
Response of Poplar Leaf Transcriptome to Changed Management and Environmental Conditions in Pure and Mixed with Black Locust Stands

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
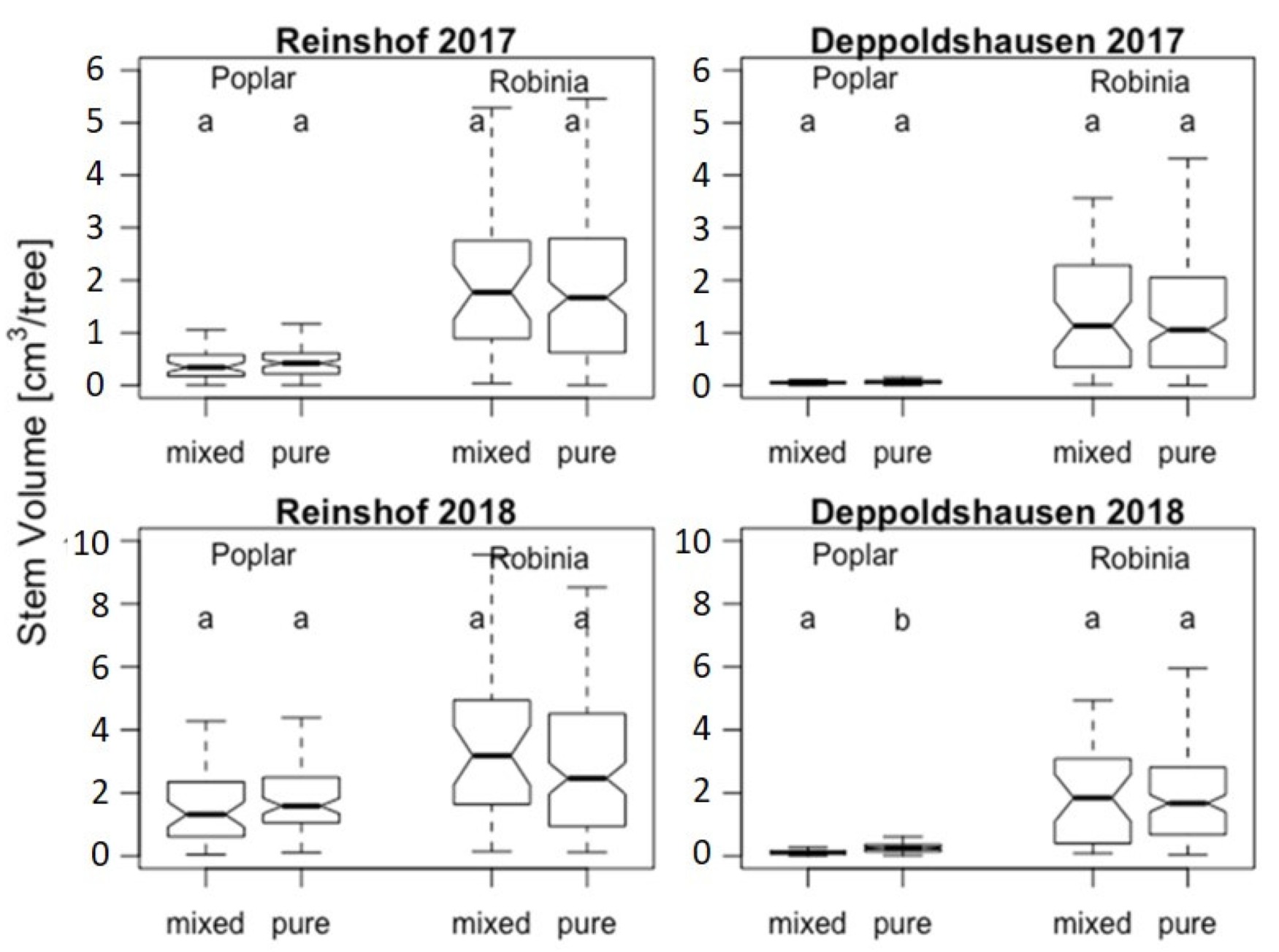
2.2. Stem Volume Measurements
2.3. Sampling for Transcriptome Studies
2.4. RNA Extraction
2.5. RNA Sequencing
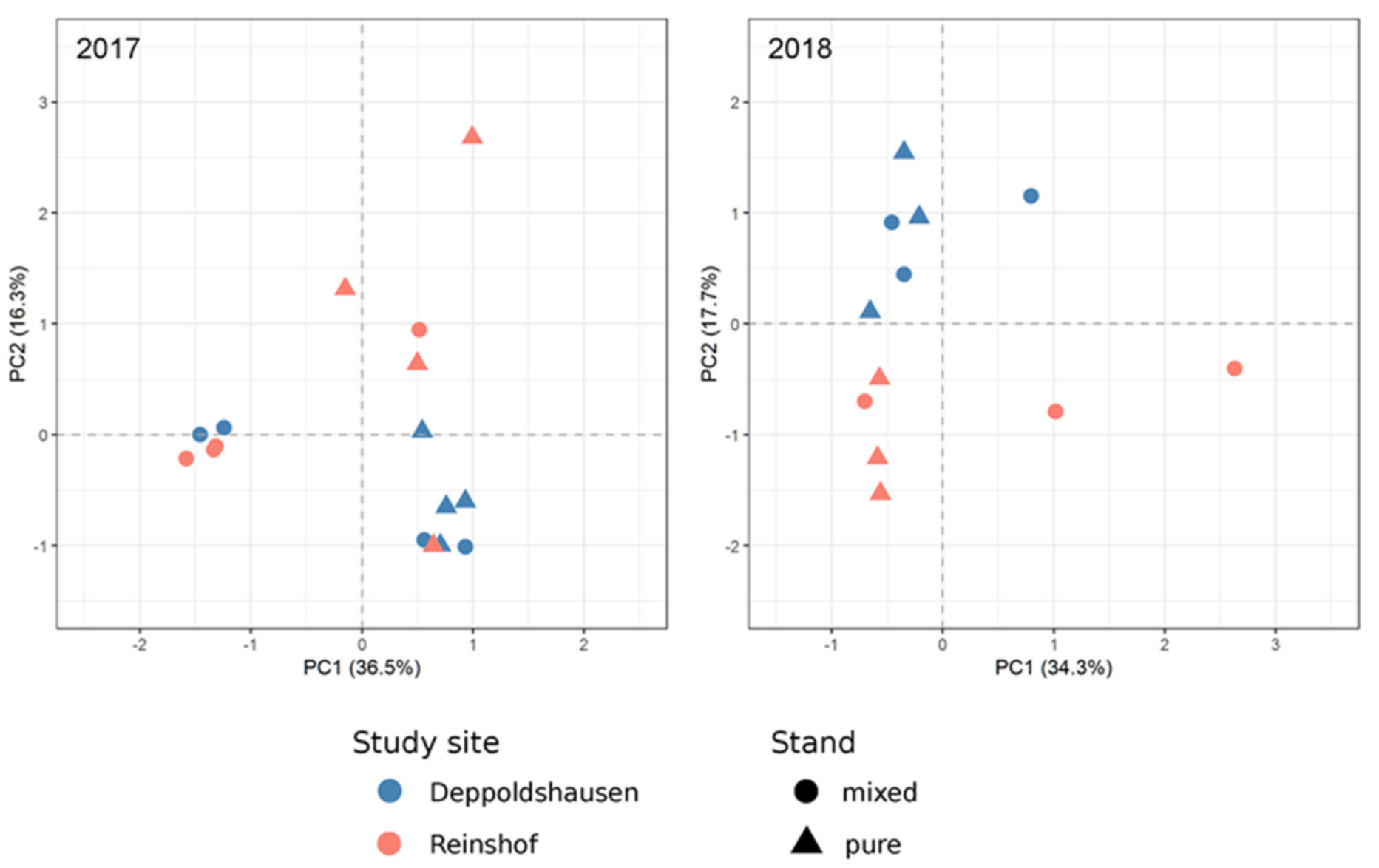
2.6. Principal Component Analysis (PCA)
2.7. Identification of Differently Expressed Genes (DEGs) and Sequence Annotation
2.8. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR)
3. Results
3.1. Weather Data
3.2. Stem Volume Measurements
3.3. RNA Sequencing Output and Sequence Annotation
3.4. Principal Component Analysis (PCA)
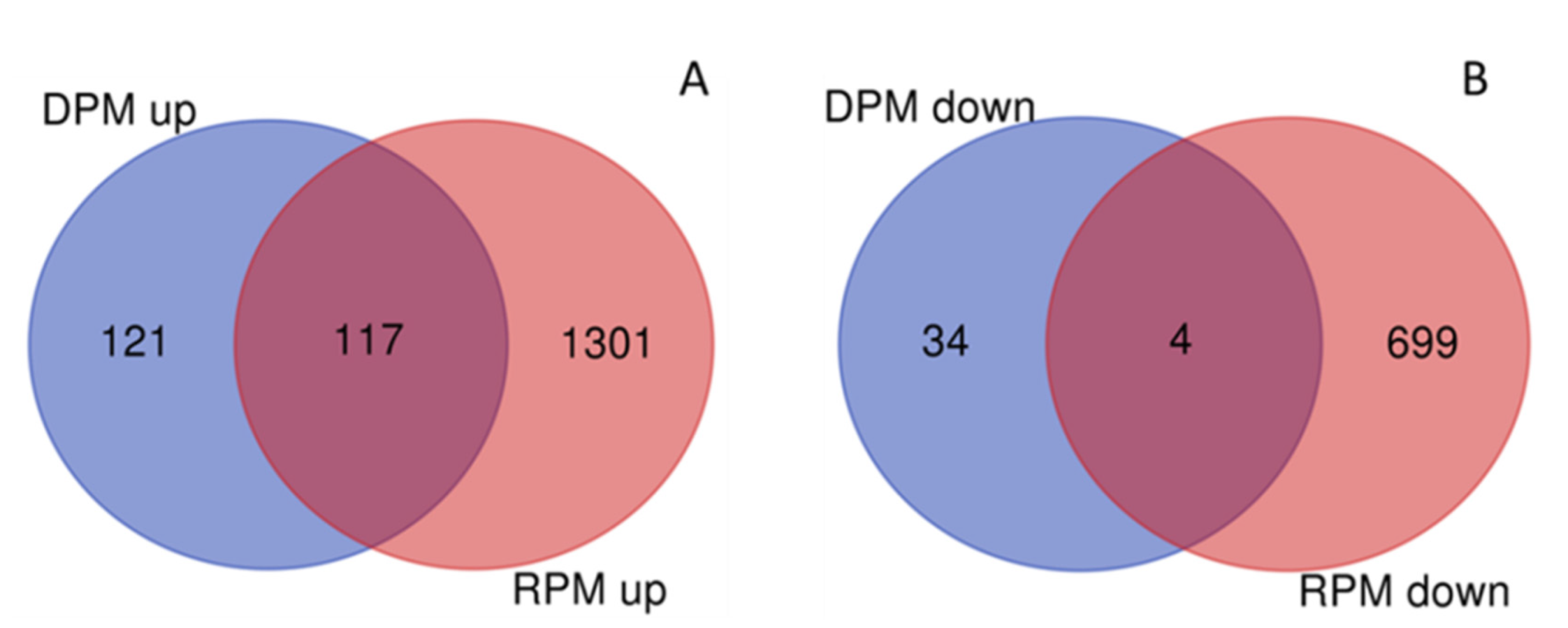
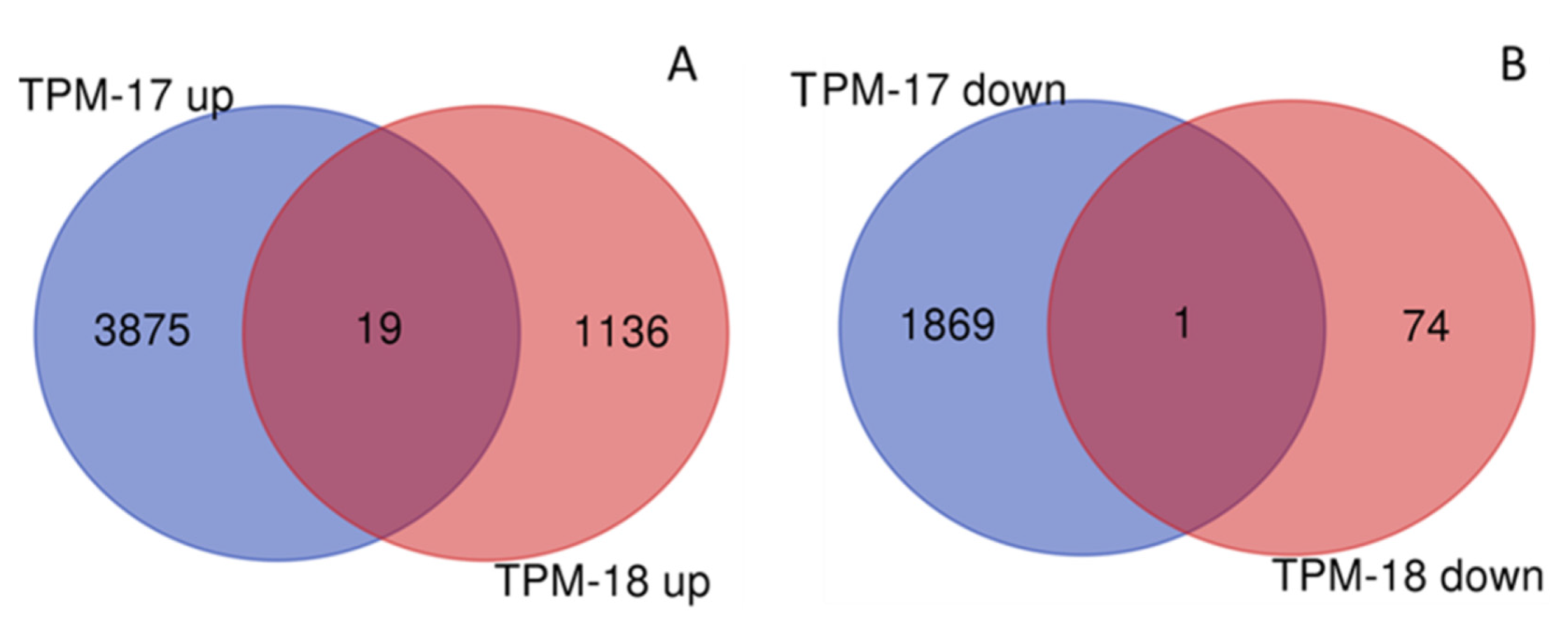
3.5. DEGs between Study Sites and Pure and Mixed Stands
3.6. GO Term Enrichment Analysis
3.6.1. 2017 Samples
3.6.2. 2018 Samples
3.7. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR)
4. Discussion
4.1. Stem Volume Measurements
4.2. PCA and DEG Analyses
4.3. GO Term Enrichment
4.3.1. 2017 Samples
4.3.2. 2018 Samples
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baltodano, J. Monoculture forestry: A critique from an ecological perspective. In Tree Trouble: A Compilation of Testimonies on the Negative Impact of Large-Scale Monoculture Tree Plantations; Prepared for the 6th COP of the FCCC.; Friends of the Earth International: Amsterdam, The Netherlands, 2000; pp. 2–10. [Google Scholar]
- Bowyer, J. Forest plantations Threatening or Saving Natural Forests? Arborvitae (IUCN/WWF For. Conserv. Newsl.) 2006, 31, 8–9. [Google Scholar]
- Morris, J.; Ningnan, Z.; Zengjiang, Y.; Collopy, J.; Daping, X. Water use by fast-growing Eucalyptus urophylla plantations in southern China. Tree Physiol. 2004, 24, 1035–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnus, J.M.; Parrotta, J.; Brockerhoff, E.; Arbez, M.; Jactel, H.; Kremer, A.; Lamb, D.; O’Hara, K.; Walters, B. Planted forests and biodiversity. J. For. 2006, 104, 65–77. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Ferraz, S.F.B. Role of eucalypt and other planted forests in biodiversity conservation and the provision of biodiversity-related ecosystem services. For. Ecol. Manag. 2013, 301, 43–50. [Google Scholar] [CrossRef]
- Liu, C.L.C.; Kuchma, O.; Krutovsky, K.V. Mixed-species versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Glob. Ecol. Conserv. 2018, 15, e00419. [Google Scholar] [CrossRef]
- Ammer, C. Diversity and forest productivity in a changing climate. New Phytol. 2019, 221, 50–66. [Google Scholar] [CrossRef] [Green Version]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: A review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef] [Green Version]
- Sayyad, E.; Hosseini, S.M.; Mokhtari, J.; Mahdavi, R.; Jalali, S.G.; Akbarinia, M.; Tabari, M. Comparison of growth, nutrition and soil properties of pure and mixed stands of Populus deltoides and Alnus subcordata. Silva Fenn. 2006, 40, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Piotto, D. A meta-analysis comparing tree growth in monocultures and mixed plantations. For. Ecol. Manag. 2008, 255, 781–786. [Google Scholar] [CrossRef]
- Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.H.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A. Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann. For. Sci. 2010, 67, 712. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Dieler, J.; Seifert, T.; Rötzer, T. Climate effects on productivity and resource-use efficiency of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) in stands with different spatial mixing patterns. Trees Struct. Funct. 2012, 26, 1343–1360. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef]
- Benomar, L.; DesRochers, A.; Larocque, G.R. Comparing growth and fine root distribution in monocultures and mixed plantations of hybrid poplar and spruce. J. For. Res. 2013, 24, 247–254. [Google Scholar] [CrossRef]
- Grossiord, C.; Gessler, A.; Granier, A.; Pollastrini, M.; Bussotti, F.; Bonal, D. Interspecific competition influences the response of oak transpiration to increasing drought stress in a mixed Mediterranean forest. For. Ecol. Manag. 2014, 318, 54–61. [Google Scholar] [CrossRef]
- Saha, S.; Kuehne, C.; Kohnle, U.; Brang, P.; Ehring, A.; Geisel, J.; Leder, B.; Muth, M.; Petersen, R.; Peter, J.; et al. Growth and quality of young oaks (Quercus robur and Quercus petraea) grown in cluster plantings in Central Europe: A weighted meta-analysis. For. Ecol. Manag. 2012, 283, 106–118. [Google Scholar] [CrossRef]
- Pretzsch, H.; Rais, A. Wood quality in complex forests versus even-aged monocultures: Review and perspectives. Wood Sci. Technol. 2016, 50, 845–880. [Google Scholar] [CrossRef]
- Petráš, R.; Mecko, J.; Bošelâ, M.; Šebeň, V. Wood quality and value production in mixed fir-spruce-beech stands: Long-term research in the Western Carpathians. For. J. 2016, 62, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Russo, D.; Marziliano, P.A.; Macrì, G.; Zimbalatti, G.; Tognetti, R.; Lombardi, F. Tree growth and wood quality in pure vs. mixed-species stands of European beech and Calabrian pine in Mediterranean mountain forests. Forests 2020, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Marron, N.; Epron, D. Are mixed-tree plantations including a nitrogen-fixing species more productive than monocultures? For. Ecol. Manag. 2019, 441, 242–252. [Google Scholar] [CrossRef]
- DeBell, D.S.; Cole, T.G.; Whitesell, C.D. Growth, development, and yield in pure and mixed stands of Eucalyptus and Albizia. For. Sci. 1997, 43, 286–298. [Google Scholar] [CrossRef]
- Rédei, K.; Veperdi, I.; Meilby, H. Stand structure and growth of mixed white poplar (Populus alba L.) and black locust (Robinia pseudoacacia L.) plantations in Hungary. Acta Silv. Lignaria Hung. 2006, 2, 23–32. [Google Scholar]
- Moukoumi, J.; Farrell, R.E.; Van Rees, K.J.C.; Hynes, R.K.; Bélanger, N. Intercropping Caragana arborescens with Salix miyabeana to satisfy nitrogen demand and maximize growth. Bioenergy Res. 2012, 5, 719–732. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, N.; del Río, M.; Forrester, D.I.; Rodríguez-Soalleiro, R.; Pérez-Cruzado, C.; Cañellas, I.; Sixto, H. Mixed short rotation plantations of Populus alba and Robinia pseudoacacia for biomass yield. For. Ecol. Manag. 2018, 410, 48–55. [Google Scholar] [CrossRef]
- Tanaka-Oda, A.; Kenzo, T.; Koretsune, S.; Sasaki, H.; Fukuda, K. Ontogenetic changes in water-use efficiency (δ13C) and leaf traits differ among tree species growing in a semiarid region of the Loess Plateau, China. For. Ecol. Manag. 2010, 259, 953–957. [Google Scholar] [CrossRef]
- Rédei, K.; Keseru, Z.; Rásó, J.; Juhász, L.; Gyori, J.; Antal, B. Growth and yield of mixed black locust (Robinia pseudoacacia L.) and white poplar (Populus alba L.) stands under sandy soil conditions in Hungary: A case study. Silva Balc. 2012, 13, 20–29. [Google Scholar]
- Marron, N.; Priault, P.; Gana, C.; Gérant, D.; Epron, D. Prevalence of interspecific competition in a mixed poplar/black locust plantation under adverse climate conditions. Ann. For. Sci. 2018, 75, 23. [Google Scholar] [CrossRef] [Green Version]
- Rebola-Lichtenberg, J.; Schall, P.; Annighöfer, P.; Ammer, C.; Leinemann, L.; Polle, A.; Euring, D. Mortality of different Populus genotypes in recently established mixed short rotation coppice with Robinia pseudoacacia L. Forests 2019, 10, 410. [Google Scholar] [CrossRef] [Green Version]
- Rebola-Lichtenberg, J.; Streit, J.; Schall, P.; Ammer, C.; Seidel, D. From facilitation to competition: The effect of black locust (Robinia pseudoacacia L.) on the growth performance of four poplar-hybrids (Populus spp.) in mixed short rotation coppice. New For. 2021, 52, 639–656. [Google Scholar] [CrossRef]
- Rebola-Lichtenberg, J.; Schall, P.; Ammer, C. Biomass production in mixed short rotation coppice with poplar-hybrids (Populus spp.) and black locust (Robinia pseudoacacia L.). GCB Bioenergy 2021, 13, 1924–1938. [Google Scholar] [CrossRef]
- Euring, D.; Janz, D.; Polle, A. Wood properties and transcriptional responses of poplar hybrids in mixed cropping with the nitrogen-fixing species Robinia pseudoacacia. Tree Physiol. 2021, 41, 865–881. [Google Scholar] [CrossRef]
- Grünewald, H.; Böhm, C.; Quinkenstein, A.; Grundmann, P.; Eberts, J.; von Wühlisch, G. Robinia pseudoacacia L.: A lesser known tree species for biomass production. Bioenergy Res. 2009, 2, 123–133. [Google Scholar] [CrossRef]
- Mantovani, D.; Veste, M.; Freese, D. Effects of drought frequency on growth performance and transpiration of young black locust (Robinia pseudoacacia L.). Int. J. For. Res. 2014, 2014, 821891. [Google Scholar] [CrossRef] [Green Version]
- Nicolescu, V.N.; Rédei, K.; Mason, W.L.; Vor, T.; Pöetzelsberger, E.; Bastien, J.C.; Brus, R.; Benčať, T.; Đodan, M.; Cvjetkovic, B.; et al. Ecology, growth and management of black locust (Robinia pseudoacacia L.), a non-native species integrated into European forests. J. For. Res. 2020, 31, 1081–1101. [Google Scholar] [CrossRef] [Green Version]
- Vítková, M.; Müllerová, J.; Sádlo, J.; Pergl, J.; Pyšek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, M.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr.&Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef]
- Neale, D.B.; Kremer, A. Forest tree genomics: Growing resources and applications. Nat. Rev. Genet. 2011, 12, 111–122. [Google Scholar] [CrossRef]
- Joshi, C.P.; DiFazio, S.P.; Kole, C. Genetics, Genomics and Breeding of Poplar; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Cohen, D.; Bogeat-Triboulot, M.B.; Tisserant, E.; Balzergue, S.; Martin-Magniette, M.L.; Lelandais, G.; Ningre, N.; Renou, J.P.; Tamby, J.P.; Le Thiec, D.; et al. Comparative transcriptomics of drought responses in Populus: A meta-analysis of genome-wide expression profiling in mature leaves and root apices across two genotypes. BMC Genom. 2010, 11, 630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gugger, P.F.; Peñaloza-Ramírez, J.M.; Wright, J.W.; Sork, V.L. Whole-transcriptome response to water stress in a California endemic oak, Quercus lobata. Tree Physiol. 2017, 37, 632–644. [Google Scholar] [CrossRef]
- Yu, D.; Janz, D.; Zienkiewicz, K.; Herrfurth, C.; Feussner, I.; Chen, S.; Polle, A. Wood formation under severe drought invokes adjustment of the hormonal and transcriptional landscape in Poplar. Int. J. Mol. Sci. 2021, 22, 9899. [Google Scholar] [CrossRef]
- Hess, M.; Wildhagen, H.; Junker, L.V.; Ensminger, I. Transcriptome responses to temperature, water availability and photoperiod are conserved among mature trees of two divergent Douglas-fir provenances from a coastal and an interior habitat. BMC Genom. 2016, 17, 682. [Google Scholar] [CrossRef] [Green Version]
- Janz, D.; Behnke, K.; Schnitzler, J.P.; Kanawati, B.; Schmitt-Kopplin, P.; Polle, A. Pathway analysis of the transcriptome and metabolome of salt sensitive and tolerant poplar species reveals evolutionary adaption of stress tolerance mechanisms. BMC Plant Biol. 2010, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Lane, T.; Best, T.; Zembower, N.; Davitt, J.; Henry, N.; Xu, Y.; Koch, J.; Liang, H.; McGraw, J.; Schuster, S.; et al. The green ash transcriptome and identification of genes responding to abiotic and biotic stresses. BMC Genom. 2016, 17, 702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.; Seifert, S.; Lübbe, T.; Leuschner, C.; Finkeldey, R. De novo transcriptome assembly and analysis of differential gene expression in response to drought in European beech. PLoS ONE 2017, 12, e0184167. [Google Scholar] [CrossRef] [Green Version]
- Philippe, R.N.; Ralph, S.G.; Külheim, C.; Jancsik, S.I.; Bohlmann, J. Poplar defense against insects: Genome analysis, full-length cDNA cloning, and transcriptome and protein analysis of the poplar Kunitz-type protease inhibitor family. New Phytol. 2009, 184, 865–884. [Google Scholar] [CrossRef] [PubMed]
- Euring, D.; Bai, H.; Janz, D.; Polle, A. Nitrogen-driven stem elongation in poplar is linked with wood modification and gene clusters for stress, photosynthesis and cell wall formation. BMC Plant Biol. 2014, 14, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Zhou, J.; Li, H.; Shi, W.; Polle, A.; Lu, M.; Sun, X.; Luo, Z.-B. Bin Global poplar root and leaf transcriptomes reveal links between growth and stress responses under nitrogen starvation and excess. Tree Physiol. 2015, 35, 1283–1302. [Google Scholar] [CrossRef] [Green Version]
- Behnke, K.; Grote, R.; Brüggemann, N.; Zimmer, I.; Zhou, G.; Elobeid, M.; Janz, D.; Polle, A.; Schnitzler, J.P. Isoprene emission-free poplars—A chance to reduce the impact from poplar plantations on the atmosphere. New Phytol. 2012, 194, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Izawa, T. Deciphering and prediction of plant dynamics under field conditions. Curr. Opin. Plant Biol. 2015, 24, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Brosché, M.; Vinocur, B.; Alatalo, E.R.; Lamminmäki, A.; Teichmann, T.; Ottow, E.A.; Djilianov, D.; Afif, D.; Bogeat-Triboulot, M.B.; Altman, A.; et al. Gene expression and metabolite profiling of Populus euphratica growing in the Negev desert. Genome Biol. 2005, 6, R101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holliday, J.A.; Ralph, S.G.; White, R.; Bohlmann, J.; Aitken, S.N. Global monitoring of autumn gene expression within and among phenotypically divergent populations of Sitka spruce (Picea sitchensis). New Phytol. 2008, 178, 103–122. [Google Scholar] [CrossRef]
- Ning, K.; Ding, C.; Huang, Q.; Zhang, W.; Yang, C.; Liang, D.; Fan, R.; Su, X. Transcriptome profiling revealed diverse gene expression patterns in poplar (Populus × euramericana) under different planting densities. PLoS ONE 2019, 14, e0217066. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qian, C.; Yin, X.; Fan, X.; Zhao, X.; Gu, M.; Wang, T.; Ma, X.F. A whole-transcriptome approach to evaluate reference genes for quantitative diurnal gene expression studies under natural field conditions in Tamarix ramosissima leaves. Electron. J. Biotechnol. 2018, 35, 48–56. [Google Scholar] [CrossRef]
- Allwright, M.R.; Payne, A.; Emiliani, G.; Milner, S.; Viger, M.; Rouse, F.; Keurentjes, J.J.B.B.; Bérard, A.; Wildhagen, H.; Faivre-Rampant, P.; et al. Biomass traits and candidate genes for bioenergy revealed through association genetics in coppiced European Populus nigra (L.). Biotechnol. Biofuels 2016, 9, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaling, M.; Schmidt, A.; Moritz, F.; Rosenkranz, M.; Witting, M.; Kasper, K.; Janz, D.; Schmitt-Kopplin, P.; Schnitzler, J.-P.P.; Polle, A. Mycorrhiza-triggered transcriptomic and metabolomic networks impinge on herbivore fitness. Plant Physiol. 2018, 176, 2639–2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BodSchätzG. Gesetz zur Schätzung des Landwirtschaftlichen Kulturbodens [Law on the Estimation of Soil Fertility]. 2007. Available online: http://www.gesetze-im-internet.de/bodsch_tzg_2008/BodSch%C3%A4tzG.pdf) (accessed on 18 April 2020).
- Röhle, H.; Hartmann, K.U.; Gerold, D.; Steinke, C.; Schröder, J. Biomass functions for short rotation forestry. Allg. Forst- und Jagdzeitung 2006, 177, 178–187. [Google Scholar]
- Ammer, C.; Brang, P.; Knoke, T.; Wagner, S. Methoden zur waldbaulichen Untersuchung von Jungwüchsen. Forstarchiv 2004, 75, 83–110. [Google Scholar]
- Annighöfer, P.; Ameztegui, A.; Ammer, C.; Balandier, P.; Bartsch, N.; Bolte, A.; Coll, L.; Collet, C.; Ewald, J.; Frischbier, N.; et al. Species-specific and generic biomass equations for seedlings and saplings of European tree species. Eur. J. For. Res. 2016, 135, 313–329. [Google Scholar] [CrossRef] [Green Version]
- Urbanek, S.; Bibiko, H.-J.; Stefano, M.L. R: A Language and Environment for Statistical Computing. The R Foundation for Statistical Computing. Available online: http://www.r-project.org/ (accessed on 22 November 2019).
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. FASTQ: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Vu, V.Q. Ggbiplot: A Ggplot2 Based Biplot. R Package, Version 0.55. Available online: http://github.com/ (accessed on 22 November 2019).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunner, A.M.; Yakovlev, I.A.; Strauss, S.H. Validating internal controls for quantitative plant gene expression studies. BMC Plant Biol. 2004, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kibbe, W.A. OligoCalc: An online oligonucleotide properties calculator. Nucleic Acids Res. 2007, 35, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Barbaroux, C.; Le Thiec, D.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides x Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Kuchma, O.; Janz, D.; Leinemann, L.; Polle, A.; Krutovsky, K.V.; Gailing, O. Hybrid and environmental effects on gene expression in poplar clones in pure and mixed with black locust stands. Forests 2020, 11, 1075. [Google Scholar] [CrossRef]
- Boyden, S.; Binkley, D.; Senock, R. Competition and facilitation between Eucalyptus and nitrogen-fixing Falcataria in relation to soil fertility. Ecology 2005, 86, 992–1001. [Google Scholar] [CrossRef]
- Bouillet, J.P.; Laclau, J.P.; de M. Gonçalves, J.L.; Voigtlaender, M.; Gava, J.L.; Leite, F.P.; Hakamada, R.; Mareschal, L.; Mabiala, A.; Tardy, F.; et al. Eucalyptus and Acacia tree growth over entire rotation in single- and mixed-species plantations across five sites in Brazil and Congo. For. Ecol. Manag. 2013, 301, 89–101. [Google Scholar] [CrossRef]
- Vítková, M.; Tonika, J.; Müllerová, J. Black locust–successful invader of a wide range of soil conditions. Sci. Total Environ. 2015, 505, 315–328. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Zhang, Z.; Lv, Y.; Yang, N.; Zhang, G.; Wu, M.; Lv, S.; Pan, L.; Joosten, M.H.A.J.; et al. Transcriptional regulation of receptor-like protein genes by environmental stresses and hormones and their overexpression activities in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 3339–3351. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.P.; Leach, J.E. Abiotic and biotic stresses induce a core transcriptome response in rice. Sci. Rep. 2019, 9, 6273. [Google Scholar] [CrossRef]
- Ballaré, C.L.; Pierik, R. The shade-avoidance syndrome: Multiple signals and ecological consequences. Plant Cell Environ. 2017, 40, 2530–2543. [Google Scholar] [CrossRef]
- Casal, J.J. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 2013, 64, 403–427. [Google Scholar] [CrossRef] [PubMed]
- Roig-Villanova, I.; Martínez-García, J.F. Plant responses to vegetation proximity: A whole life avoiding shade. Front. Plant Sci. 2016, 7, 236. [Google Scholar] [CrossRef] [Green Version]
- Courbier, S.; Pierik, R. Canopy light quality modulates stress responses in plants. iScience 2019, 22, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Andersen, E.J.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M.P. Disease resistance mechanisms in plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, S.; Amasino, R.M. Making sense of senescence: Molecular genetic regulation and manipulation of leaf senescence. Plant Physiol. 1997, 113, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munné-Bosch, S.; Alegre, L. Die and let live: Leaf senescence contributes to plant survival under drought stress. Funct. Plant Biol. 2004, 31, 203–216. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Abbreviation | Species or Hybrid |
|---|---|---|
| Poplar clones | ||
| AF2 | P1 | Populus deltoides × P. nigra |
| Fritzi Pauley | P2 | P. trichocarpa |
| Hybride 275 | P3 | P. maximowiczii × P. trichocarpa |
| I214 | P4 | P. deltoides × P. nigra |
| Matrix 11 | P5 | P. maximowiczii × P. trichocarpa |
| Matrix 49 | P6 | P. maximowiczii × P. trichocarpa |
| Max 1 | P7 | P. nigra × P. maximowiczii |
| Muhle Larsen | P8 | P. trichocarpa |
| Black locust provenances | ||
| HGK 81901, Germany | R1 | Robinia pseudoacacia |
| HGK 81902, Germany | R2 | R. pseudoacacia |
| Nagybudmry, Hungary | R3 | R. pseudoacacia |
| Year | Species | Reinshof | Deppoldshausen | ||||
|---|---|---|---|---|---|---|---|
| Mixed | Pure | p | Mixed | Pure | p | ||
| 2017 | Populus “Max 1” | 4924 | 5205 | 0.79 | 691 | 855 | 0.06 |
| Robinia p. | 23,009 | 20,478 | 0.44 | 13,127 | 14,531 | 0.49 | |
| 2018 | Populus “Max 1” | 16,722 | 19,641 | 0.28 | 1292 | 2605 | 9.5 × 10−8 |
| Robinia p. | 35,564 | 31,359 | 0.77 | 18,337 | 19,969 | 0.59 | |
| Year | DEGs | TPM a | TSS | RPM a | DPM a |
|---|---|---|---|---|---|
| 2017 | Upregulated | 3894 | 0 | 1418 | 238 |
| Downregulated | 1870 | 0 | 703 | 38 | |
| Total | 5764 | 0 | 2121 | 276 | |
| 2018 | Upregulated | 1155 | 1139 b | 1654 | 0 |
| Downregulated | 75 | 1357 c | 111 | 1 | |
| Total | 1230 | 2496 | 1765 | 1 |
| GO Term | TPM | DPM | RPM | TSS-D | TSS-R | |||
|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2018 | 2018 | |
| Downregulated | ||||||||
| photosynthesis (GO: 0015979) | 176 | - | 13 | - | - | - | - | - |
| generation of precursor metabolites and energy (GO: 0006091) | 205 | - | 13 | - | - | - | - | - |
| pigment biosynthetic process (GO: 0046148) | 77 | - | - | - | - | - | - | - |
| Upregulated | ||||||||
| response to stress (GO: 0006950) | 927 | - | 72 | - | 368 | - | 270 | 317 |
| response to wounding (GO: 0009611) | 103 | - | 14 | - | 55 | 50 | - | - |
| response to starvation (GO: 0042594) | 99 | - | 15 | - | - | - | - | - |
| response to water deprivation (GO: 0009414) | 116 | - | - | - | 59 | - | 54 | - |
| response to biotic stimulus (GO: 0009607) | 459 | - | 39 | - | 170 | - | - | 185 |
| response to fungus (GO: 0009620) | 140 | - | - | - | 64 | - | - | - |
| response to bacterium (GO: 0009617) | 146 | - | - | - | 65 | - | - | 85 |
| hormone metabolic process (GO:0042445) | 210 | - | 9 | - | 86 | - | - | - |
| leaf senescence (GO:0010150) | 32 | - | - | - | 15 | - | - | - |
| shade avoidance (GO: 0009641) | 10 | - | - | - | 6 | - | - | 8 |
| photosynthesis (GO: 0015979) | - | - | - | - | - | - | 43 | - |
| NADPH regeneration (GO: 0006740) | - | - | - | - | - | - | 27 | - |
| response to heat (GO: 0009408) | - | - | - | - | - | - | 92 | - |
| pigment biosynthetic process (GO: 0046148) | - | - | - | - | - | - | 36 | - |
| pigment accumulation (GO: 0043476) | - | - | - | - | - | 31 | - | 15 |
| hormone metabolic process (GO: 0042445) | - | - | - | - | - | - | - | 103 |
| response to auxin (GO: 0009733) | - | 37 | - | - | - | 48 | - | 43 |
| hormone transport (GO: 0009914) | - | 26 | - | - | - | 36 | - | - |
| shade avoidance (GO: 0009641) | - | - | - | - | - | - | - | 8 |
| nitrate transport (GO: 0015706) | - | 21 | - | - | - | 26 | - | - |
| plant organ development (GO: 0099402) | - | 105 | - | - | - | 149 | - | 134 |
| cell wall organization or biogenesis (GO: 0071554) | - | 93 | - | - | - | 164 | - | - |
| cell growth (GO: 0016049) | - | 59 | - | - | - | 92 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuchma, O.; Rebola-Lichtenberg, J.; Janz, D.; Krutovsky, K.V.; Ammer, C.; Polle, A.; Gailing, O. Response of Poplar Leaf Transcriptome to Changed Management and Environmental Conditions in Pure and Mixed with Black Locust Stands. Forests 2022, 13, 147. https://doi.org/10.3390/f13020147
Kuchma O, Rebola-Lichtenberg J, Janz D, Krutovsky KV, Ammer C, Polle A, Gailing O. Response of Poplar Leaf Transcriptome to Changed Management and Environmental Conditions in Pure and Mixed with Black Locust Stands. Forests. 2022; 13(2):147. https://doi.org/10.3390/f13020147
Chicago/Turabian StyleKuchma, Oleksandra, Jessica Rebola-Lichtenberg, Dennis Janz, Konstantin V. Krutovsky, Christian Ammer, Andrea Polle, and Oliver Gailing. 2022. "Response of Poplar Leaf Transcriptome to Changed Management and Environmental Conditions in Pure and Mixed with Black Locust Stands" Forests 13, no. 2: 147. https://doi.org/10.3390/f13020147