
Relationship between Photosynthetic Characteristics, Anatomical Structure, and Physiological Indexes of Two Halophytes in Different Habitats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Sites
2.2. Experimental Design
2.3. Sample Collection
2.4. Measurement Index
2.4.1. Determination of Soil Indexes
2.4.2. Determination of Photosynthetic Parameters
2.4.3. Determination of Leaf Morphology and Anatomical Structure
2.4.4. Determination of Leaf Water Potential
2.4.5. Determination of Relative Chlorophyll Content in Leaves
2.5. Statistical Analyses
3. Results
3.1. The Variation Characteristics of Soil Environmental Factors in Different Habitats
3.2. The SPAD and Water Potential of Leaves in Different Habitats
3.3. Leaf Morphology and Anatomical Characteristics in Different Habitats
3.4. Changes of Photosynthetic Parameters of Two Species in Different Habitats
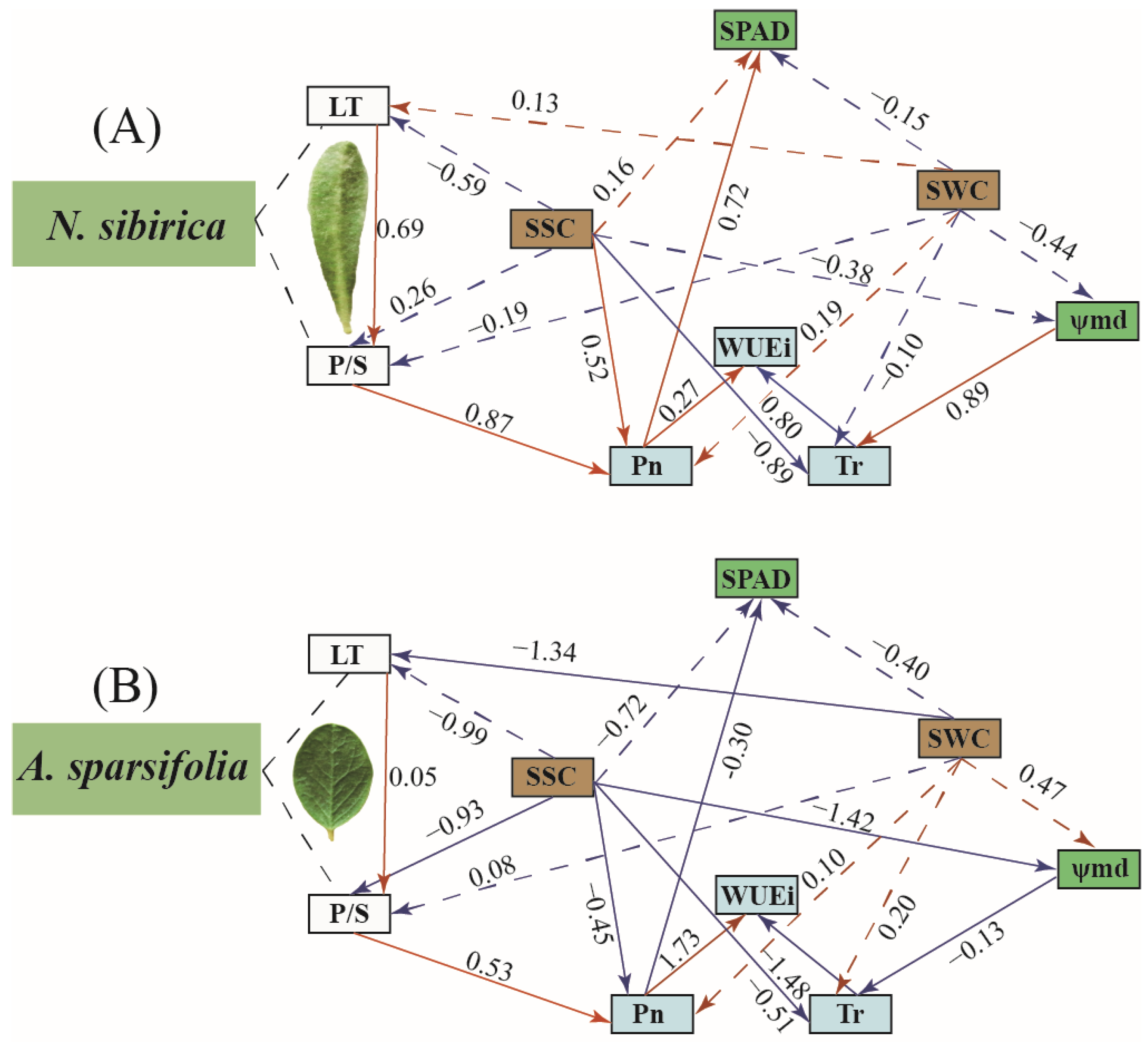
3.5. Driving Factors of WUEi in the Leaves of Two Plants in Different Habitats
4. Discussion
4.1. Species-Specific Adaptations of Leaf Morphology and Anatomy
4.2. Effects of Soil Water and Salt Environment on Plant Photosynthesis
4.3. Correlation between Photosynthetic Characteristics and Other Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, R.; Tian, Q.; Zhang, Y.; Wu, Y.; Li, Z.; Tang, Z.; Zhou, A. Response of Leaf Functional Traits of Landscape Plants to Urban Green Space Environment in Lanzhou, China. Forests 2022, 13, 682. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive La Différence: Plant Functional Diversity Matters to Ecosystem Processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Fortunel, C.; Fine, P.V.A.; Baraloto, C. Leaf, Stem and Root Tissue Strategies across 758 Neotropical Tree Species. Funct. Ecol. 2012, 26, 1153–1161. [Google Scholar] [CrossRef]
- Liu, Y.; Li, P.; Xu, G.C.; Xiao, L.; Ren, Z.P.; Li, Z.B. Growth, Morphological, and Physiological Responses to Drought Stress in Bothriochloa ischaemum. Front. Plant Sci. 2017, 8, 230. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, E.H.; Springer, C.J.; Helliker, B.R.; Poethig, R.S. MicroRNA156-mediated Changes in Leaf Composition Lead to Altered Photosynthetic Traits during Vegetative Phase Change. New Phytol. 2021, 231, 1008–1022. [Google Scholar] [CrossRef]
- Nagano, A.J.; Kawagoe, T.; Sugisaka, J.; Honjo, M.N.; Iwayama, K.; Kudoh, H. Annual Transcriptome Dynamics in Natural Environments Reveals Plant Seasonal Adaptation. Nat. Plants 2019, 5, 74–83. [Google Scholar] [CrossRef]
- Guerfel, M.; Baccouri, O.; Boujnah, D.; Chaïbi, W.; Zarrouk, M. Impacts of Water Stress on Gas Exchange, Water Relations, Chlorophyll Content and Leaf Structure in the Two Main Tunisian Olive (Olea europaea L.) Cultivars. Sci. Hortic. 2009, 119, 257–263. [Google Scholar] [CrossRef]
- He, W.; Yan, K.; Zhang, Y.; Bian, L.; Mei, H.; Han, G. Contrasting Photosynthesis, Photoinhibition and Oxidative Damage in Honeysuckle (Lonicera japonica Thunb.) under Iso-Osmotic Salt and Drought Stresses. Environ. Exp. Bot. 2021, 182, 104313. [Google Scholar] [CrossRef]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Walcroft, A.; Le Roux, X.; Diaz-Espejo, A.; Dones, N.; Sinoquet, H. Effects of Crown Development on Leaf Irradiance, Leaf Morphology and Photosynthetic Capacity in a Peach Tree. Tree Physiol. 2002, 22, 929–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meir, P.; Levy, P.E.; Grace, J.; Jarvis, P.G. Photosynthetic Parameters from Two Contrasting Woody Vegetation Types in West Africa. Plant Ecol. 2007, 192, 277–287. [Google Scholar] [CrossRef]
- Tholen, D.; Boom, C.; Zhu, X.-G. Opinion: Prospects for Improving Photosynthesis by Altering Leaf Anatomy. Plant Sci. 2012, 197, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The Worldwide Leaf Economics Spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Yuan, Y.; Fu, D.P.; Lv, G.H. Inter-specific relations of the dominant plants of the wetland vegetation in the Erbin Lake wetland in Xinjiang Uygur Autonomous Region. Wetl. Sci. 2008, 6, 486–491. [Google Scholar]
- Xi, J.B.; Zhang, F.S.; Mao, D.R.; Tian, C.Y.; Dong, Z.C.; Wang, K.F. Species Diversity and Distribution of Halophytic Vegetation in Xinjiang. Sci. Silvae Sin. 2006, 10, 6–12. [Google Scholar]
- Tang, X.; Zhang, H.; Shabala, S.; Li, H.; Yang, X.; Zhang, H. Tissue Tolerance Mechanisms Conferring Salinity Tolerance in a Halophytic Perennial Species Nitraria sibirica Pall. Tree Physiol. 2021, 41, 1264–1277. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Y.; Zhu, J.-K. Thriving under Stress: How Plants Balance Growth and the Stress Response. Dev. Cell 2020, 55, 529–543. [Google Scholar] [CrossRef]
- Traditional Uses, Chemistry, Pharmacology, Toxicology and Quality Control of Alhagi sparsifolia Shap: A Review. Available online: https://pubmed.ncbi.nlm.nih.gov/34721046/ (accessed on 18 November 2022).
- He, X.; Lv, G.; Qin, L.; Chang, S.; Yang, M.; Yang, J.; Yang, X. Effects of Simulated Nitrogen Deposition on Soil Respiration in a Populus euphratica Community in the Ebinur Lake Area, a Desert Ecosystem of Northwestern China. PLoS ONE 2015, 10, e0137827. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Lv, G.; Guo, Z.; Chen, Y.; Cao, J. Influence of Aridity and Salinity on Plant Nutrients Scales up from Species to Community Level in a Desert Ecosystem. Sci. Rep. 2017, 7, 6811. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-D.; Zhang, X.-N.; Lv, G.-H.; Ali, A. Linking Populus euphratica Hydraulic Redistribution to Diversity Assembly in the Arid Desert Zone of Xinjiang, China. PLoS ONE 2014, 9, e109071. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.N.; Lv, G.H.; Yang, X.D.; Qin, L.; He, X.M.; Liu, H.Q. Responses of desert plant diversity, community and interspecific association to soil salinity gradient. Acta Ecol. Sin. 2013, 33, 5714–5722. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and AgrIcultural Chemistry Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Guo, Z.C.; Zeng, F.J.; Liu, P.; Li, C.J.; Zhang, B. Photosynthesis and water metabolism of Alhagi sparsifolia Shap. in different geographical populations. Arid. Zone Res. 2016, 33, 371–378. [Google Scholar]
- Olaranont, Y.; Stewart, A.B.; Traiperm, P. Effects of Crude Oil on Plant Growth and Leaf Anatomical Structures in a Common Coastal Plant. Int. J. Phytoremediat. 2021, 23, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.F.; Li, C.H.; Liu, G.H.; Zhou, S.Q.; Su, Y.L.T. Analysis of drought resistance by leaf anatomical structure of six species of sandy willows. J. Desert Res. 2009, 29, 480–484. [Google Scholar]
- Landi, S.; Capasso, G.; Esposito, S. Different G6PDH Isoforms Show Specific Roles in Acclimation to Cold Stress at Various Growth Stages of Barley (Hordeum vulgare) and Arabidopsis thaliana. Plant Physiol. Biochem. 2021, 169, 190–202. [Google Scholar] [CrossRef]
- Liang, W.B.; Zhao, L.J.; Li, J.X.; Xiao, L. Leaves comparative anatomy of Styrax in Hunan. Plant Res. 2014, 34, 148–158. [Google Scholar]
- Shiyab, S.M.; Shatnawi, M.A.; Shibli, R.A.; Al Smeirat, N.G.; Ayad, J.; Akash, M.W. Growth, Nutrient Acquisition, And Physiological Responses Of Hydroponic Grown Tomato To Sodium Chloride Salt Induced Stress. J. Plant Nutr. 2013, 36, 665–676. [Google Scholar] [CrossRef]
- Ni, Q.; Ma, Y.J.; Yang, W.P.; Yang, Y.Y. Effect of NaCl treatment on the anatomy of Lycium ruthenicum leaves. Pratacult. Sci. 2019, 36, 1803–1810. [Google Scholar]
- Ramachandra, R.A.; Viswanatha, C.K.; Munusamy, V. Drought-Induced Responses of Photosynthesis and Antioxidant Metabolism in Higher Plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar]
- Ramin, A.A. Effects of Salinity and Temperature on Germination Seedling Establishment of Sweet Basil (Ocimum basilicum L.). J. Herbs Spices Med. Plants 2006, 11, 81–90. [Google Scholar] [CrossRef]
- Deng, X.; Li, X.M.; Zhang, X.M.; Ye, W.H. A study of the gas exchange characteristics of four desert plants. Chin. J. Plant Ecol. 2002, 26, 605–612. [Google Scholar]
- Hernandez-Santana, V.; Fernández, J.; Rodriguez-Dominguez, C.; Romero, R.; Diaz-Espejo, A. The Dynamics of Radial Sap Flux Density Reflects Changes in Stomatal Conductance in Response to Soil and Air Water Deficit. Agric. For. Meteorol. 2016, 218–219, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Gao, G.L.; Feng, Q.; Zhang, X.Y.; Si, J.H.; Yu, T.F. An overview of stomatal and non-stomatal limitations to photosynthesis of plants. Arid. Zone Res. 2018, 35, 929–937. [Google Scholar] [CrossRef]
- Medlyn, B.E.; De Kauwe, M.G.; Lin, Y.-S.; Knauer, J.; Duursma, R.A.; Williams, C.A.; Arneth, A.; Clement, R.; Isaac, P.; Limousin, J.-M.; et al. How Do Leaf and Ecosystem Measures of Water-Use Efficiency Compare? New Phytol. 2017, 216, 758–770. [Google Scholar] [CrossRef] [Green Version]
- Jian, S.; Wan, S.; Piao, S.; Hui, D.; Hovenden, M.J.; Ciais, P.; Liu, Y.; Liu, Y.; Zhong, M.; Zheng, M. Elevated CO2 Does Not Stimulate Carbon Sink in a Semi-arid Grassland. Ecol. Lett. 2019, 22, 458–468. [Google Scholar] [CrossRef]
- Battipaglia, G.; Saurer, M.; Cherubini, P.; Calfapietra, C.; Mccarthy, H.R.; Norby, R.J.; Cotrufo, M.F. Elevated CO2 Increases Tree-level Intrinsic Water Use Efficiency: Insights from Carbon and Oxygen Isotope Analyses in Tree Rings across Three Forest FACE Sites. New Phytol. 2013, 197, 544–554. [Google Scholar] [CrossRef] [Green Version]
- Duan, B.B.; Zhao, C.Z.; Xu, T.; Zheng, H.L.; Feng, W.; Han, L. Correlation analysis between vein density and stomatal traits of Robinia pseudoacacia in different aspects of Beishan Mountain in Lanzhou. Chin. J. Plant Ecol. 2016, 40, 1289–1297. [Google Scholar]
- Chartzoulakis, K.; Patakas, A.; Kofidis, G.; Bosabalidis, A.; Nastou, A. Water Stress Affects Leaf Anatomy, Gas Exchange, Water Relations and Growth of Two Avocado Cultivars. Sci. Hortic. 2002, 95, 39–50. [Google Scholar] [CrossRef]
- Fei, X.; Guo, W.; Xu, W.; Wei, Y.; Wang, R. Leaf Morphology Correlates with Water and Light Availability: What Consequences for Simple and Compound Leaves? Prog. Nat. Sci. 2009, 19, 1789–1798. [Google Scholar] [CrossRef]
- Nobel, L. Salinity Effects on Leaf Anatomy. Plant Physiol. 1979, 63, 700–703. [Google Scholar] [CrossRef]
- Jarvis, P.G.; Mcnaughton, K.G. Stomatal Control of Transpiration: Scaling Up from Leaf to Region. Adv. Ecol. Res. 1986, 15, 1–49. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Reid, D.M.; Yeung, E.C. Morphological and Physiological Responses of Canola (Brassica napus) Siliquas and Seeds to UVB and CO2 under Controlled Environment Conditions. Environ. Exp. Bot. 2007, 60, 428–437. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Longitude | Latitude | Altitude (m) |
|---|---|---|---|
| HWHS | 83°33′45″ | 44°37′11″ | 260.94 |
| MWMS | 83°33′57″ | 44°38′35″ | 245.45 |
| LWLS | 83°34′00″ | 44°39′37″ | 242.80 |
| Plot | SWC (%) | SSC (g/kg) | pH |
|---|---|---|---|
| HWHS | 11.80 ± 3.44 A | 11.63 ± 1.82 A | 8.26 ± 0.26 A |
| MWMS | 3.04 ± 1.27 B | 4.46 ± 0.22 B | 7.94 ± 0.06 B |
| LWLS | 1.42 ± 0.65 C | 3.00 ± 0.13 C | 7.87 ± 0.06 B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.; Fan, Y.; Li, Z.; Lv, G. Relationship between Photosynthetic Characteristics, Anatomical Structure, and Physiological Indexes of Two Halophytes in Different Habitats. Forests 2022, 13, 2189. https://doi.org/10.3390/f13122189
Chang Y, Fan Y, Li Z, Lv G. Relationship between Photosynthetic Characteristics, Anatomical Structure, and Physiological Indexes of Two Halophytes in Different Habitats. Forests. 2022; 13(12):2189. https://doi.org/10.3390/f13122189
Chicago/Turabian StyleChang, Yaling, Yuxian Fan, Zhoukang Li, and Guanghui Lv. 2022. "Relationship between Photosynthetic Characteristics, Anatomical Structure, and Physiological Indexes of Two Halophytes in Different Habitats" Forests 13, no. 12: 2189. https://doi.org/10.3390/f13122189