
Promotion Effects of Taxus chinensis var. mairei on Camptotheca acuminata Seedling Growth in Interplanting Mode
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pot Experiment
2.2. C. acuminata Seedlings Bioassay
2.2.1. Determination of Plant Height and Basal Diameter
2.2.2. Determination of Main Photosynthetic Indicators and Chlorophyll Content
2.2.3. Determination of Total Nitrogen and Organic Carbon in Soil
2.2.4. Dynamic Changes in Soil Microbial Population and Enzyme Activities
2.3. Quantitation of Allelochemicals in Rhizosphere and Non-Rhizosphere Soils
2.4. Statistical Analysis
3. Results and Discussion
3.1. Effect of Interplanting on Plant Height and Basal Diameter of C. acuminata Seedlings
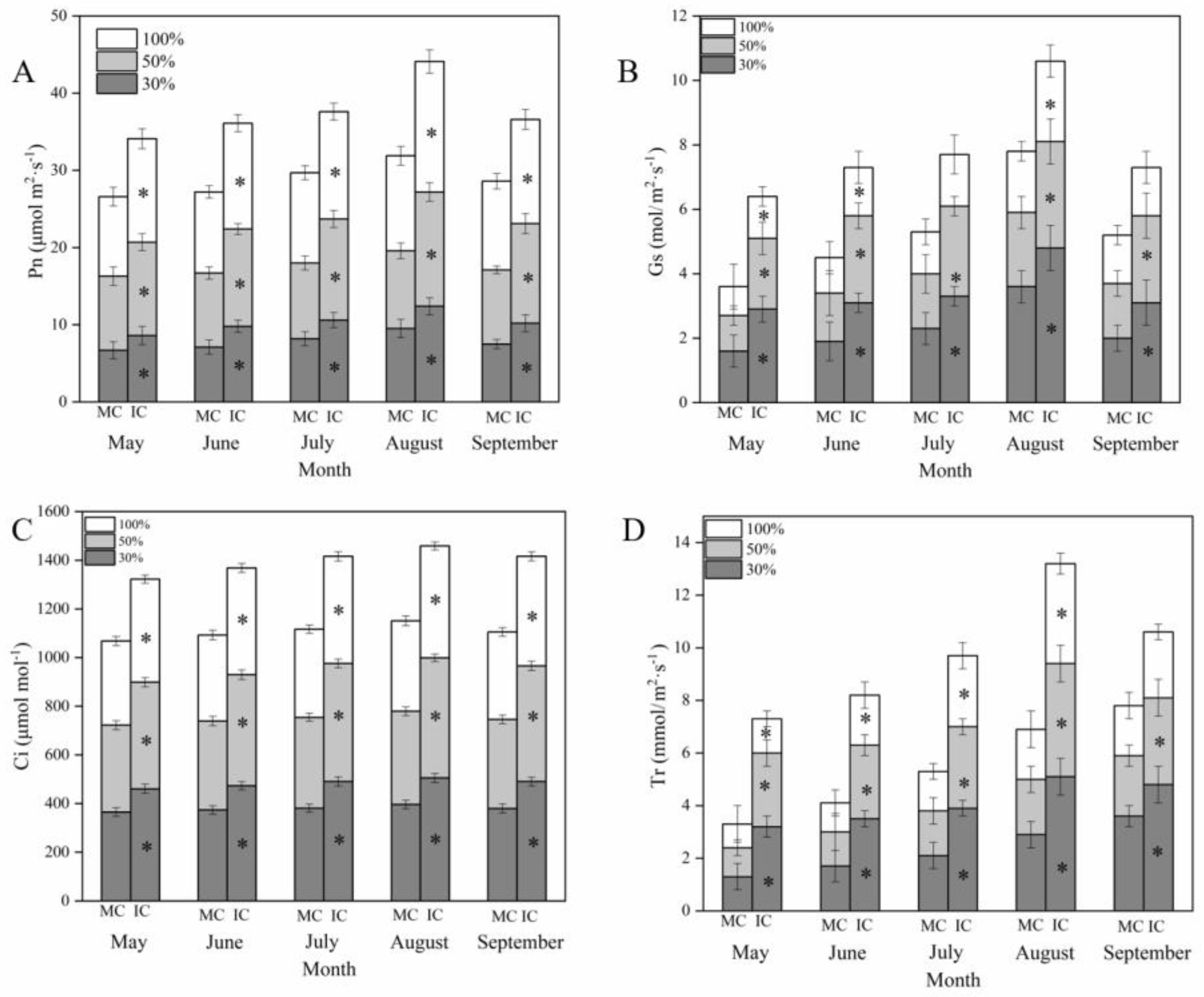
3.2. Effect of Interplanting on Photosynthesis of C. acuminata Seedlings
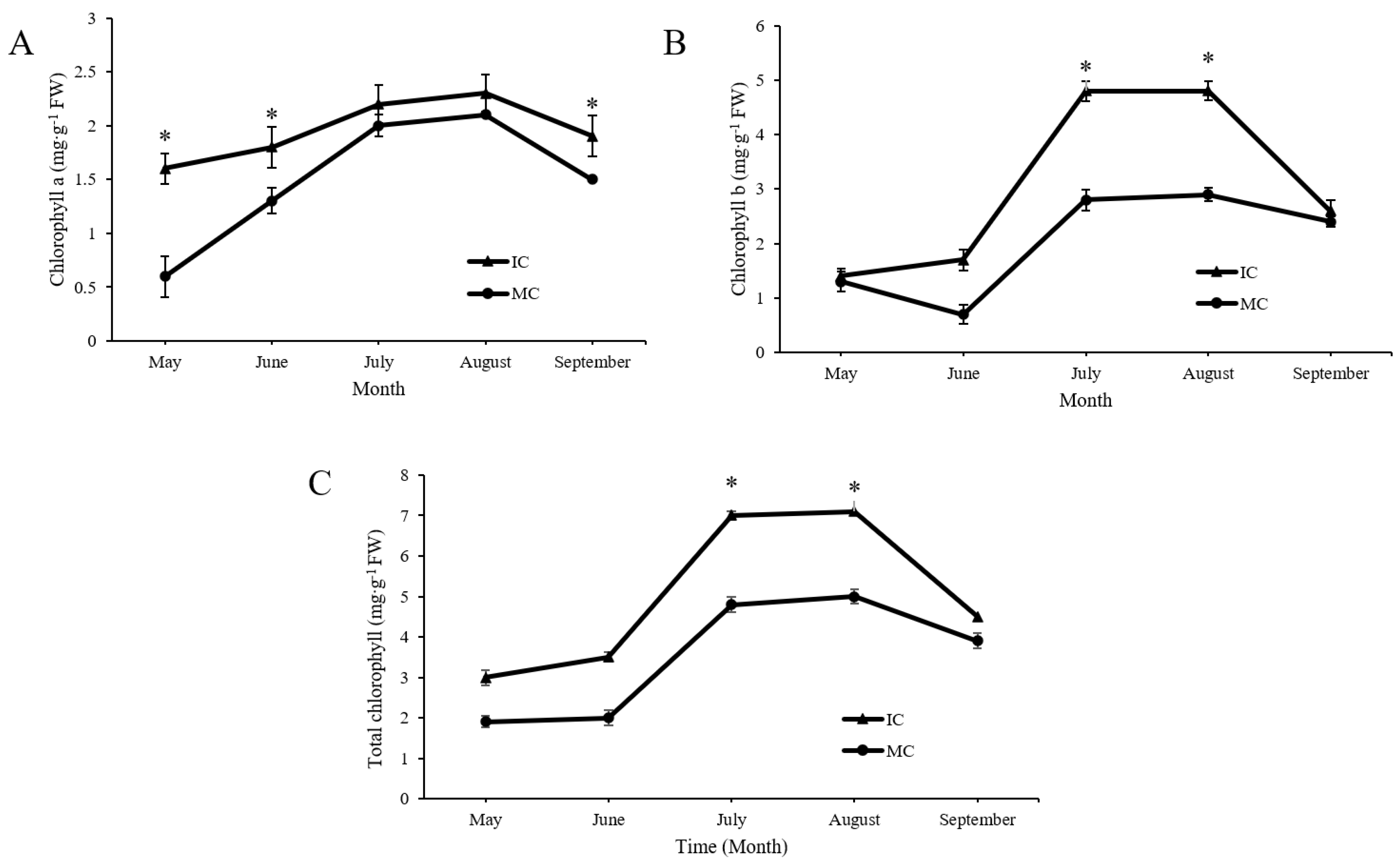
3.3. Effect of Interplanting on Chlorophyll Content of C. acuminata Seedlings
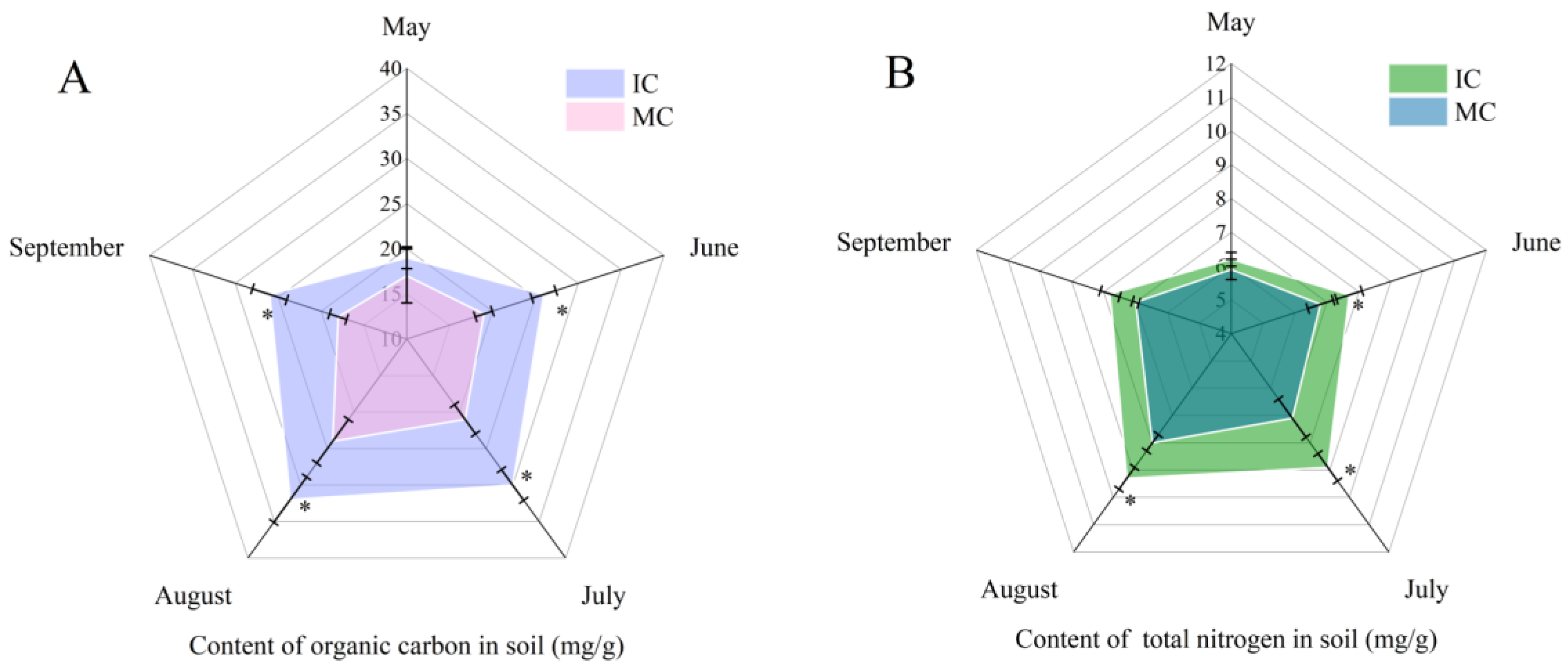
3.4. Effect of Interplanting on Content of Organic Carbon and Total Nitrogen in Soil of C. acuminata Seedlings
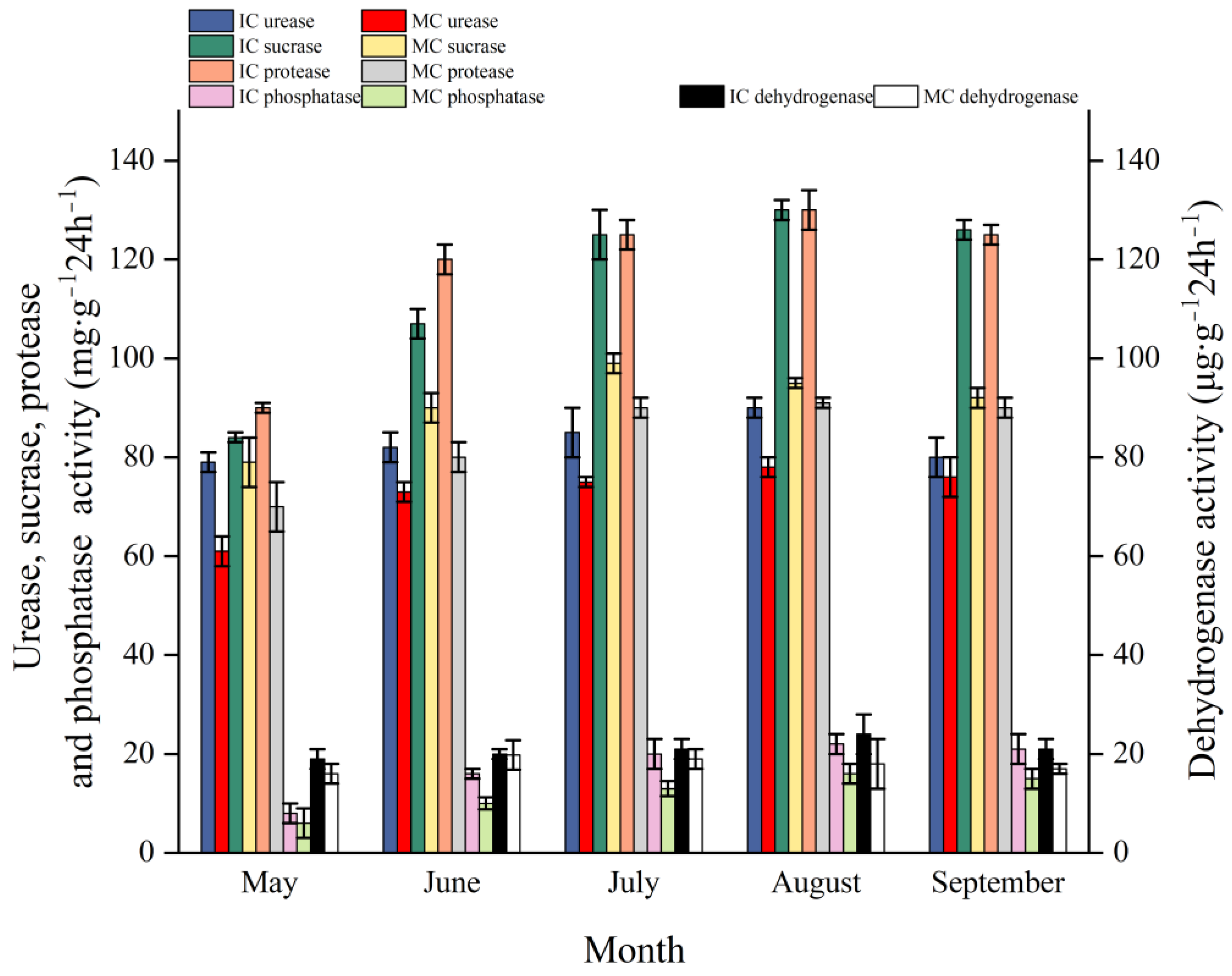
3.5. Effect of Interplanting on Enzyme Activity in the Soil of C. acuminata Seedlings
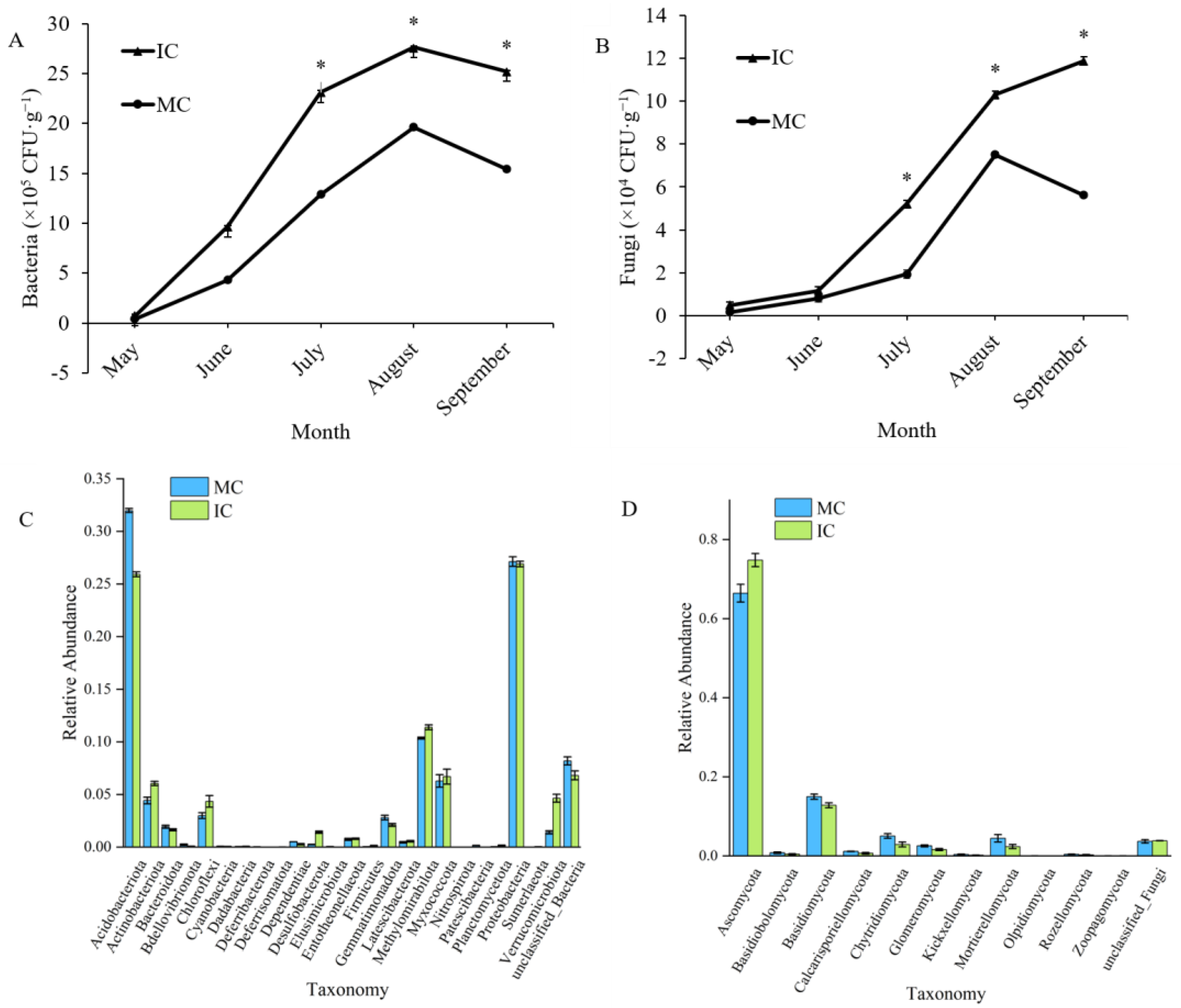
3.6. Effect of Interplanting on Microbial Populations in the Soil of C. acuminata Seedlings
3.7. Release of Taxane Allelochemicals from T. chinensis var. Mairei in Interplanting Mode
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chmura, D.J.; Guzicka, M.; Rokowski, R. Accumulation of standing aboveground biomass carbon in Scots pine and Norway spruce stands affected by genetic variation. For. Ecol. Manag. 2021, 496, 119476. [Google Scholar] [CrossRef]
- Xu, Y.; Cheng, H.F.; Kong, C.H.; Meiners, S.J. Intra-specific kin recognition contributes to inter-specific allelopathy: A case study of allelopathic rice interference with paddy weeds. Plant Cell Environ. 2021, 44, 3479–3491. [Google Scholar] [CrossRef] [PubMed]
- Subrahmaniam, H.J.; Libourel, C.; Journet, E.P.; Morel, J.B.; Munos, S.; Niebel, A.; Raffaele, S.; Roux, F. The genetics underlying natural variation of plant-plant interactions, a beloved but forgotten member of the family of biotic interactions. Plant J. 2018, 93, 747–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmann, S. As above so below: Recent and future advances in plant-mediated above- and belowground interactions. Am. J. Bot. 2022, 109, 672–675. [Google Scholar] [CrossRef]
- Barberán, A.; Mcguire, K.L.; Wolf, J.A.; Jones, F.A.; Wright, S.J.; Turner, B.L.; Essene, A.; Hubbell, S.P.; Faircloth, B.C.; Fierer, N. Relating belowground microbial composition to the taxonomic, phylogenetic, and functional trait distributions of trees in a tropical forest. Ecol. Lett. 2015, 18, 1397–1405. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.R.; Huang, Y.T.; Fu, S.L.; Wang, J.X.; Ming, A.G.; Li, X.Z.; Yao, M.J.; Li, H. Tree species mixture inhibits soil organic carbon mineralization accompanied by decreased r-selected bacteria. Plant Soil 2018, 431, 203–216. [Google Scholar] [CrossRef]
- Shi, S.; Cheng, J.B.; Ahmad, N.; Zhao, W.Y.; Tian, M.F.; Yuan, Z.Y.; Li, C.Y.; Zhao, C.J. Effects of potential allelochemicals in a water extract of Abutilon theophrasti Medik. on germination and growth of Glycine max L., Triticum aestivum L., and Zea mays L. J. Sci. Food Agric. 2022. [Google Scholar] [CrossRef]
- Reynolds, H.L.; Packer, A.; Bever, J.D.; Clay, K. Grassroots ecology: Plant-microbe-soil interactions as drivers of plant community structure and dynamics. Ecology 2003, 84, 2281–2291. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Huang, B.; Du, N.; Guo, S.; Shu, S.; Sun, J. Effects of garlic/cucumber relay intercropping on soil enzyme activities and the microbial environment in continuous cropping. Hortscience 2017, 52, 78–84. [Google Scholar] [CrossRef]
- Rehman, S.; Shahzad, B.; Bajwa, A.A.; Hussain, S.; Rehman, A.; Cheema, S.A.; Abbas, T.; Ali, A.; Shah, L.; Adkins, S.; et al. Utilizing the allelopathic potential of brassica species for sustainable crop production: A review. J. Plant Growth Regul. 2019, 38, 343–356. [Google Scholar] [CrossRef]
- Li, S.; He, H.; Xi, Y.; Li, L. Chemical constituents and pharmacological effects of the fruits of Camptotheca acuminata: A review of its phytochemistry. Asian J. Tradit. Med. 2018, 13, 40–48. [Google Scholar]
- Krishnan, J.J.; Gangaprasad, A.; Satheeshkumar, K. In vitro mass multiplication and estimation of camptothecin (CPT) in Ophiorrhiza mungos L. var. angustifolia (Thw.) Hook. f. Ind. Crops Prod. 2018, 119, 64–72. [Google Scholar] [CrossRef]
- Zhao, C.; Li, C.; Wang, L.; Zu, Y.; Yang, L. Determination of camptothecin and 10-hydroxycamptothecin in Camptotheca acuminata by LC-ESI-MS/MS. Anal. Lett. 2010, 43, 2681–2693. [Google Scholar] [CrossRef]
- Sadre, R.; Magallanes-Lundback, M.; Pradhan, S.; Salim, V.; Mesberg, A.; Jones, A.D.; Dellapenna, D. Metabolite diversity in alkaloid biosynthesis: A multilane (diastereomer) highway for camptothecin synthesis in Camptotheca acuminata. Plant Cell 2016, 28, 1926–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, B.; Yang, P. Implications of seed germination ecology for conservation of Camptotheca acuminata, a rare, endemic, and endangered species in China. Plant Ecol. 2021, 222, 209–219. [Google Scholar] [CrossRef]
- Trueman, S.J.; Richardson, D.M. Propagation and chlorophyll fluorescence of Camptotheca acuminata cuttings. J. Med. Plants Res. 2011, 5, 1–6. [Google Scholar]
- Li, L.; Yang, F.; Pang, H.; Gao, Y.; Sun, J. Relationship between dynamic change of growth and meteorological factors in artificial composite community of Camptotheca acuminata. Chin. J. Plant Res. 2008, 28, 486–490. [Google Scholar]
- Wang, J. Studies on Directional Cultivation of Active Substances for Artificial Complex Community of Camptotheca chinensis and Rosemary; Northeast Forestry University: Harbin, China, 2006. [Google Scholar]
- Li, L.; Chen, Y.; Ma, Y.; Wang, Z.; Wang, T.; Xie, Y. Optimization of Taxol extraction process using response surface methodology and investigation of temporal and spatial distribution of taxol in Taxus mairei. Molecules 2021, 26, 5485. [Google Scholar] [CrossRef]
- Yang, F.; Liu, W.; Wang, W.; Zu, Y. Characteristics of life history type spectrum of mixed communities of Camptotheca acuminata and Taxus chinensis. Bull. Bot. Res. 2011, 31, 711–715. [Google Scholar]
- Shi, S.; Gao, Y.X.; Li, C.Y.; Zhang, J.J.; Guan, J.J.; Zhao, C.J.; Fu, Y.J. Allelopathy of Taxus chinensis var. mairei on Camptotheca acuminata seedling growth and identification of the active principles. J. Plant Interact. 2022, 17, 33–42. [Google Scholar] [CrossRef]
- Lee, T.-C.; Shih, T.-H.; Huang, M.-Y.; Lin, K.-H.; Huang, W.-D.; Yang, C.-M. Eliminating interference by anthocyanins when determining the porphyrin ratio of red plant leaves. J. Photochem. Photobiol. B-Biol. 2018, 187, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Martz, M.; Heil, J.; Marschner, B.; Stumpe, B. Effects of soil organic carbon (SOC) content and accessibility in subsoils on the sorption processes of the model pollutants nonylphenol (4-n-NP) and perfluorooctanoic acid (PFOA). Sci. Total Environ. 2019, 672, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.K.D.; Panda, R.K.P.; Chakraborty, A.P. Quantification of nitrogen transformation and leaching response to agronomic management for maize crop under rainfed and irrigated condition. Environ. Pollut. 2020, 265, 114866. [Google Scholar] [CrossRef] [PubMed]
- Borowik, A.; Wyszkowska, J.; Kucharski, J.; Baćmaga, M.; Tomkiel, M. Response of microorganisms and enzymes to soil contamination with a mixture of terbuthylazine, mesotrione, and S-metolachlor. Environ. Sci. Pollut. Res. 2017, 24, 1910–1925. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, F.; Wu, S.; Guo, Z.; Huang, X.; Hu, X.; Holmes, M.; Zou, X. Noise-free microbial colony counting method based on hyperspectral features of agar plates. Food Chem. 2019, 274, 925–932. [Google Scholar] [CrossRef]
- Spohn, M.; Klaus, K.; Wanek, W.; Richter, A. Microbial carbon use efficiency and biomass turnover times depending on soil depth—Implications for carbon cycling. Soil Biol. Biochem. 2016, 96, 74–81. [Google Scholar] [CrossRef]
- Castano, C.; Berlin, A.; Durling, M.B.; Ihrmark, K.; Lindahl, B.D.; Stenlid, J.; Clemmensen, K.E.; Olson, A. Optimized metabarcoding with Pacific biosciences enables semi-quantitative analysis of fungal communities. New Phytol. 2020, 228, 1149–1158. [Google Scholar] [CrossRef]
- Gromes, R.; Mueller, K.; Nagel, E.; Uhlmann, J. The use of photometric test kits for investigations of soil enzyme activities. J. Plant Nutr. Soil Sci. 2015, 164, 431–433. [Google Scholar] [CrossRef]
- Bin, Q.; Siming, N.; Qianqian, L.; Zahid, M.; Jiabo, C.; Zhanyu, Y.; Chunying, L.; Chunjian, Z. Quick and in-situ detection of different polar allelochemicals in Taxus soil by microdialysis combined with UPLC-MS/MS. J. Agric. Food Chem. 2022. [Google Scholar] [CrossRef]
- Kong, C. Allelopathic potential of root exudates of larch (Larix gmelini) on Manchurian walnut (Juglans mandshurica). Allelopath. J. 2007, 20, 127–134. [Google Scholar]
- Wang, X.; Zhang, R.; Wang, J.; Di, L.; Sikdar, A. The effects of leaf extracts of four tree species on Amygdalus pedunculata seedlings growth. Front. Plant Sci. 2021, 11, 587579. [Google Scholar] [CrossRef]
- Walter, J.; Kromdijk, J. Here comes the sun: How optimization of photosynthetic light reactions can boost crop yields. J. Integr. Plant Biol. 2022, 64, 564–591. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Tang, Y.; Wang, Z.; Gou, Y.; Wang, J. Nitrogen-cycling genes and rhizosphere microbial community with reduced nitrogen application in maize/soybean strip intercropping. Nutr. Cycl. Agroecosystems 2019, 113, 35–49. [Google Scholar] [CrossRef]
- Beneragama, C.; Goto, K. Chlorophyll a:b ratio increases under low-light in ‘Shade-tolerant’ Euglena gracilis. Trop. Agric. Res. 2010, 22, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Zhang, J.; Wen, R.; Chen, T.; Xia, N.; Liu, Y.; Wang, Z. Genetically modified sugarcane intercropping soybean impact on rhizosphere bacterial communities and co-occurrence patterns. Front. Microbiol. 2021, 12, 742341. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Han, Y.; Xu, Z.; Wang, S.; Jiang, M.; Wang, G. Hummock-hollow microtopography affects soil enzyme activity by creating environmental heterogeneity in the sedge-dominated peatlands of the Changbai Mountains, China. Ecol. Indic. 2021, 121, 107187. [Google Scholar] [CrossRef]
- Yu, P.; Tang, X.; Zhang, A.; Fan, G.; Liu, S. Responses of soil specific enzyme activities to short-term land use conversions in a salt-affected region, northeastern China. Sci. Total Environ. 2019, 687, 939–945. [Google Scholar] [CrossRef]
- Zeng, J.; Liu, J.; Lu, C.; Ou, X.; Luo, K.; Li, C.; He, M.; Zhang, H.; Yan, H. Intercropping with turmeric or ginger reduce the continuous cropping obstacles that affect Pogostemon cablin (Patchouli). Front. Microbiol. 2020, 11, 579719. [Google Scholar] [CrossRef]
- Zhao, Q.L.; Song, X.L.; Sun, X.Z. Studies on soil microorganism quantities and soil enzyme activities in the garlic-cotton and wheat-cotton intercropping systems. Plant Nutr. Fertil. Sci. 2011, 17, 1474–1480. [Google Scholar]
- Liu, T.; Cheng, Z.; Meng, H.; Ahmad, I.; Zhao, H. Growth, yield and quality of spring tomato and physicochemical properties of medium in a tomato/garlic intercropping system under plastic tunnel organic medium cultivation. Sci. Hortic. 2014, 170, 159–168. [Google Scholar] [CrossRef]
- Gong, H.; Du, Q.; Xie, S.; Hu, W.; Deng, J. Soil microbial DNA concentration is a powerful indicator for estimating soil microbial biomass C and N across arid and semi-arid regions in northern China. Appl. Soil Ecol. 2021, 160, 1–8. [Google Scholar] [CrossRef]
- Ghani, M.I.; Ali, A.; Atif, M.J.; Ali, M.; Amin, B.; Anees, M.; Khurshid, H.; Cheng, Z. Changes in the soil microbiome in eggplant monoculture revealed by high-throughput Illumina MiSeq Sequencing as influenced by raw garlic stalk amendment. Int. J. Mol. Sci. 2019, 20, 2125–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ma, W.; He, H.; Wang, Z.; Cao, Y. Effects of Sugarcane and soybean intercropping on the nitrogen-fixing bacterial community in the Rhizosphere. Front. Microbiol. 2021, 12, 3349–3353. [Google Scholar] [CrossRef] [PubMed]
- Elshahawy, I.E.; Osman, S.A.; Abd-El-Kareem, F. Protective effects of silicon and silicate salts against white rot disease of onion and garlic, caused by Stromatinia cepivora. J. Plant Pathol. 2020, 103, 27–43. [Google Scholar] [CrossRef]
- Boparai, A.K.; Manchanda, J.S. Response of cotton and wheat cultivars to soil-applied boron in a boron-deficient, noncalcareous typic ustochrept. Commun. Soil Sci. Plant Anal. 2019, 50, 108–118. [Google Scholar] [CrossRef]
- Tian, X.; Li, C.; Zhang, M.; Li, T.; Lu, Y.; Liu, L. Controlled release urea improved crop yields and mitigated nitrate leaching under cotton-garlic intercropping system in a 4-year field trial. Soil Tillage Res. 2018, 175, 158–167. [Google Scholar] [CrossRef]
- Ali, A.; Ghani, M.I.; Ding, H.; Fan, Y.; Cheng, Z.; Iqbal, M. Co-amended synergistic interactions between arbuscular mycorrhizal fungi and the organic substrate-induced cucumber yield and fruit quality associated with the regulation of the AM-fungal community structure under anthropogenic cultivated soil. Int. J. Mol. Sci. 2019, 20, 1539. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Chen, L.; Zhang, S.; Miao, Y.; Zhang, Y.; Li, Z.; Zhao, J.; Yu, L.; Zhang, J.; Qin, X.; et al. Plant interaction patterns shape the soil microbial community and nutrient cycling in different intercropping scenarios of aromatic plant species. Front. Microbiol. 2022, 13, 888789. [Google Scholar] [CrossRef]
- Burns, K.N.; Bokulich, N.A.; Cantu, D.; Greenhut, R.F.; Kluepfel, D.A.; O’Geen, A.T.; Strauss, S.L.; Steenwerth, K.L. Vineyard soil bacterial diversity and composition revealed by 16S rRNA genes: Differentiation by vineyard management. Soil Biol. Biochem. 2016, 103, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Li, S.; Li, R.; Zhang, J.; Liu, Y.; Lv, L.; Zhu, H.; Wu, W.; Li, W. Plant cultivars imprint the rhizosphere bacterial community composition and association networks. Soil Biol. Biochem. 2017, 109, 145–155. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, U.B.; Sahu, P.K.; Paul, S.; Kumar, A.; Malviya, D.; Singh, S.; Kuppusamy, P.; Singh, P.; Paul, D.; et al. Linking soil microbial diversity to modern agriculture practices: A review. Int. J. Environ. Res. Public Health 2022, 19, 3141. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, C.; Wang, X.; Ma, X.; Jiang, L.; Zhu, J.; Gao, J.; Song, C. Microbial abundance as an indicator of soil carbon and nitrogen nutrient in permafrost peatlands. Ecol. Indic. 2020, 115, 106362. [Google Scholar] [CrossRef]
- Chen, K.J.; Zheng, Y.Q.; Kong, C.H.; Zhang, S.Z.; Li, J.; Liu, X.G. 2,4-Dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DIMBOA) and 6-Methoxy-benzoxazolin-2-one (MBOA) levels in the wheat rhizosphere and their effect on the soil microbial community structure. J. Agric. Food Chem. 2010, 58, 12710–12716. [Google Scholar] [CrossRef] [PubMed]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Thompson, A.R.; Roth-Monzón, A.J.; Aanderud, Z.T.; Adams, B.J. Phagotrophic protists and their associates: Evidence for preferential grazing in an abiotically driven soil ecosystem. Microorganisms 2021, 9, 1555. [Google Scholar] [CrossRef]
- Qi, L.; Zhou, P.; Yang, L.; Gao, M. Effects of land reclamation on the physical, chemical, and microbial quantity and enzyme activity properties of degraded agricultural soils. J. Soil Sediments 2020, 20, 973–981. [Google Scholar] [CrossRef]
- Wu, C.; Chen, Y.; GrenierHéon, D. Effects of allelopathy and competition for nutrients and water on the survival and growth of plant species in a natural Dacrydium forest. Can. J. For. Res. 2021, 52, 100–108. [Google Scholar] [CrossRef]
- Kong, C.H.; Xuan, T.D.; Khanh, T.D.; Tran, H.D.; Trung, N.T. Allelochemicals and signaling chemicals in plants. Molecules 2019, 24, 2737–2746. [Google Scholar] [CrossRef] [Green Version]
- Vicua, R.; González, B. The microbial world in a changing environment. Rev. Chil. de Hist. Nat. 2021, 94, 2–5. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Planting Mode | Plant Height (cm) | |||||
|---|---|---|---|---|---|---|
| April | May | June | July | August | September | |
| MC | 50.1 ± 1.2 | 55.9 ± 2.3 | 73.9 ± 2.6 | 92.1 ± 2.4 | 95.7 ± 2.4 | 97.9 ± 2.5 |
| IC | 49.8 ± 1.1 | 56.7 ± 2.1 | 80.9 ± 2.3 * | 105.8 ± 2.6 * | 108.3 ± 2.6 * | 109.8 ± 2.7 * |
| Planting Mode | Basal Diameter (mm) | |||||
|---|---|---|---|---|---|---|
| April | May | June | July | August | September | |
| MC | 7.04 ± 0.04 | 7.23 ± 0.32 | 8.73 ± 0.44 | 9.04 ± 0.36 | 10.12 ± 0.32 | 10.33 ± 0.44 |
| IC | 7.02 ± 0.06 | 8.09 ± 0.34 * | 9.23 ± 0.42 * | 10.84 ± 0.42 * | 11.36 ± 0.38 * | 11.65 ± 0.56 * |
| Compounds (μg/g) | April | May | June | July | August | September | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rhizosphere Soil | Non-Rhizosphere Soil | Rhizosphere Soil | Non-Rhizosphere Soil | Rhizosphere Soil | Non-Rhizosphere Soil | Rhizosphere Soil | Non-Rhizosphere Soil | Rhizosphere Soil | Non-Rhizosphere Soil | Rhizosphere Soil | Non-Rhizosphere Soil | |
| Cephalomannine | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 |
| 10-DAB III | 0.23 ± 0.01 | 0.09 ± 0.01 | 0.34 ± 0.01 | 0.12 ± 0.01 | 0.46 ± 0.01 | 0.15 ± 0.01 | 0.65 ± 0.01 | 0.20 ± 0.01 | 0.67 ± 0.01 | 0.21 ± 0.01 | 0.63 ± 0.01 | 0.19 ± 0.01 |
| Paclitaxel | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 | <0.01 ± 0.01 |
| 7-Epi-10-DAT | 0.13 ± 0.01 | 0.04 ± 0.01 | 0.17 ± 0.01 | 0.06 ± 0.01 | 0.22 ± 0.01 | 0.09 ± 0.01 | 0.30 ± 0.01 | 0.11 ± 0.01 | 0.32 ± 0.01 | 0.12 ± 0.01 | 0.31 ± 0.01 | 0.11 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, C.; Shi, S.; Ahmad, N.; Gao, Y.; Xu, C.; Guan, J.; Fu, X.; Li, C. Promotion Effects of Taxus chinensis var. mairei on Camptotheca acuminata Seedling Growth in Interplanting Mode. Forests 2022, 13, 2119. https://doi.org/10.3390/f13122119
Zhao C, Shi S, Ahmad N, Gao Y, Xu C, Guan J, Fu X, Li C. Promotion Effects of Taxus chinensis var. mairei on Camptotheca acuminata Seedling Growth in Interplanting Mode. Forests. 2022; 13(12):2119. https://doi.org/10.3390/f13122119
Chicago/Turabian StyleZhao, Chunjian, Sen Shi, Naveed Ahmad, Yinxiang Gao, Chunguo Xu, Jiajing Guan, Xiaodong Fu, and Chunying Li. 2022. "Promotion Effects of Taxus chinensis var. mairei on Camptotheca acuminata Seedling Growth in Interplanting Mode" Forests 13, no. 12: 2119. https://doi.org/10.3390/f13122119