
Volumetric Spore Traps Are a Viable Alternative Tool for Estimating Heterobasidion Infection Risk

Abstract
:1. Introduction
2. Materials and Methods
2.1. Air Sampling
2.2. Identification of Heterobasidion annosum s.l.
2.3. DNA Extraction from Air Samples and qPCR
2.4. Absolute Quantification
2.5. Statistical Analyses
3. Results
3.1. Performance of qPCR Assays
3.2. Comparison of Spore Trapping Methods
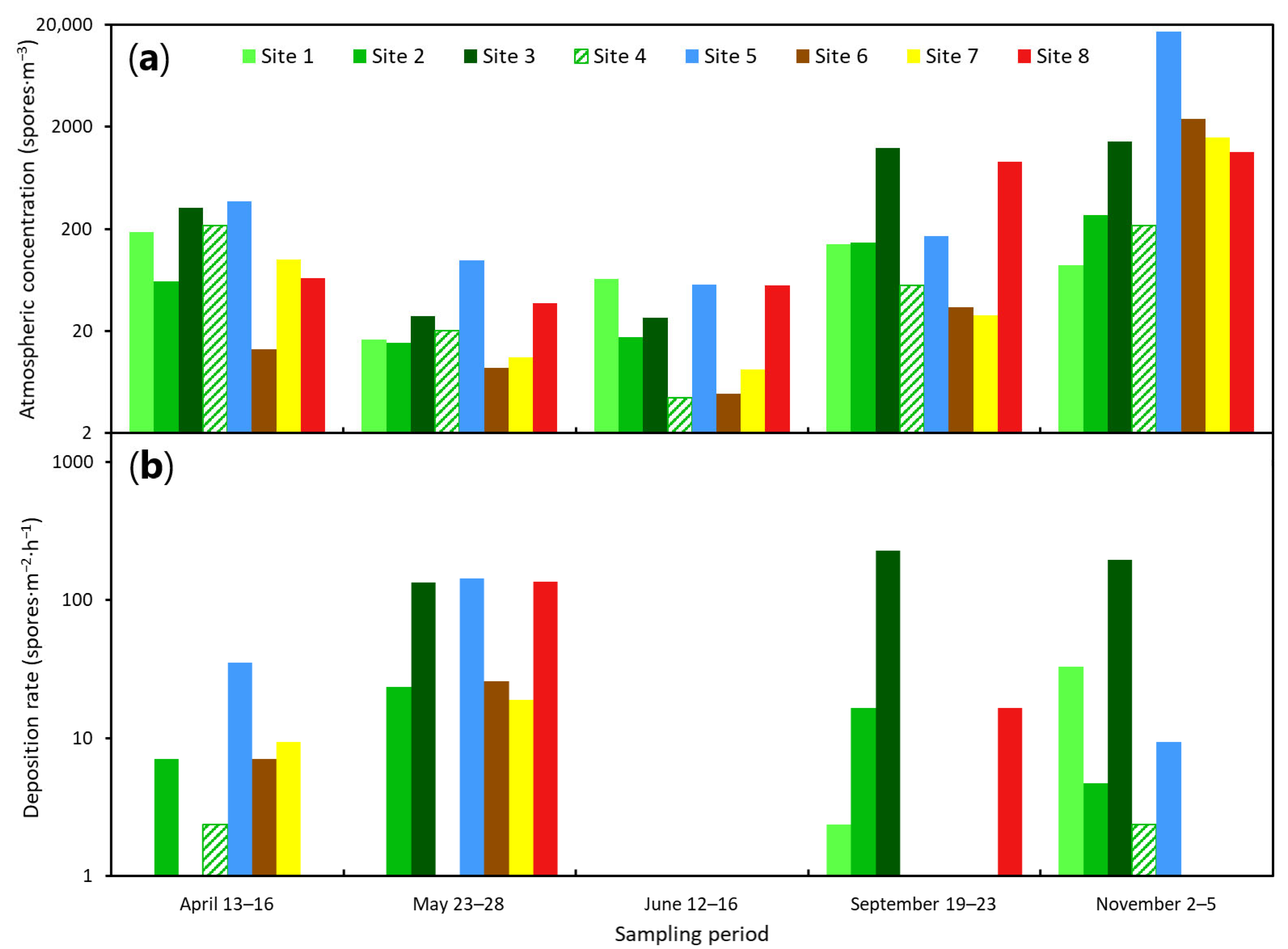
3.3. Effects of Site, Season, and Weather on the Abundance of Heterobasidion annosum s.l. Airspora
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asiegbu, F.O.; Adomas, A.; Stenlid, J. Conifer root and butt rot caused by Heterobasidion annosum (Fr.) Bref. s.l. Mol. Plant Pathol. 2005, 6, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Gonthier, P.; Thor, M. Annosus root and butt rots. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CABI: Wallingford, UK, 2013; pp. 128–158. [Google Scholar]
- Honkaniemi, J.; Ahtikoski, A.; Piri, T. Financial incentives to perform stump treatment against Heterobasidion root rot in Norway spruce dominated forests, the case of Finland. For. Policy Econ. 2019, 105, 1–9. [Google Scholar] [CrossRef]
- Kallio, T. Aerial Distribution of the Root-Rot Fungus Fomes annosus (Fr.) Cooke in Finland. Acta For. Fenn. 1970, 107, 55. [Google Scholar] [CrossRef] [Green Version]
- Stanosz, G.R.; Smith, D.R.; Juzwik, J. Seasonal availability of inoculum of the Heterobasidion root disease pathogen in central Wisconsin. Can. J. For. Res. 2016, 46, 1076–1080. [Google Scholar] [CrossRef] [Green Version]
- Hirst, J.M. An automatic volumetric spore trap. Ann. Appl. Biol. 1952, 39, 257–265. [Google Scholar] [CrossRef]
- Grinn-Gofroń, A.; Strzelczak, A.; Przestrzelska, K. Seasonal variation of Ganoderma spore concentrations in urban and suburban districts of the city of Szczecin, Poland. Ann. Agric. Environ. Med. 2015, 22, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Aguayo, J.; Husson, C.; Chancerel, E.; Fabreguettes, O.; Chandelier, A.; Fourrier-Jeandel, C.; Dupuy, N.; Dutech, C.; Ioos, R.; Robin, C.; et al. Combining permanent aerobiological networks and molecular analyses for large-scale surveillance of forest fungal pathogens: A proof-of-concept. Plant Pathol. 2021, 70, 181–194. [Google Scholar] [CrossRef]
- Dvořák, M.; Drápela, K.; Jankovský, L. Dothistroma septosporum: Spore production and weather conditions. For. Syst. 2012, 21, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, M.; Rotkova, G.; Botella, L. Detection of Airborne Inoculum of Hymenoscyphus fraxineus and H. albidus during Seasonal Fluctuations Associated with Absence of Apothecia. Forests 2015, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Hietala, A.M.; Timmermann, V.; Børja, I.; Solheim, H. The invasive ash dieback pathogen Hymenoscyphus pseudoalbidus exerts maximal infection pressure prior to the onset of host leaf senescence. Fungal Ecol. 2013, 6, 302–308. [Google Scholar] [CrossRef]
- Timmermann, V.; Børja, I.; Hietala, A.M.; Kirisits, T.; Solheim, H. Ash dieback: Pathogen spread and diurnal patterns of ascospore dispersal, with special emphasis on Norway. EPPO Bull. 2011, 41, 14–20. [Google Scholar] [CrossRef]
- Lacey, M.E.; West, J.S. The Air Spora; Springer: Dordrecht, The Netherlands, 2006; p. 156. [Google Scholar]
- Sinclair, W.A.; Lyon, H.H. Diseases of Trees and Shrubs, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 2005; p. 680. [Google Scholar]
- Chandelier, A.; Hulin, J.; San Martin, G.; Debode, F.; Massart, S. Comparison of qPCR and Metabarcoding Methods as Tools for the Detection of Airborne Inoculum of Forest Fungal Pathogens. Phytopathology 2021, 111, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Gonthier, P.; Garbelotto, M.M.; Varese, G.C.; Nicolotti, G. Relative abundance and potential dispersal range of intersterility groups of Heterobasidion annosum in pure and mixed forests. Can. J. Bot. 2001, 79, 1057–1065. [Google Scholar] [CrossRef]
- Stenlid, J. Regional differentiation in Heterobasidion annosum. In Proceedings of the Eighth International Conference on Root and Butt Rots, Wik, Sweden and Haikko, Finland, 9–16 August 1993; Johansson, M., Stenlid, J., Eds.; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1994; pp. 243–248. [Google Scholar]
- James, R.L.; Cobb, F.W., Jr. Spore Deposition by Heterobasidion annosum in Forests of California. Plant Dis. 1984, 68, 246–248. [Google Scholar] [CrossRef]
- Gonthier, P.; Garbelotto, M.M.; Nicolotti, G. Seasonal Patterns of Spore Deposition of Heterobasidion Species in Four Forests of the Western Alps. Phytopathology 2005, 95, 759–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dálya, L.B.; Sedlák, P. Prevalence of major wood-decay Agaricomycetes in artificial, managed near-natural and undisturbed forests of South Moravia, Czechia. For. Pathol. 2020, 50, e12636. [Google Scholar] [CrossRef]
- Johannesson, H.; Stenlid, J. Molecular markers reveal genetic isolation and phylogeography of the S and F intersterility groups of the wood-decay fungus Heterobasidion annosum. Mol. Phylogenet. Evol. 2003, 29, 94–101. [Google Scholar] [CrossRef]
- Oliva, J.; Messal, M.; Wendt, L.; Elfstrand, M. Quantitative interactions between the biocontrol fungus Phlebiopsis gigantea, the forest pathogen Heterobasidion annosum and the fungal community inhabiting Norway spruce stumps. For. Ecol. Manag. 2017, 402, 253–264. [Google Scholar] [CrossRef]
- Ioos, R.; Chrétien, P.; Perrault, J.; Jeandel, C.; Dutech, C.; Gonthier, P.; Sillo, F.; Hietala, A.M.; Solheim, H.; Hubert, J. Multiplex real-time PCR assays for the detection and identification of Heterobasidion species attacking conifers in Europe. Plant Pathol. 2019, 68, 1493–1507. [Google Scholar] [CrossRef]
- Klymus, K.E.; Merkes, C.M.; Allison, M.J.; Goldberg, C.S.; Helbing, C.C.; Hunter, M.E.; Jackson, C.A.; Lance, R.F.; Mangan, A.M.; Monroe, E.M.; et al. Reporting the limits of detection and quantification for environmental DNA assays. Environ. DNA 2020, 2, 271–282. [Google Scholar] [CrossRef]
- Canales, R.A.; Wilson, A.M.; Pearce-Walker, J.I.; Verhougstraete, M.P.; Reynolds, K.A. Methods for Handling Left-Censored Data in Quantitative Microbial Risk Assessment. Appl. Environ. Microbiol. 2018, 84, e01203-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redfern, D.B.; Stenlid, J. Spore dispersal and infection. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CABI: Wallingford, UK, 1998; pp. 105–124. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2022. Available online: https://www.R-project.org/ (accessed on 18 September 2022).
- Brūna, L.; Lione, G.; Kenigsvalde, K.; Burņeviča, N.; Zaļuma, A.; Kļaviņa, D.; Gaitnieks, T.; Gonthier, P. Inferences on the Susceptibility of Wood of Different Tree Species to Heterobasidion annosum Sensu Lato Primary Infections and on the Range of Pathogen Spores Dispersal. Forests 2021, 12, 854. [Google Scholar] [CrossRef]
- Oliva, J.; Bendz-Hellgren, M.; Stenlid, J. Spread of Heterobasidion annosum s.s. and Heterobasidion parviporum in Picea abies 15 years after stump inoculation. FEMS Microbiol. Ecol. 2011, 75, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Zaļuma, A.; Gaitnieks, T.; Arhipova, N.; Vasaitis, R. Growth rates of Heterobasidion annosum s.s. and H. parviporum in functional sapwood of Pinus sylvestris and Picea abies. For. Pathol. 2015, 45, 437–439. [Google Scholar] [CrossRef]
- Jackson, S.L.; Bayliss, K.L. Spore traps need improvement to fulfil plant biosecurity requirements. Plant Pathol. 2011, 60, 801–810. [Google Scholar] [CrossRef]
- Redondo, M.Á.; Berlin, A.; Boberg, J.; Oliva, J. Vegetation type determines spore deposition within a forest-agricultural mosaic landscape. FEMS Microbiol. Ecol. 2020, 96, fiaa082. [Google Scholar] [CrossRef]
- Hsiang, T.; Edmonds, R.L.; Driver, C.H. Conidia of Heterobasidion annosum from Tsuga heterophylla forests in western Washington. Can. J. Bot. 1989, 67, 1262–1266. [Google Scholar] [CrossRef] [Green Version]
- Möykkynen, T. Liberation of Heterobasidion annosum conidia by airflow. For. Pathol. 1997, 27, 283–289. [Google Scholar] [CrossRef]
- Hunt, R.S.; Wilcox, W.W.; Cobb Jr., F. W. Resistance of Stump Tops to Colonization by Fomes annosus. Can. J. For. Res. 1974, 4, 140–142. [Google Scholar] [CrossRef]
- Kuhlman, E.G. Number of conidia necessary for stump root infection by Fomes annosus. Phytopathology 1969, 59, 1168–1169. [Google Scholar]
- Möykkynen, T.; Von Weissenberg, K.; Pappinen, A. Estimation of dispersal gradients of S- and P-type basidiospores of Heterobasidion annosum. For. Pathol. 1997, 27, 291–300. [Google Scholar] [CrossRef]
- Möykkynen, T.; Kontiokari, J. Spore deposition of Heterobasidion annosum coll. in Picea abies stands of North Karelia, eastern Finland. For. Pathol. 2001, 31, 107–114. [Google Scholar] [CrossRef]
- Sylvestre-Guinot, G.; Delatour, C. Recherches sur les variations saisonnières de l’inoculum aérien du Fomes annosus (Fr.) Cooke dans l’est de la France. Ann. For. Sci. 1978, 35, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Bérubé, J.A.; Potvin, A.; Stewart, D. Importance of local and long-distance Heterobasidion irregulare aerial basidiospore dispersal for future infection centres in thinned red pine plantations in Quebec. For. Chron. 2017, 93, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Otto, E.; Held, B.; Redford, S.; Blanchette, R.A. Detecting Heterobasidion irregulare in Minnesota and Assessment of Indigenous Fungi on Pines. Forests 2021, 12, 57. [Google Scholar] [CrossRef]


{kind=link}
| Site | Latitude N | Longitude E | Stand Age (Years) | Dominant Tree Species | Management | Heterobasidion Species Present 1 |
|---|---|---|---|---|---|---|
| 1 | 49.300284 | 16.601441 | 12 | Picea abies | managed | abietinum |
| 2 | 49.269397 | 16.684964 | 45 | Picea abies | managed | annosum |
| 3 | 49.325002 | 16.719278 | 107 | Picea abies | managed | annosum |
| 4 | 49.070277 | 15.674212 | 62 | Picea abies | managed | annosum |
| 5 | 48.912336 | 17.158900 | 71 | Pinus sylvestris | managed | annosum |
| 6 | 49.222491 | 16.673499 | 0–149 | Quercus petraea | unmanaged | - |
| 7 | 49.283868 | 16.734009 | 0–49 | Fagus sylvatica | unmanaged | - |
| 8 | 49.304245 | 16.696342 | 0–146 | mixed 2 | unmanaged | - |
| Target | LOD in Ioos et al. [23], Nuclei/Rxn | LOD as a Discrete Threshold | LOD with Curve-Fitting Modeling | LOQ with Curve-Fitting Modeling | |||
|---|---|---|---|---|---|---|---|
| Spores/Sample | Nuclei/Rxn | Spores/Sample | Nuclei/Rxn | Spores/Sample | Nuclei/Rxn | ||
| HA | nd | 100 * | ≤7.5 | nd | nd | nd | nd |
| Ab | ~30 | 1000 | ≤75 | 589 | ≤45 | 5460 | ≤410 |
| An | 311 | 1000 | ≤75 | 589 | ≤45 | 3297 | ≤248 |
| Pa | 29 | 1000 | ≤75 | 589 | ≤45 | 2368 | ≤178 |
| Site | April | May | June | September | November | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AVST | WDE | R | AVST | WDE | R | AVST | WDE | R | AVST | WDE | R | AVST | WDE | R | |||||||||||||||||||||
| Ab | An | Pa | Ab | An | Pa | Ab | An | Pa | Ab | An | Pa | Ab | An | Pa | Ab | An | Pa | Ab | An | Pa | Ab | An | Pa | Ab | An | Pa | Ab | An | Pa | ||||||
| 1 | + | ++ | − | − | − | − | A | o | o | o | − | − | − | A | ++ | + | + | − | − | − | A | ++ | ++ | + | o | o | o | A | o | o | o | ++ | − | − | W |
| 2 | + | ++ | ++ | o | o | o | nd | − | ++ | − | − | − | ++ | I | − | ++ | − | − | − | − | A | ++ | + | − | − | − | ++ | I | − | ++ | + | ++ | − | − | I |
| 3 | + | ++ | + | − | − | − | A | ++ | + | ++ | − | − | ++ | A | + | + | ++ | − | − | − | A | ++ | + | + | + | − | ++ | A | + | ++ | ++ | ++ | + | − | A |
| 4 | − | ++ | − | o | o | o | nd | ++ | ++ | ++ | − | − | − | A | − | ++ | − | − | − | − | A | − | + | ++ | − | − | − | A | − | ++ | − | o | o | o | A |
| 5 | − | ++ | − | o | o | o | nd | − | ++ | − | − | + | ++ | W | − | ++ | − | − | − | − | A | + | ++ | − | − | − | − | A | − | ++ | − | ++ | − | − | I |
| 6 | o | o | o | o | o | o | nd | − | ++ | − | − | − | ++ | I | − | − | ++ | − | − | − | A | ++ | − | ++ | − | − | − | A | − | ++ | − | − | − | − | A |
| 7 | − | ++ | − | o | o | o | nd | ++ | − | − | − | − | ++ | I | o | o | o | − | − | − | A | ++ | − | ++ | − | − | − | A | − | ++ | − | − | − | − | A |
| 8 | ++ | + | ++ | − | − | − | A | + | − | ++ | − | − | ++ | A | ++ | + | ++ | − | − | − | A | + | + | ++ | ++ | − | + | A | + | + | ++ | − | − | − | A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dálya, L.B.; Dvořák, M.; Sedlák, P. Volumetric Spore Traps Are a Viable Alternative Tool for Estimating Heterobasidion Infection Risk. Forests 2022, 13, 2146. https://doi.org/10.3390/f13122146
Dálya LB, Dvořák M, Sedlák P. Volumetric Spore Traps Are a Viable Alternative Tool for Estimating Heterobasidion Infection Risk. Forests. 2022; 13(12):2146. https://doi.org/10.3390/f13122146
Chicago/Turabian StyleDálya, László Benedek, Miloň Dvořák, and Petr Sedlák. 2022. "Volumetric Spore Traps Are a Viable Alternative Tool for Estimating Heterobasidion Infection Risk" Forests 13, no. 12: 2146. https://doi.org/10.3390/f13122146