Evaluation of Undergrowth under the Canopy of Deciduous Forests on Very Fertile Soils in the Lithuanian Hemiboreal Forest


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Data Collection
2.3. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Lehman, C.L.; Thomson, K.T. Plant diversity and ecosystem productivity: Theoretical considerations. Proc. Natl. Acad. Sci. USA 1997, 94, 1857–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengtsson, J.; Nilsson, S.G.; Franc, A.; Menozzi, P. Biodiversity, disturbances, ecosystem function and management of European forests. For. Ecol. Manag. 2000, 132, 39–50. [Google Scholar] [CrossRef]
- Härdtle, W.; von Oheimb, G.; Westphal, C.W. The effects of light and soil conditions on the species richness of the ground vegetation of deciduous forests in northern Germany (Schleswig-Holstein). For. Ecol. Manag. 2003, 182, 327–338. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y. Understory vegetation dynamics of North American boreal forests. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Gil, W. Krzewy w Gospodarce Leśnej [Shrubs in Forest Management]; PWRIL Państwowe Wydawnictwo Rolnicze i Leśne: Warsawa, Poland, 2014; 214p. (In Polish) [Google Scholar]
- Das Gupta, S.; Pinno, B.D. Drivers of understory species richness in reconstructed boreal ecosystems: A structural equation modeling analysis. Sci. Rep. 2020, 10, 11555. [Google Scholar] [CrossRef]
- Maestre, F.T.; Bowker, M.A.; Puche, M.D.; Hinojosa, M.B.; Martínez, I.; García-Palacios, P.; Castillo, A.P.; Soliveres, S.; Luzuriaga, A.L.; Sánchez, A.M.; et al. Shrub encroachment can reverse desertification in semi-arid Mediterranean grasslands. Ecol. Lett. 2009, 12, 930–941. [Google Scholar] [CrossRef]
- Dhar, A.; Comeau, P.G.; Vassov, R. Effects of cover soil stockpiling on plant community development following reclamation of oil sands sites in Alberta. Restor. Ecol. 2019, 27, 352–360. [Google Scholar] [CrossRef]
- Alberdi, I.; Condés, S.; Mcroberts, R.E.; Winter, S. Mean species cover: A harmonized indicator of shrub cover for forest inventories. Eur. J. For. Res. 2018, 137, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Van Auken, O.W. Causes and Consequences of Woody Plant Encroachment into Western North American Grasslands. J. Environ. Manag. 2009, 90, 2931–2942. [Google Scholar] [CrossRef]
- Kitzberger, T.; Steinaker, D.F.; Veblen, T.T. Effects of climatic variability on facilitation of tree establishment in Northern Patagonia. Ecology 2000, 80, 1914–1924. [Google Scholar] [CrossRef]
- Jensen, A.M.; Götmark, F.; Löf, M. Shrubs protect oak seedlings against ungulate browsing in temperate broadleaved forests of conservation interest: A field experiment. For. Ecol. Manag. 2012, 266, 187–193. [Google Scholar] [CrossRef]
- Nilsson, M.C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Tomppo, E.; Gschwanter, T.; Lawrence, M.; McRoberts, R.E. National Forest Inventories: Pathways for Harmonised Reporting; Springer: Dordrecht, The Netherlands, 2010; 612p. [Google Scholar]
- Petrokas, R.; Baliuckas, V.; Manton, M. Successional categorization of European hemi-boreal forest tree species. Plants 2020, 9, 1381. [Google Scholar] [CrossRef] [PubMed]
- Lithuanian Hydrometeorological Service under the Ministry of Environment. Climate Atlas of Lithuania; Petro ofsetas: Vilnius, Lithuania, 2013. [Google Scholar]
- Lietuvos Hidrometeorologijos Tarnyba [Lithuanian Hydrometeorological Service]. Standartinė Klimato Norma 1991–2020 m. [Standard Climate Normal in 1991–2020]; Lietuvos Hidrometeorologijos Tarnyba: Vilnius, Lithuania, 2021. [Google Scholar]
- Europe’s biodiversity—Biogeographical Regions and Seas. EEA Report 2002 No 1. 2002. Available online: https://www.eea.europa.eu/publications/report_2002_0524_154909 (accessed on 6 November 2022).
- Ministry of Environment, State Forest Service. Lithuanian Statistical Yearbook of Forestry; Butkus, A., Dumčienė, V., Eigirdas, M., Kuliešis, A., Vižlenskas, D., Eds.; Lututė: Kaunas, Lithuania, 2021. [Google Scholar]
- Vaičys, M.; Karazija, S.; Kuliešis, A.; Rutkauskas, A. Miškų Augavietės [Forest Sites]; Lututė: Kaunas, Lithuania, 2006; p. 95. (In Lithuanian) [Google Scholar]
- World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. World Soil Resources Reports No. 106. FAO: Rome, Italy. Available online: https://www.fao.org/3/i3794en/I3794en.pdf (accessed on 16 December 2022).
- Karazija, S. Lietuvos Miškų Tipai [Lithuanian Forest Types]; Mokslas: Vilnius, Lithuania, 1988; p. 212. (In Lithuanian) [Google Scholar]
- Šilingas, M.; Šilingienė, G. The lower storeys of main tree species in deciduous pioneer tree stands of fertile forest sites: Case of Lithuania. Balt. For. 2022, 28, 592. [Google Scholar] [CrossRef]
- Navasaitis, M.; Ozolinčius, R.; Smaliukas, R.; Balevičienė, J. Lietuvos dendroflora [Dendroflora of Lithuania]; Lututė: Kaunas, Lithuania, 2003; 576p, (In Lithuanian with English Summary). [Google Scholar]
- Instructions for Forest Management, Vilnius: Lietuvos Respublikos Seimas. 2010. Updated in 2021. (In Lithuanian). Available online: https://e-seimas.lrs.lt/portal/legalAct/lt/TAD/TAIS.370142/asr (accessed on 22 November 2022).
- Kull, O.; Niinemets, U. Variations in leaf morphometry and nitrogen concentration in Betula pendula Roth., Corylus avellana L. and Lonicera xylosteum L. Tree Physiol. 1993, 12, 311–318. [Google Scholar] [CrossRef]
- Enescu, C.M.; Houston Durrant, T.; de Rigo, D.; Caudullo, G. Corylus avellana in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Huntley, B. Rapid early Holocene migration and high abundance of hazel (Corylus avellana L.): Alternative hypotheses. In Climate Change and Human Impact on the Landscape; Chambers, F.M., Ed.; Chapman & Hall: London, England, 1993. [Google Scholar]
- EUFORGEN—The European Forest Genetic Resources Programme 2022. Corylus avellana. Common hazel. Available online: https://www.euforgen.org/species/corylus-avellana/ (accessed on 10 November 2022).
- Karazija, S. Ąžuolynų bendrijų žemutinių ardų fitocenotinės struktūros kaitos [Changes of phytocenotical structure of lower vegetation layers in oak stand communities]. Miškininkystė 2005, 1, 54–61, (In Lithuanian with English Summary). [Google Scholar]
- Diekmann, M. Deciduous Forest Vegetation in Boreo-Nemoral Scandinavia; Acta Phytogeographica Suecica 80; Opulus Press AB: Uppsala, Sweden, 1994. [Google Scholar]
- Koorem, K.; Moora, M. Positive association between understory species richness and a dominant shrub species (Corylus avellana) in a boreonemoral spruce forest. For. Ecol. Manag. 2010, 260, 1407–1413. [Google Scholar] [CrossRef]
- Deiller, A.-F.; Walter, J.-M.; Tremolieres, M. Regeneration strategies in a temperate hardwood floodplain forest of the Upper Rhine: Sexual versus vegetative reproduction of woody species. For. Ecol. Manag. 2003, 180, 215–225. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Indicator values of plants in Central Europe. Scr. Geobot. 1992, 18, 1–258. [Google Scholar]
- Nestby, R.D.J. The Status of Prunus padus L. (Bird Cherry) in Forest Communities throughout Europe and Asia. Forests 2020, 11, 497. [Google Scholar] [CrossRef]
- Houston Durrant, T.; Caudullo, G. Prunus padus in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Kreyer, D.; Zerbe, S. Short-Lived Tree Species and Their Role as Indicators for Plant Diversity in the Restoration of Natural Forests. Restor. Ecol. 2006, 14, 137–147. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huuskonen, S.; Domisch, T.; Finér, L.; Hantula, J.; Hynynen, J.; Matala, J.; Miina, J.; Neuvonen, S.; Nevalainen, S.; Niemistö, P.; et al. What is the potential for replacing monocultures with mixed-species stands to enhance ecosystem services in boreal forests in Fennoscandia? For. Ecol. Manag. 2021, 479, 118558. [Google Scholar] [CrossRef]
- Allegrini, A.; Salvaneschi, P.; Schirone, B.; Cianfaglione, K.; Di Michele, A. Multipurpose plant species and circular economy: Corylus avellana L. as a study case. Front. Biosci. (Landmark Ed) 2022, 27, 11. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.D.; Thompson, J.H. Effects of logging history on invasion of eastern white pine forests by exotic glossy buckthorn (Frangula alnus P. Mill.). For. Ecol. Manag. 2012, 265, 201–210. [Google Scholar] [CrossRef]
- Cunard, C.E.; Lee, T.D. Is patience a virtue? Succession, light, and the death of invasive glossy buckthorn (Frangula alnus). Biol. Invasions 2008, 11, 577–586. [Google Scholar] [CrossRef]
- Kouki, J.; Arnold, K.; Martikainen, P. Long-term persistence of aspen, a key host for many threatened species, is endangered in old-growth conservation areas in Finland. J. Nat. Conserv. 2004, 12, 41–52. [Google Scholar] [CrossRef]
- Ozolinčius, R.; Lekevičius, E.; Stakėnas, V.; Galvonaitė, A.; Samas, A.; Valiukas, D. Lithuanian forests and climate change: Possible effects on tree species composition. Eur. J. For. Res. 2014, 133, 51–60. [Google Scholar] [CrossRef]








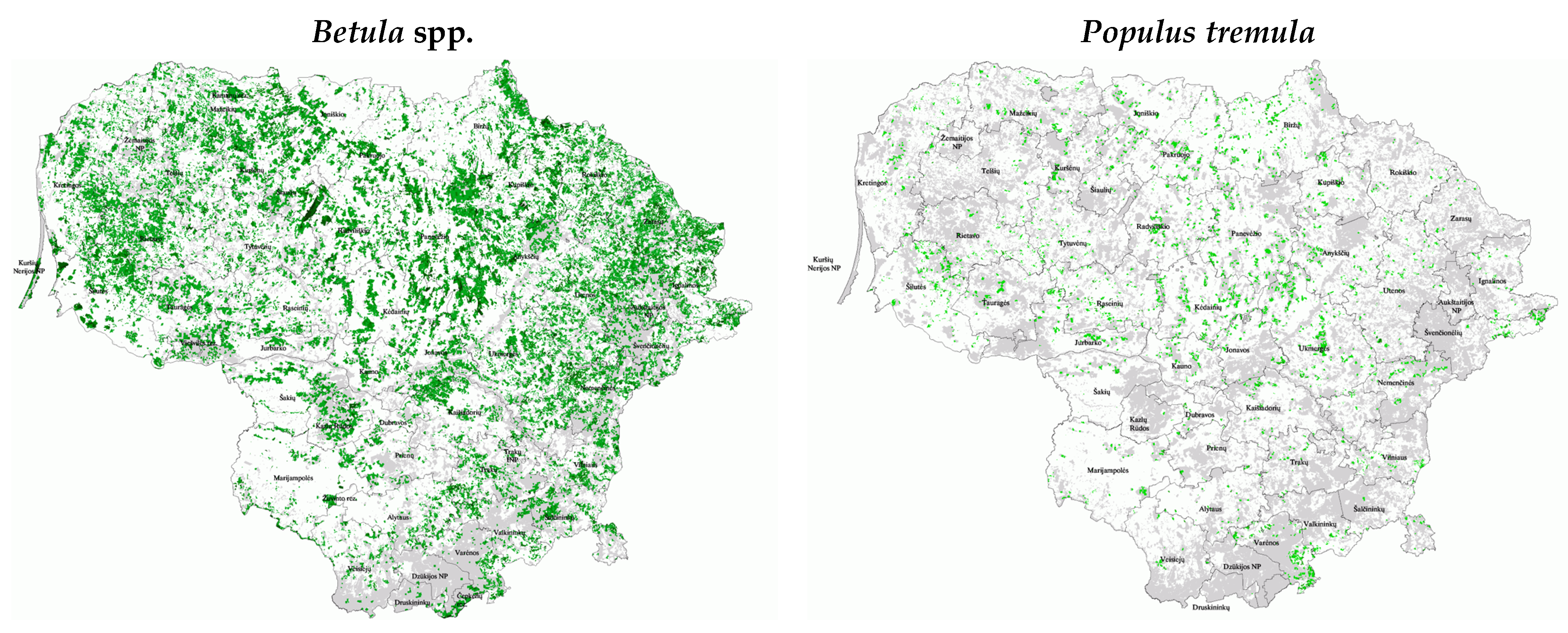{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
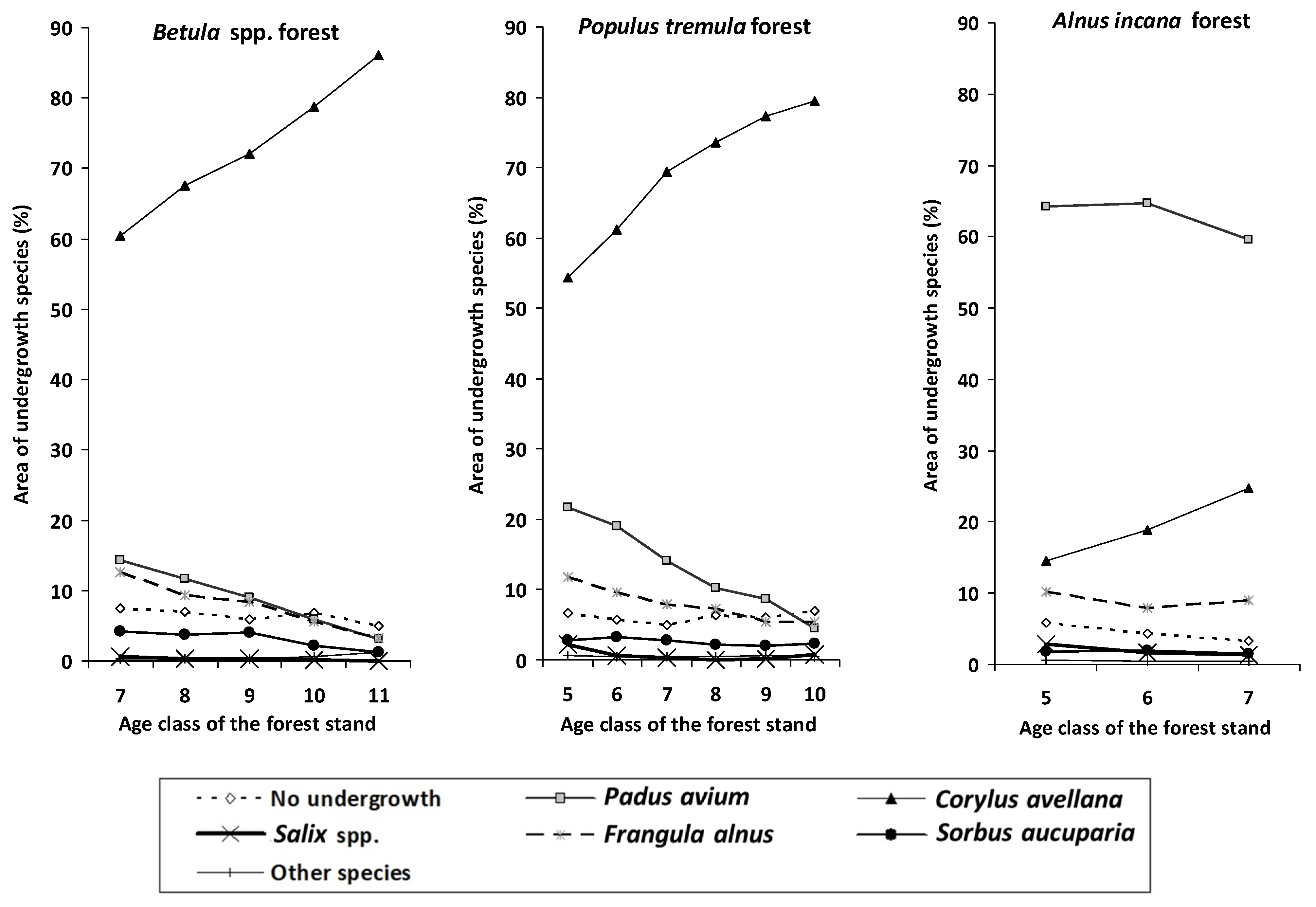{kind=link}
{kind=link}
| Forest Site Type 1: Code–Description | Soil Group 2 | Basic Soil Properties 3 | Dominant Ground Vegetation Species 3 | Undergrowth Species 3 |
|---|---|---|---|---|
| Šd—eutrophic/very fertile mineral soils on slopes (more than 15 degrees of slope) | Cambisols | Forest floor depth 2–5 cm; A horizon * depth 15–20 cm. Forest floor pHKCl 5.5; mineral 0–20 cm topsoil pHKCl 5.0–5.4, and humus amount 2.4–4.8%. Moisture drains through the soil surface. | Hepatica nobilis Mill., Stellaria holostea L., Galeobdolon luteum Huds. | Corylus avellana L., Padus avium L., Euonymus verrucosus Scop., Sorbus aucuparia L., Lonicera xylosteum L. |
| Nd—eutrophic/very fertile mineral soils (relatively high nutrient concentrations) of normal moisture. | Leptosols, Cambisols, Luvisols, Fluvisols | Forest floor with a depth of 2–6 cm gradually transitioning to the A horizon; Bw horizon ** more typical in deciduous forests. Forest floor pHKCl 5.4; mineral 0–20 cm topsoil pHKCl 4.8–5.2, mean C:N ratio 14 ± 1; humus amount 2.6–5.0%. Groundwater level below 2 m. | Hepatica nobilis Mill., Oxalis acetosella L., Galeobdolon luteum Huds., Stellaria holostea L. | Corylus avellana L., Sorbus aucuparia L., Lonicera xylosteum L., Frangula alnus Mill., Daphne mezereum L., Euonymus verrucosus Scop., Viburnum opulus L., Padus avium L., Rhamnus cathartica L. |
| Ld—temporary over-moistened eutrophic/very fertile mineral soils. | Cambisols, Luvisols, Albeluvisols, Gleysols, Fluvisols | Forest floor depth 5–12 cm; A horizon depth 20–25 cm with high root density; Bt horizon *** in some places. Forest floor pHKCl 5.2; mineral 0–20 cm topsoil pHKCl 5.1–5.5, mean C:N ratio 11 ± 1; humus amount 3.1–6.4%. Groundwater level 1.5–2.0 m. | Oxalis acetosella L., Anemone nemorosa L., Aegopodium podagraria L., Geum urbanum L., Galeobdolon luteum Huds. | Sorbus aucuparia L., Corylus avellana L., Frangula alnus Mill., Daphne mezereum L., Lonicera xylosteum L., Padus avium L., Euonymus europaeus L., Ribes nigrum L., Salix cinerea L. |
| Nf—very eutrophic/highly fertile (very high nutrient concentrations) mineral soils of normal moisture. | Cambisols | Forest floor depth 1–3 cm; A horizon depth 20–30 cm; typical Bw horizon. Forest floor pHKCl 5.6; mineral 0–20 cm topsoil pHKCl 5.5–6.0; mean C:N ratio 9 ± 1; humus amount 4.3–7.1%. Groundwater level below 2 m. | Aegopodium podagraria L., Anemone nemorosa L., Asarum europaeum L., Galeobdolon luteum Huds., Stellaria holostea L., Geum urbanum L. | Corylus avellana L., Sorbus aucuparia L., Lonicera xylosteum L., Frangula alnus Mill., Padus avium L., Daphne mezereum L., Euonymus verrucosus Scop., Viburnum opulus L., Ribes nigrum L., Euonymus europaeus L., Rhamnus cathartica L. |
| Lf—temporary over-moistened very eutrophic/highly fertile mineral soils. | Cambisols, Luvisols | Forest floor depth 3–5(8) cm; A horizon depth 25–40 cm. Forest floor pHKCl 5.4; mineral 0–20 cm topsoil pHKCl 5.6–5.7; humus amount 4.6–8.7%. Groundwater level 1.5–2.0 m. | Aegopodium podagraria L., Anemone nemorosa L., Asarum europaeum L., Galeobdolon luteum Huds., Stellaria holostea L., Geum urbanum L. | Frangula alnus Mill., Corylus avellana L., Sorbus aucuparia L., Padus avium L., Rhamnus cathartica L., Euonymus europaeus L., Salix cinerea L., Lonicera xylosteum L., Euonymus verrucosus Scop., Viburnum opulus L., Ribes nigrum L., Daphne mezereum L. |
| Undergrowth Species | Predominant Tree Species of the Main Tree Story | ||
|---|---|---|---|
| Betula spp. | Populus tremula | Alnus incana | |
| Distribution (%) | |||
| No undergrowth | 6.9 a | 5.9 b | 5.1 c |
| Corylus avellana | 67.0 a | 69.2 b | 16.9 c |
| Padus avium | 11.5 a | 13.5 b | 64.1 c |
| Sorbus aucuparia | 3.8 a | 2.6 b | 1.8 c |
| Frangula alnus. | 10.0 a | 7.9 b | 9.3 c |
| Salix spp. | 0.4 a | 0.5 a | 2.3 b |
| Other species * | 0.4 a | 0.5 b | 0.5 b |
| Total area (ha) | 64,976 | 34,380 | 47,615 |
| Undergrowth Species | Betula spp. | Populus tremula | Alnus incana | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Pure | Low Mixed | Mixed | Pure | Low Mixed | Mixed | Pure | Low Mixed | Mixed | |
| Distribution (%) | |||||||||
| No undergrowth | 9.9 a | 6.9 b | 6.1 c | 7.3 a | 5.9 b | 5.6 b | 5.4 a | 5.2 ab | 4.7 b |
| Corylus avellana | 56.0 a | 63.8 b | 71.4 c | 66.2 a | 67.4 a | 70.4 b | 8.9 a | 16.7 b | 26.2 c |
| Padus avium | 14.9 a | 12.7 b | 10.1 c | 14.2 ab | 14.3 a | 13.0 b | 72.5 a | 63.4 b | 55.1 c |
| Sorbus aucuparia | 4.5 a | 4.0 a | 3.6 b | 1.6 a | 2.9 b | 2.6 b | 1.6 a | 2.1 b | 1.7 a |
| Frangula alnus | 13.7 a | 11.7 b | 8.1 c | 9.9 a | 8.6 a | 7.3 b | 9.1 a | 9.3 a | 9.4 a |
| Salix spp. | 0.6 a | 0.5 a | 0.4 b | 0.4 a | 0.4 a | 0.5 a | 1.9 a | 2.6 b | 2.5 b |
| Other species | 0.4 a | 0.4 a | 0.3 a | 0.5 a | 0.4 a | 0.6 a | 0.5 a | 0.6 a | 0.4 a |
| Total area (ha) | 9371 | 18,649 | 36,956 | 3599 | 9551 | 21,230 | 3599 | 9551 | 21,230 |
| Undergrowth Species | Forest Site Types | ||||
|---|---|---|---|---|---|
| Šd* | Nd | Ld | Nf | Lf | |
| Distribution in Betula spp. forests (%) | |||||
| No undergrowth | 5.6 a | 9.2 b | 7.0 a | 7.9 ab | 2.7 c |
| Corylus avellana | 69.7 a | 65.8 ab | 64.4 b | 82.4 c | 82.5 c |
| Padus avium | 12.0 abc | 13.5 c | 11.4 b | 6.5 d | 10.3 a |
| Sorbus aucuparia | 5.3 ab | 5.4 b | 4.0 a | 0.6 c | 0.5 c |
| Frangula alnus. | 4.1 ab | 5.0 b | 12.4 c | 1.5 d | 3.7 a |
| Salix spp. | 0.4 ab | 0.1 bc | 0.6 a | 0.0 abc | 0.1 c |
| Other | 3.0 a | 0.9 b | 0.2 c | 1.1 ab | 0.2 c |
| Total (ha) | 1009 | 10,446 | 45,194 | 818 | 7509 |
| Distribution in Populus tremula forests (%) | |||||
| No undergrowth | 8.3 a | 6.7 a | 6.1 a | 1.3 b | 2.8 b |
| Corylus avellana | 66.9 ab | 70.9 b | 66.3 a | 83.1 c | 79.7 c |
| Padus avium | 14.7 ab | 14.4 b | 13.1 a | 13.1 ab | 13.8 ab |
| Sorbus aucuparia | 1.8 a | 3.0 a | 2.9 a | 0.0 b | 0.2 b |
| Frangula alnus. | 3.3 a | 3.6 a | 10.8 b | 2.1 a | 3.0 a |
| Salix spp. | 0.6 ab | 0.3 b | 0.6 a | 0.0 ab | 0.3 ab |
| Other | 4.3 a | 1.2 b | 0.2 c | 0.4 bc | 0.2 c |
| Total (ha) | 828 | 7869 | 21,163 | 467 | 4054 |
| Distribution in Alnus incana forests (%) | |||||
| No undergrowth | 5.4 a | 3.9 b | 5.5 a | 2.9 ab | 4.0 ab |
| Corylus avellana | 25.3 a | 21.1 b | 13.2 c | 33.3 ad | 31.6 d |
| Padus avium | 57.1 a | 64.6 b | 65.7 b | 56.9 ab | 57.6 a |
| Sorbus aucuparia | 0.9 a | 3.7 b | 1.5 c | 1.0 abc | 0.0 d |
| Frangula alnus. | 7.7 a | 4.8 b | 11.0 c | 5.9 abc | 5.1 b |
| Salix spp. | 2.4 a | 1.1 b | 2.7 a | 0.0 ab | 1.6 ab |
| Other | 1.2 a | 0.8 ab | 0.3 c | 0.0 abc | 0.1 bc |
| Total (ha) | 6916 | 8782 | 30,740 | 102 | 1076 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šilingas, M.; Suchockas, V.; Varnagirytė-Kabašinskienė, I. Evaluation of Undergrowth under the Canopy of Deciduous Forests on Very Fertile Soils in the Lithuanian Hemiboreal Forest. Forests 2022, 13, 2172. https://doi.org/10.3390/f13122172
Šilingas M, Suchockas V, Varnagirytė-Kabašinskienė I. Evaluation of Undergrowth under the Canopy of Deciduous Forests on Very Fertile Soils in the Lithuanian Hemiboreal Forest. Forests. 2022; 13(12):2172. https://doi.org/10.3390/f13122172
Chicago/Turabian StyleŠilingas, Marius, Vytautas Suchockas, and Iveta Varnagirytė-Kabašinskienė. 2022. "Evaluation of Undergrowth under the Canopy of Deciduous Forests on Very Fertile Soils in the Lithuanian Hemiboreal Forest" Forests 13, no. 12: 2172. https://doi.org/10.3390/f13122172