scholar] [CrossRef]
- Liu, W.D.; Su, J.R.; Li, S.F.; Zhang, Z.J.; Li, Z.W. Stoichiometry study of C, N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province. Acta Ecol. Sin. 2010, 30, 6581–6590. [Google Scholar]
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Robert, B. Jackson Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Bao, S.D. Agro-Chemical Analysis of Soil; Agricultural Publish House of China: Beijing, China, 2000. [Google Scholar]
- Ali, A.; Dai, D.; Akhtar, K.; Teng, M.; Yan, Z.; Urbina-Cardona, N.; Mullerova, J.; Zhou, Z. Response of understory vegetation, tree regeneration, and soil quality to manipulated stand density in a Pinus massoniana plantation. Glob. Ecol. Conserv. 2019, 20, e00775. [Google Scholar] [CrossRef]
- Rahman, M.; Zhang, K.; Wang, Y.; Ahmad, B.; Ahmad, A.; Zhang, Z.; Khan, D.; Muhammad, D.; Ali, A. Variations in soil physico-chemical properties, soil stocks, and soil stoichiometry under different soil layers, the major forest region Liupan Mountains of Northwest China. Braz. J. Biol. 2022, 84, e256565. [Google Scholar] [CrossRef] [PubMed]
- Vance, E.D.; Brookes, P.C.; Jeninson, D.S. An Extraction Method for Measuring Microbial Biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.; An, S. Ecological stoichiometry in leaves, roots, litters and soil among di ff erent plant communities in a deserti fi ed region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: A review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lei, P.; Xiang, W.; Yan, W.; Chen, X. Accumulation of soil organic C and N in planted forests fostered by tree species mixture. Biogeosciences 2017, 14, 3937–3945. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Tang, Z.; Ji, C.; Liu, H.; Ma, W.; Mohhamot, A.; Zheng, C. Scaling of nitrogen and phosphorus across plant organs in shrubland biomes. Sci. Rep. 2014, 4, 5448. [Google Scholar] [CrossRef]
- Yang, J.J.; Zhang, X.R.; Ma, L.S.; Chen, Y.N.; Dang, Y.H.; An, S.S. Ecological stoichiometric relationships between components of Robinia pseudoacacia forest in Loess Plateau. Acta Pedol. Sin. 2014, 51, 133–142. [Google Scholar]
- Liu, J.; Gu, Z.; Shao, H.; Zhou, F.; Peng, S. N–P stoichiometry in soil and leaves of Pinus massoniana forest at different stand ages in the subtropical soil erosion area of China. Environ. Earth Sci. 2016, 75, 1091. [Google Scholar] [CrossRef]
- Pang, D.; Wang, G.; Li, G.; Sun, Y.; Liu, Y. Ecological Stoichiometric Characteristics of Two Typical Plantations in the Karst Ecosystem of Southwestern China. Forests 2018, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Xie, G.; Fan, S.; Zhen, L. Variation in vegetation structure and soil properties, and the relation between understory plants and environmental variables under different phyllostachys pubescens forests in Southeastern China. Environ. Manag. 2010, 45, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R.; Caluwe, H.D.; Beltman, B. Is the relation between nutrient supply and biodiversity co-determined by the type of nutrient limitation? Oikos 2003, 101, 489–498. [Google Scholar] [CrossRef]
- Russo, D.; Marziliano, P.A.; Macri, G.; Proto, A.R.; Zimbalatti, G.; Lombardi, F. Does Thinning Intensity Affect Wood Quality? An Analysis of Calabrian Pine in Southern Italy Using a Non-Destructive Acoustic Method. Forests 2019, 10, 303. [Google Scholar] [CrossRef] [Green Version]
- Matzek, V.; Vitousek, P.M. LETTER N:P stoichiometry and protein: RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis. Ecol. Lett. 2009, 12, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Jacobs, D.F.; Zeng, S.; Yang, Z.; Guo, S.; Liu, B. Relationship between above-ground biomass allocation and stand density index in Populus × euramericana stands. Forestry 2012, 85, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Schreeg, L.A.; Santiago, L.S.; Wright, S.J.B.L.T. Stem, root, and older leaf N:P ratios are more responsive R eports R eports. Ecology 2014, 95, 2062–2068. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Fang, J.; Guo, D.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- Reed, S.C.; Townsend, A.R.; Davidson, E.A.; Cleveland, C.C. Stoichiometric patterns in foliar nutrient resorption across multiple scales. New Phytol. 2012, 196, 173–180. [Google Scholar] [CrossRef]
- Cheng, X.; Yu, M.; Wang, G.G. Effects of Thinning on Soil Organic Carbon Fractions and Soil Properties in Cunninghamia lanceolata Stands in Eastern China. Forests 2017, 8, 198. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Zhong, Z.; Sun, Z.; Han, X.; Ren, C.; Yang, G. Soil available phosphorus and moisture drive nutrient resorption patterns in plantations on the Loess Plateau. For. Ecol. Manag. 2020, 461, 117910. [Google Scholar] [CrossRef]
- Han, X.; Gao, G.; Chang, R.; Li, Z.; Ma, Y.; Wang, S.; Wang, C.; Lü, Y.; Fu, B. Changes in soil organic and inorganic carbon stocks in deep pro fi les following cropland abandonment along a precipitation gradient across the Loess Plateau of China. Agric. Ecosyst. Environ. 2018, 258, 1–13. [Google Scholar] [CrossRef]
- Houlton, B.Z.; Wang, Y.-P.; Vitousek, P.M.; Field, C.B. A unifying framework for dinitrogen fixation in the terrestrial biosphere. Nature 2008, 454, 327. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Age (Year) | Stand Density (Trees ha−1) | DBH (cm) | Height (m) | Light Transmittance (%) |
|---|---|---|---|---|
| 9-yr | 3651 ± 393 | 9.1 ± 0.7 | 5.74 ± 0.3 | 35.1 ± 2.8 |
| 18-yr | 1556 ± 239 | 14.9 ± 0.2 | 11.4 ± 0.9 | 33.6 ± 2.5 |
| 28-yr | 1196 ± 51 | 21.6 ± 0.4 | 15.3 ± 0.6 | 49.8 ± 3.5 |
| 48-yr | 774 ± 92 | 25.5 ± 2.9 | 16.8 ± 2.2 | 48.2 ± 5.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.; Hussain, M.; Ali, S.; Akhtar, K.; Muhammad, M.W.; Zamir, A.; Ali, A.; Nizami, S.M.; Ahmad, B.; Harrison, M.T.; et al. Ecological Stoichiometry in Pinus massoniana L. Plantation: Increasing Nutrient Limitation in a 48-Year Chronosequence. Forests 2022, 13, 469. https://doi.org/10.3390/f13030469
Ali A, Hussain M, Ali S, Akhtar K, Muhammad MW, Zamir A, Ali A, Nizami SM, Ahmad B, Harrison MT, et al. Ecological Stoichiometry in Pinus massoniana L. Plantation: Increasing Nutrient Limitation in a 48-Year Chronosequence. Forests. 2022; 13(3):469. https://doi.org/10.3390/f13030469
Chicago/Turabian StyleAli, Ashfaq, Majid Hussain, Saqib Ali, Kashif Akhtar, Mamoona Wali Muhammad, Ahmad Zamir, Arshad Ali, Syed Moazzam Nizami, Bilal Ahmad, Matthew Tom Harrison, and et al. 2022. "Ecological Stoichiometry in Pinus massoniana L. Plantation: Increasing Nutrient Limitation in a 48-Year Chronosequence" Forests 13, no. 3: 469. https://doi.org/10.3390/f13030469