1. Introduction
Seed production in many temperate tree species is subject to substantial interannual variability, which depends on mast events separated by seasons with little seed yield [
1] and high variability among individuals [
2]. Thus, to ensure the continuous operation of forest nurseries, seeds may need to be stored over several seasons [
3]. Long-term storage has, however, been problematic in the case of seeds representing the recalcitrant category [
4], i.e., those that do not tolerate desiccation, such as the seeds of oaks. Pedunculate oak (
Quercus robur L.) [
5] is an important species native to Europe and in the future it is expected to experience high mortality related to climate changes [
6]. The storage of oak acorns invariably results in a decline in viability: a reduced germination rate and lower seedling vigor [
5,
7,
8,
9]. Storage conditions, i.e., temperature and seed moisture content, are two major determinants of the half-life of seeds [
10]. Therefore, improvements in storage protocols will contribute to the reduction of the process of seed deterioration [
11] and to successful genebanking [
3].
The mechanisms of the deterioration of vitality in recalcitrant seeds during storage have not been fully explained. Nevertheless, they have been attributed to several processes, including structural damage to the vacuole, cytoskeleton, and membrane organization, changes in the state of macromolecules, metabolic damage mediated by the production of reactive oxygen species [
12], and infections with pathogens [
13]. Short-term storage (from several weeks to several months) of recalcitrant seeds is possible in a soil seed bank under controlled optimal conditions [
14]. However, even under short-term storage in optimal conditions, pedunculate oak seeds display a provenance-dependent reduction in vitality due to the sequential accumulation of reactive oxygen species resulting from storage [
15]. Apart from oxidation reactions, alcoholic fermentation may be involved in seed deterioration during aging and desiccation stress of pedunculate oak seeds [
16]. Metabolomic analyses revealed that the loss of viability in pedunculate oak seeds is caused by failure to activate prosurvival mechanisms during storage [
17].
Physiological studies of seed deterioration during storage tend to focus on embryonic axes; however, storage tissues—endosperm or cotyledons—often constitute major seed compartments. Cotyledons of some recalcitrant seeds are more sensitive to dehydration than embryonic axes [
18] and, therefore, more prone to deterioration. In oak seeds, the reserve materials predominantly include starch, which constitutes up to 55% of the dry weight in
Quercus spp. [
19], and a smaller quantity of lipids and proteins, all stored within the large cotyledons. Reserves are mobilized during germination and seedling establishment [
20,
21,
22]; thus, their availability is a prerequisite for the success of a seedling. On the other hand, experiments revealed that partial removal of cotyledons does not affect acorn survival and seedling development [
23]. Moreover, a reduction in
Q. robur cotyledons by one-fifth accelerated their emergence and increased the germination rate [
24]. Such experiments, however, remove the distal parts of the cotyledons, while reserves contained in the apical parts may be more accessible to the embryonic organs. In principle, the storage regime should attempt to inhibit premature breakdown and respiratory use of seed reserves. For orthodox seeds, this is efficiently achieved by dry storage [
10]. However, the tissues of recalcitrant seeds must remain partly hydrated during storage, implying sustained respiratory and other metabolic activity [
25,
26]. Reserve materials might also be depleted if recalcitrant seeds experience stress during storage and respond to it by accumulating defensive and anti-stress compounds [
17]. Heat generation occasionally reported from stored recalcitrant and non-recalcitrant seed batches indicates that reserves are indeed being depleted, both by seed respiration and accompanying metabolic activities as well as by microorganismal activity [
27]. Storage methods for acorns, as well as for other recalcitrant seeds, must balance both the requirements for the maintenance of embryo vitality and the retention of seed reserves.
In practice, acorns are stored for one year; however, it was reported that storage for 2 or even 3 years with a relatively low reduction in germination rate is possible [
28]. Temperature reduction has been effective in extending the storage potential of some recalcitrant seeds [
12] and, in the short to medium term, an alternative to the laborious and expensive cryopreservation [
25]. In the storage of temperate oak,
Q. robur and
Q. petraea (Matt.) Liebl. acorns, low temperature has been beneficial; however, below-freezing temperatures may cause damage to hydrated seed tissue, even though acorns of both species are frost-tolerant in nature (down to −9 °C) [
17,
29]. Moreover, even storing acorns at −2 °C did not prevent the consumption of reserve carbohydrates [
20].
A temperature of −3 °C and a moisture content (MC) of 40%–42% are recommended for the storage of pedunculate oak seeds [
28,
30]; however, many attempts have been made to further improve the storage protocols of oak acorns. The loss of viability in acorns is determined by a critical MC in cotyledons [
5]. Oak seeds show tolerance to temperatures down to −9 °C, which is the threshold for cellular water freezing [
29]. Constant storage temperatures (−3, −5, −7 °C) and their combinations (−3 → −5 °C or −3 → −5 → −7 °C) enabling seed acclimation to cold have also been evaluated [
15]. In practice, gradual acclimation procedures [
15] appear to be the most promising approaches for the extension of the storage time of pedunculate oak seeds. Therefore, a slight acorn drying to below the recommended 40% MC followed by storage at subzero temperature may exert a protective effect on seed survival [
29].
The objective of this study was to investigate the effect of storage regime modification on seed viability and the stability of reserves in cotyledons of pedunculate oak acorns during 18 months of storage under controlled moisture and temperature. We used a factorial experiment combining −3 °C or −5 °C storage temperatures with 40% (control) or 38% (dehydrated) MC. We hypothesized that the reduction in temperature below the recommended level combined with a decrease in seed MC would reduce the rate of reserve depletion during storage and be beneficial for the long-term storage of acorns.
2. Materials and Methods
2.1. Seed Material
Acorns of Q. robur were collected in the Wejherowo forest district in northern Poland (54°36′15″ N 18°14′56″ E) in September 2019. Prior to the beginning of the experiment, acorns were divided into two lots: control, which consisted of undried seeds (40% moisture content), and dehydrated, in which MC was adjusted to 38% by drying on trays at room temperature (20 °C) for 5 days.
2.2. Seed Storage
Seeds representing each MC (control and dehydrated) were placed in two separate 80 L barrels, each filled up to ¾ barrel height. A perforated tube was placed in the barrel to ensure proper aeration of the acorns. Barrels were kept in chambers with controlled temperature at −3 or −5 °C. Seed subsamples for viability assessment and reserve compound determination were removed after 0 (control), 4, 8, 12, and 18 months.
2.3. Seed MC Determination
Acorns were sampled at the beginning of the experiment and at the end of each storage interval. They were cut lengthwise into halves and dissected into pericarp, cotyledons, and embryonic axes. In each treatment,
N = 3 replicates were created for each fraction, containing 5 individual embryonic axes, 10 cotyledons, or 10 pericarps. Batches of 30 whole acorns were also assessed. Samples were dried at 103 ± 2 °C for 17 h in a forced circulation oven. The MC of the seeds was assessed using the gravimetric method according to the formula:
where FW is the fresh weight and DW is the dry weight of the sample.
2.4. Seed Germination, Seedling Establishment, and Seedling Biomass
Testing was performed at the beginning of the experiment and at the end of each storage interval. The seeds were germinated at a constant temperature of 20 °C under an 8/16 h (darkness/light) photoperiod, with light at 80 μmol m
−2 s
−1 supplied by fluorescent tubes. To ensure synchronized germination, prior to sowing, acorns were cut open, terminal 1/3 of the cotyledons was removed, and 2/3 of the seed containing the embryonic axis was sown, as described by Giertych and Suszka [
24]. After each storage interval, three batches of 30 seeds were sown in individual trays containing a mixture of quartz sand and peat (1:1
v/
v). The emerging roots (germination) and shoots (emergence) were counted every two days for over six weeks. Seeds with radicle length ≥ 2 mm were considered germinated or emerged when ≥2 mm of epicotyl was visible. After three months, the effect of storage on seedling quality was evaluated. Each emerged plant was dried at 65 °C for 72 h and weighed to obtain seedling dry mass (seedling DM).
2.5. Sampling for Carbohydrate and Fat Analyses
On the initial day of the experiment (18 December 2019), 27 batches of 30 randomly picked acorns from each moisture level were prepared and weighed. Three batches were immediately used for MC determination, and the remaining batches were placed in individual mesh bags and allocated to storage barrels. On each sampling day, three bags were removed from each barrel. Acorns were weighed, dried as described above, and weighed again. The pericarps were removed, and whole seeds or individual cotyledons bearing visible fungal growth were counted (with each cotyledon treated as ½ seed) and discarded. The largest number of infected acorns in a batch was nine. The remaining material was weighed and ground using an MF10 mill (IKA, Staufen, Germany) fitted with a 1 mm sieve.
2.6. Carbohydrate Analysis
Aliquots of 20 mg of pulverized material were subjected to extraction in a mixture of methanol, chloroform, and water. Soluble carbohydrates were determined spectrophotometrically using the anthrone reaction at λ = 625 nm. For starch determination, the residue was boiled in NaF buffer (pH 4.5) at 100 °C for 15 min, followed by digestion with amyloglucosidase and reaction with peroxidase-glucose oxidase. The color reaction was developed using a reaction with o-dianisidine, and the glucose concentration was measured spectrophotometrically against a glucose standard.
2.7. Fat Analysis
Fat was determined in 10 g of ground tissue using the Soxhlet apparatus. Hexan was used as the extraction medium. Each determination was conducted twice.
2.8. Calculations
The sum of soluble carbohydrates and starch was considered to be equivalent to the total nonstructural carbohydrate (TNC). Concentrations of soluble carbohydrates, starch, TNC, and fat were expressed on a dry mass basis. However, since dry mass changed during storage and some seeds or cotyledons were rejected because of infection, we standardized the results by calculating the biomass and pool sizes of reserve compounds per 100 healthy acorns, according to the formulas:
where M
H is the mass of healthy seeds recovered from the sample and N
H is the number of acorns in the sample after discarding the infected cotyledons.
2.9. Statistics
The experiment was designed as a 2 × 2 × 5 factorial consisting of two MC (38% and 40%) and two temperature (−3 °C and −5 °C) levels, with five sampling dates. Samples from Day 0, i.e., prior to storage, were not originally assigned to the different temperature levels; therefore, to obtain a complete factorial, we randomly assigned the results for those samples to the −3 or −5 °C treatments (within the original moisture levels). Three-way analysis of variance was applied to evaluate the effect of sampling date, temperature, and initial acorn moisture content on sample biomass, biomass of 100 healthy acorns, and concentrations and pool sizes of each reserve compound. Biomass and reserve pool sizes were log transformed and germination/seedling emergence arc-sine transformed before analysis. Test assumptions were checked using diagnostic plots. Differences between treatments in initial moisture content were evaluated using Student’s t test. We computed a matrix of Pearson’s correlation coefficients using the corrplot package [
31]. Statistical calculations were performed using JMP software (SAS Institute, Cary, NC, USA) and R statistical computing software [
32].
3. Results
Freshly harvested acorns were characterized by 40% MC and had relatively high germination capacity (82.2 ± 4.4) and seedling emergence rate (68.9 ± 2.9). At the beginning of the experiment, dehydrated acorns showed 38% MC and germinated/emerged in 89%/74%. The hydration levels of each acorn part differed (
Figure 1a). The MC of cotyledons was similar to that of whole acorns, whereas the average MC of pericarp ranged from 24 to 27%, and the MC of embryonic axes ranged from 50.6 to 53.7% on average. Each morphological fraction (cotyledons, pericarp, and embryonic axis) of the 38% MC acorns was significantly less hydrated than the corresponding fraction of the 40% MC acorns.
The MC of both whole acorns and cotyledons did not change during storage at −5 °C (
Figure 1b), whereas in seeds stored at −3 °C, a significant increase was recorded. The MC of the pericarps increased in a similar way independent of storage temperature. Interestingly, MC in the embryonic axis did not change in seeds stored at −3 °C but decreased in seeds stored at −5 °C. A significant difference between the MC of cotyledons was recorded after the 8 mo of storage at −5 °C (
Figure S1). This was further reflected in whole seeds stored for 12 mo at −5 °C. No difference was reported in cotyledons and whole seeds stored at −3 °C.
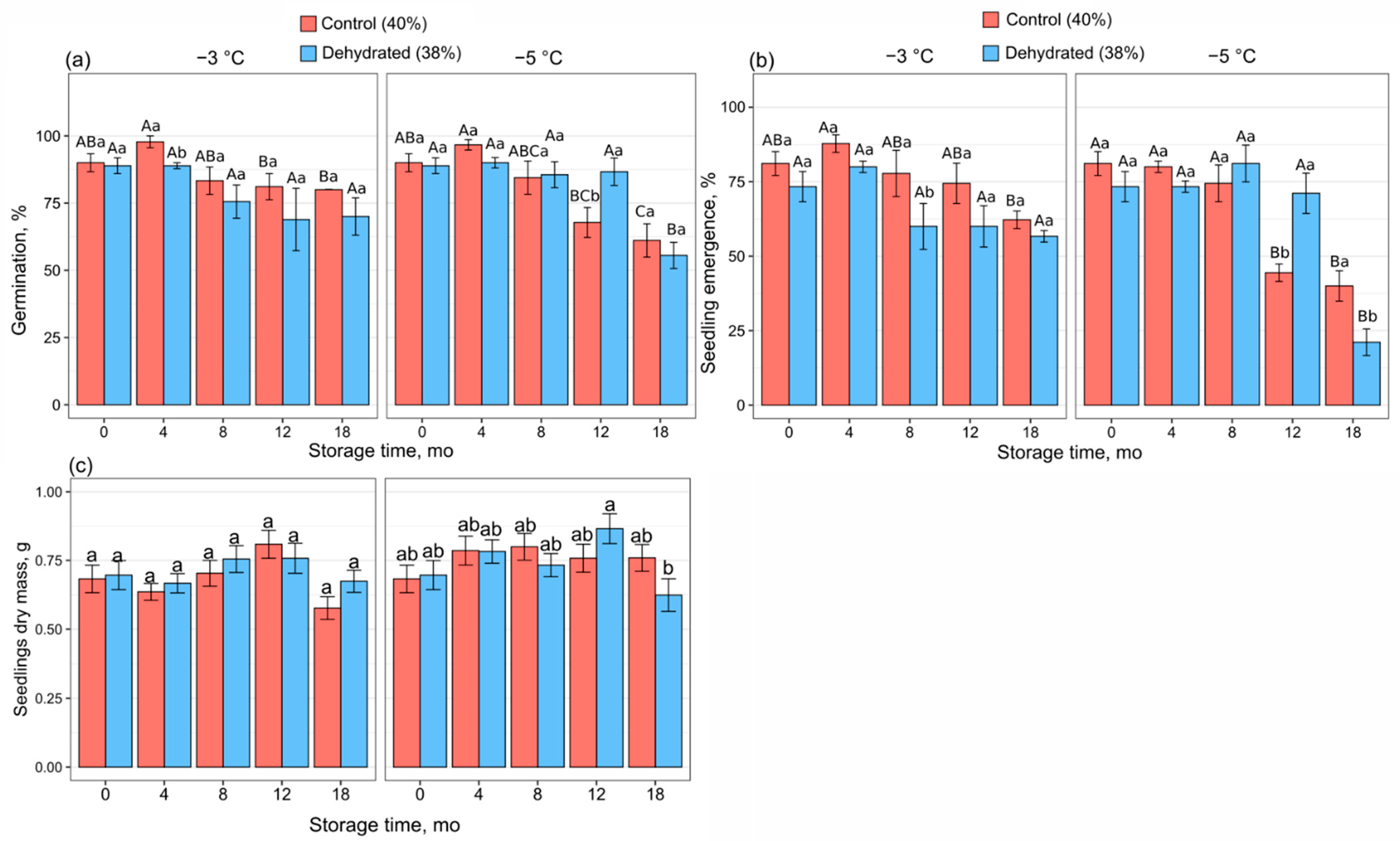
Over the course of the experiment, the germination rate remained above 50%. Despite a decreasing trend, the germination capacity of dehydrated seeds after 18 months of storage at −3 °C did not differ significantly from the control (
Figure 2a). Under the recommended storage conditions (−3 °C, 40% MC), acorns exhibited a significant drop in germination capacity after the 12th and 18th mo of storage. Seed storage at −5 °C revealed a similar decrease in the germination capacity of seeds kept at 40% MC starting from the 8th mo of storage (
Figure 2a). In contrast, the germination capacity of dehydrated seeds stored at −5 °C displayed a significant decrease only after 18 mo of storage. Except for this, the decrease in MC appeared to be non-detrimental in relation to germination capacity under lower storage temperatures.
The pattern of seedling emergence in the course of storage differed between the two analyzed storage temperatures and showed remarkable differences after 18 mo of storage. Seedling emergence also strongly depended on acorn MC when stored at −5 °C. Storage at −3 °C and 40% MC resulted in decreased seedling emergence after the 12th mo (
Figure 2b). Such a decrease was not observed at −3 °C and 38% MC storage conditions because seedling emergence did not statistically differ between the initial values and those reported after 18 mo of storage. Seedling emergence from seeds stored at −5 °C and 40% MC declined strongly after 12 mo of storage, whereas when seeds were stored at −5 °C and 38% MC, a decline appeared 6 mo later. Interestingly, the seedling emergence rate was continuously high for up to one year of storage at −5 °C when seed MC was reduced to 38%. In contrast, the recommended 40% MC combined with storage temperature reduced to −5 °C permitted uniform seedling emergence for only up to 8 mo.
Seedlings raised from stored acorns became significantly smaller after 18 mo of storage at −3 °C, regardless of MC (
Table S1). However, storage at −5 °C resulted in more even seedlings’ DM except for seedling received after 18 mo of storage of dehydrated acorns (
Figure 2c,
Table S1).
During the 18 months of storage, the average dry biomass of whole acorn batches was reduced by 6% to 10.5% (
Figure 3a,
Table S2). The differences in biomass losses among treatments appeared after 8 mo of storage and became particularly pronounced after the 12th mo of storage. The smallest loss in dry biomass occurred at −5 °C and 38% MC, i.e., in the coolest storage regime of the driest seeds. The above calculation comprises both healthy and infected acorns; therefore, the biomass of healthy embryos (cotyledons with embryonic axes) was determined separately and expressed per 100 healthy acorns. In these standardized samples, the decline in biomass per 100 seeds at the final two harvests was also lowest at −5 °C and 38% MC (
Figure 3b). Interestingly, 40% MC clearly resulted in the greatest loss in dry biomass when only healthy seeds were considered.
The concentration of starch in seeds varied between 31 and 26% of dry mass throughout the experiment with no temperature or MC effect and with a significant decline attributable to storage time (
p = 0.036;
Tables S2c and S3). In the samples standardized for acorn number, a loss of starch pool was even more evident, as shown by the strongly significant effect of storage time (
p < 0.001
Figure 4a;
Table S2b). This loss occurred mostly between the 4th and 8th mo of storage and was not related to temperature or MC.
Soluble carbohydrates initially accounted for c. 6% of acorn biomass and its concentration generally increased throughout the experiment, particularly at −5 °C, where they exceeded 9% (temperature effect
p < 0.001;
Tables S2a and S3). A slight overall decline at the final harvest was also noted. The dynamics of soluble carbohydrate pools in standardized samples overall resembled those of concentrations (
Figure 4b,
Table S2). Importantly, the levels of soluble carbohydrates were higher in seeds stored at −5 °C than in seeds stored at −3 °C, and this difference started to be evident after 12 mo of storage. However, given the minor contribution of soluble sugars to TNC, the dynamics of TNC mostly reflected that of starch, with a decline in time and no temperature or moisture effect (
Figure 4c;
Table S2b).
At the start of the experiment, acorns contained slightly over 5% fat. Gradual declines in fat concentration and pool sizes per 100 acorns occurred during storage (
Figure 4d;
Table S3). Despite the fact that the influences of storage temperature or MC were not significant, the decline in fat per 100 healthy acorns after 18 months of storage reached 4.1 g when seeds were stored at recommended conditions (–3 °C, 40% MC) and, in contrast, only 1.6 g when seeds were stored at −5 °C and 38% MC.
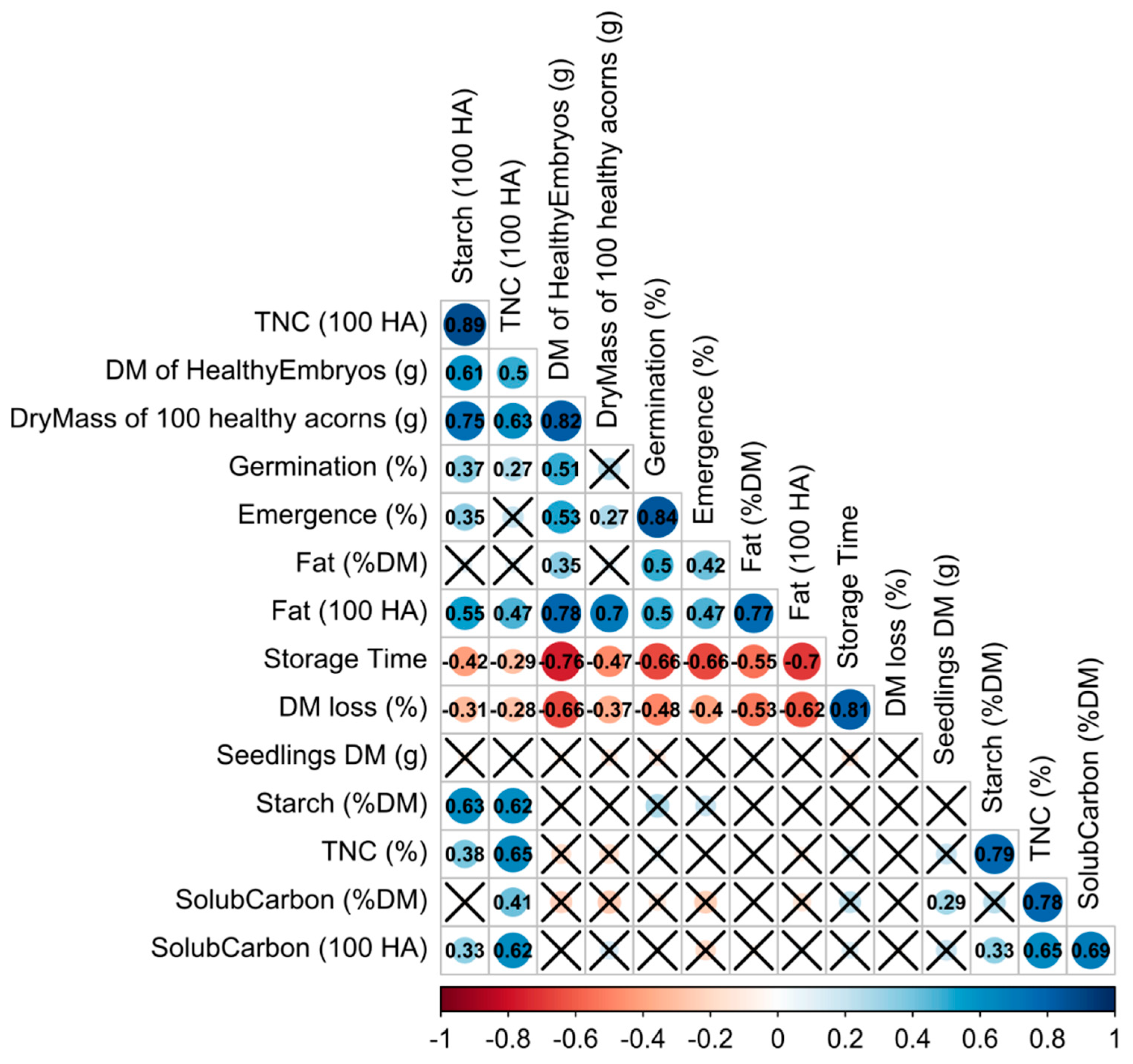
Both germination capacity and seedling emergence were negatively correlated with the loss of DM and storage time. Importantly, germination capacity and seedling emergence were positively correlated with starch and fat. Additionally, seedling emergence was positively correlated with the DM of healthy acorns. Storage temperature was positively correlated with the DM of healthy acorns, whereas a negative correlation occurred with the levels of TNC and soluble carbohydrates (
Figure 5).
4. Discussion
Although optimal short-term storage conditions have previously been established for pedunculate oak acorns, their storage still remains challenging because the seeds lose viability very quickly. Depending on seed quality and provenance, acorns might exhibit the first symptoms of seed deterioration after 6 months of storage under the recommended conditions [
17]. The longest storage of pedunculate oak acorns was recorded by Suszka and Tylkowski [
9]; however, a substantial decline in seed germination and seedling emergence was noted after three years, and after the fifth year of storage, these rates dropped to approximately 12%. In the present study, seeds stored for one year at −5 °C and 38% MC exhibited unaffected germination capacity and seedling emergence compared to the corresponding parameters before storage, which were higher than those in seeds stored at −3 °C and 40% MC (
Figure 2). A decline in seed germination and seedling emergence in seeds from selected provenances stored at −5 °C occurred after 3 months [
15]. Clear differences in acorn storage were noted depending on provenance and seed quality [
33,
34]. Therefore, the results of seed storage studies should be taken with caution, and the effect of provenance should be considered. In our study, a deterioration of seeds was clearly noticed after 18 mo of storage and manifested itself as a substantial decrease in both germination capacity and seedling emergence (
Figure 2), as well as a decline in seedling biomass. Importantly, after 18 mo, the beneficial effect of −5 °C and 38% MC storage conditions on germination and emergence was lost.
In general, seeds of recalcitrant species are recommended to be stored moist at ≥10 °C for less than 1 year [
4], as seeds maintain their metabolism during storage [
25]. As shown in our study, this basic metabolism even at temperatures below 0 °C resulted in a decline in starch and fat reserves, which together constitute approximately one-third of the cotyledon biomass. Steady-state concentrations of components of the tricarboxylic acid (TCA) cycle were not up- or downregulated during 6 mo of pedunculate oak storage [
17], suggesting that aerobic respiration was probably ongoing at a stable rate resulting in continuous reserve depletion. The continuous increase in the hydration state of the pericarp additionally suggests that water was produced from respiration (
Figure 1). Interestingly, the effect of storage temperature on the hydration status of cotyledons was evident (
Figure 1b). The increase in MC in cotyledons of seeds stored at −3 °C was evident, whereas at −5 °C, alterations in MC were not significant. Taken together, lowering the storage temperature to −5 °C increased the limitation of metabolic oxidation of carbohydrates, leading to the smallest loss in acorn biomass (
Figure 3a) and the highest biomass of healthy acorns (
Figure 3b). Loss of biomass was the highest when seeds were stored at −3 °C regardless of their MC.
We previously reported that storage of pedunculate oak seeds at a lower-than-optimal temperature strongly affects starch and sucrose metabolism as well as pentose and glucoronate interconversions pathways. An increased abundance of defense-related metabolites and anti-freezing compounds was also reported after 6 mo of storage. These metabolites included monocarbohydrates and sugar alcohols, many of which can function as osmoprotectants and cryoprotectants [
17]. It may be expected that more osmoprotectants would be needed in seeds displaying lower MC, and similarly, more cryoprotectants should be synthesized at lower storage temperatures. Notably, the combination of −5 °C and 38% MC appeared to be the treatment in which the biomass loss was minimal (
Figure 2), and at the same time, storage at −5 °C resulted in the highest levels of soluble carbohydrates. This indicates that this lower temperature stimulated the synthesis of cryoprotectants and/or osmoprotectants. It is possible that at −5 °C, the acorns experienced and responded to freezing stress, similar to the freezing stress in acorns stored at −7 °C [
17], where the risk of ice crystal formation occurs [
35]. To discriminate whether the increase in soluble carbohydrates was the manifestation of mechanisms related to better protection or to the response to stress, the fact that infected acorns had higher-than-average concentrations of starch (as deduced from the difference in patterns between the raw concentration of starch and its standardized pool size) should be considered. Healthy seeds displayed higher losses of starch and therefore were able to adjust their metabolism, including respiration, responses to stimuli, and limitation of deterioration. However, soluble carbohydrates were not correlated with germination capacity or seedling emergence but positively correlated with seedling DM. It is likely that a lower storage temperature induced the synthesis of soluble carbohydrates as a response to subzero temperatures close to freezing. Both seed germination and seedling establishment were positively correlated with pools of starch, TNC, and fat, but not with soluble carbohydrates.
Damaged and undamaged acorns differ in their metabolism during storage [
17]. The fact that infected acorns catabolized less starch (
Figure 4) supports the hypothesis that fermentation is predominantly present in infected or damaged acorns [
17]. As a result, acorns were metabolically less active and insufficient in aerobic respiration. It is possible that the limited oxygen supply caused by hypoxia accommodated the respiratory/metabolic activity of acorns. More detailed investigation is needed to determine the metabolic regulation that led seeds stored at −5 °C and 38% MC to consume less starch and simultaneously accumulate more soluble sugars than seeds stored at −3 °C and how they were later able to produce high-quality seedlings. Seedlings obtained from acorn stored at −5 °C after 12 and 18 mo were generally larger than those obtained from acorn stored at −3 °C. However, seedling emergence was lower in acorns stored at −5 °C than at −3 °C. Therefore, a possible explanation is that at −5 °C, only the most robust seeds germinated and produced larger seedlings.
The MC of embryonic axes significantly decreased at −5 °C during storage, whereas no change occurred in acorns stored at −3 °C (
Figure 1b). Assuming metabolism was the source of water, this again suggests that whole seeds decelerate their metabolism at −5 °C. This deceleration was most evident in acorns dehydrated to 38%. Anti-freezing compounds, predominantly sugar alcohols, were upregulated, particularly in embryonic axes [
17], which exhibit overall higher MC than cotyledons.
Acorn biomass loss during storage might have also included nonstructural components or proteins—both unaccounted for in this study. Our previous investigations revealed that the N concentration was halved in undamaged cotyledons stored for 6 moat −3 °C and damaged cotyledons of seeds stored at −7 °C compared to the non-stored cotyledons [
17]. Interestingly, the C/N ratio doubled, particularly in undamaged cotyledons [
17], suggesting that protein metabolism is also affected by the time of storage and acorn quality.
Larger seeds, and therefore an abundance of storage materials, have a beneficial effect on growth in seedlings suffering from defoliation, drought, shade, and competition [
36], but not in oaks in which the removal of 1/3 of cotyledons [
24] not only does not disturbs germination, but may even lead to better and synchronized growth in nurseries. It is possible that seed reserves in acorns are redundant for germination, especially under non-stressful conditions in forest nurseries, or that reserve mobilization from distal tissues in the cotyledons is inefficient. The biomass loss observed in our study did not exceed 18% (
Figure 3), and the loss of starch and TNC was of similar magnitude, indicating that acorns in all treatments had sufficient reserves to meet their carbon demand during both dormancy and germination. The overall level of reserves may thus be an indicator of seed vitality rather than a direct cause of differences in vitality between treatments. Rather, qualitative as well as quantitative dynamics of proteins, lipids, and other metabolites induced by the storage conditions likely hold clues to vitality declines during storage.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




