
Effects of Climate Change and Environmental Factors on Bamboo (Ferrocalamus strictus), a PSESP Unique to China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Distribution of F. strictus
2.2. Environment Variables
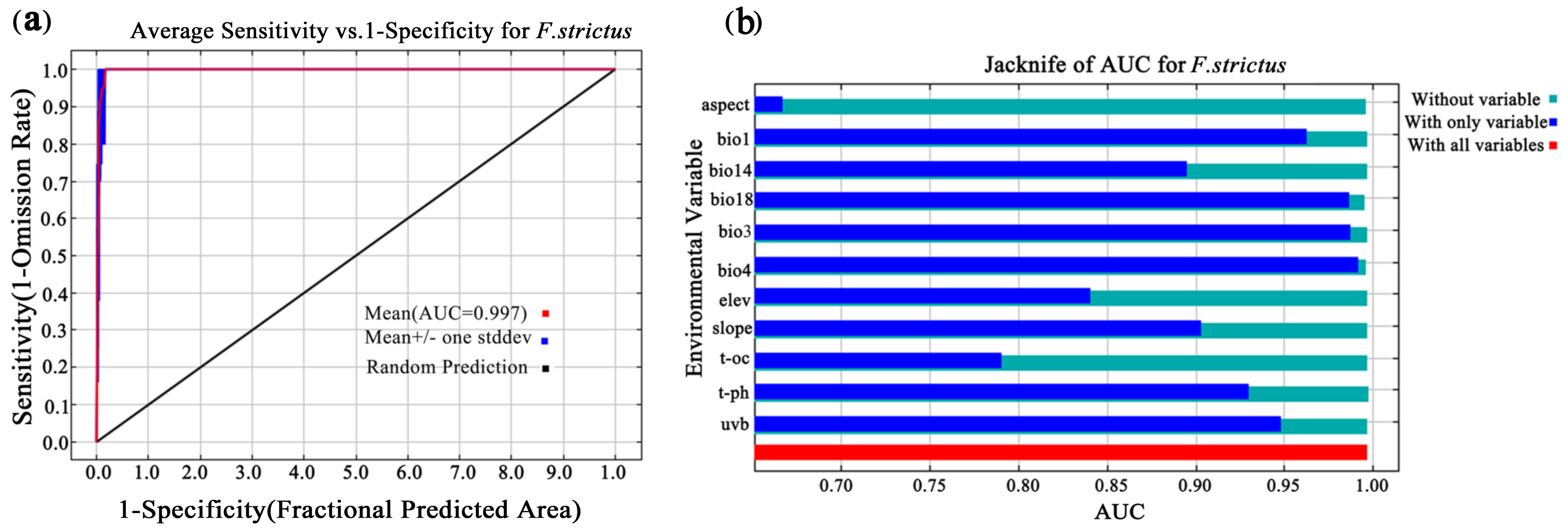
2.3. MaxEnt Model Accuracy
3. Results
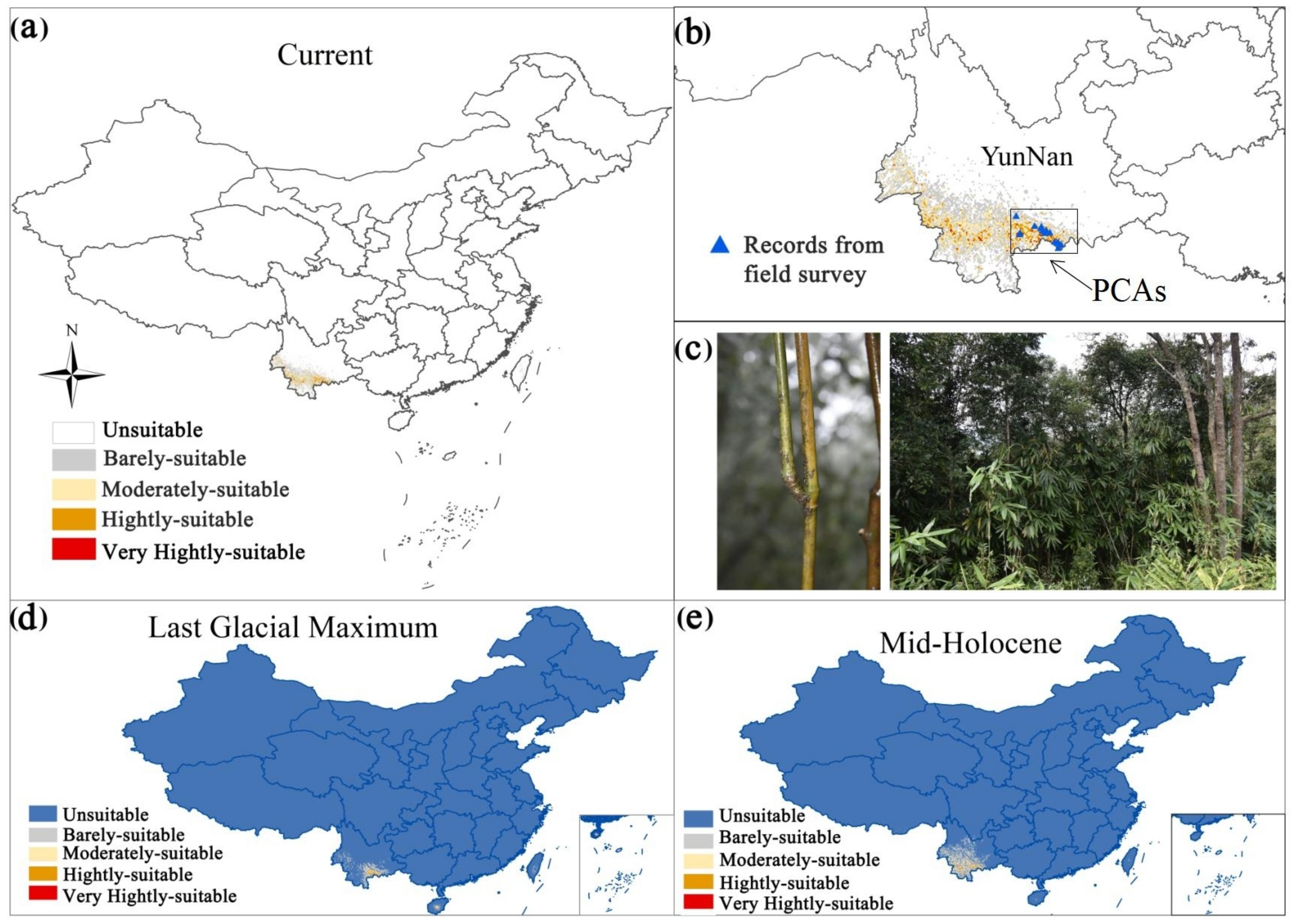
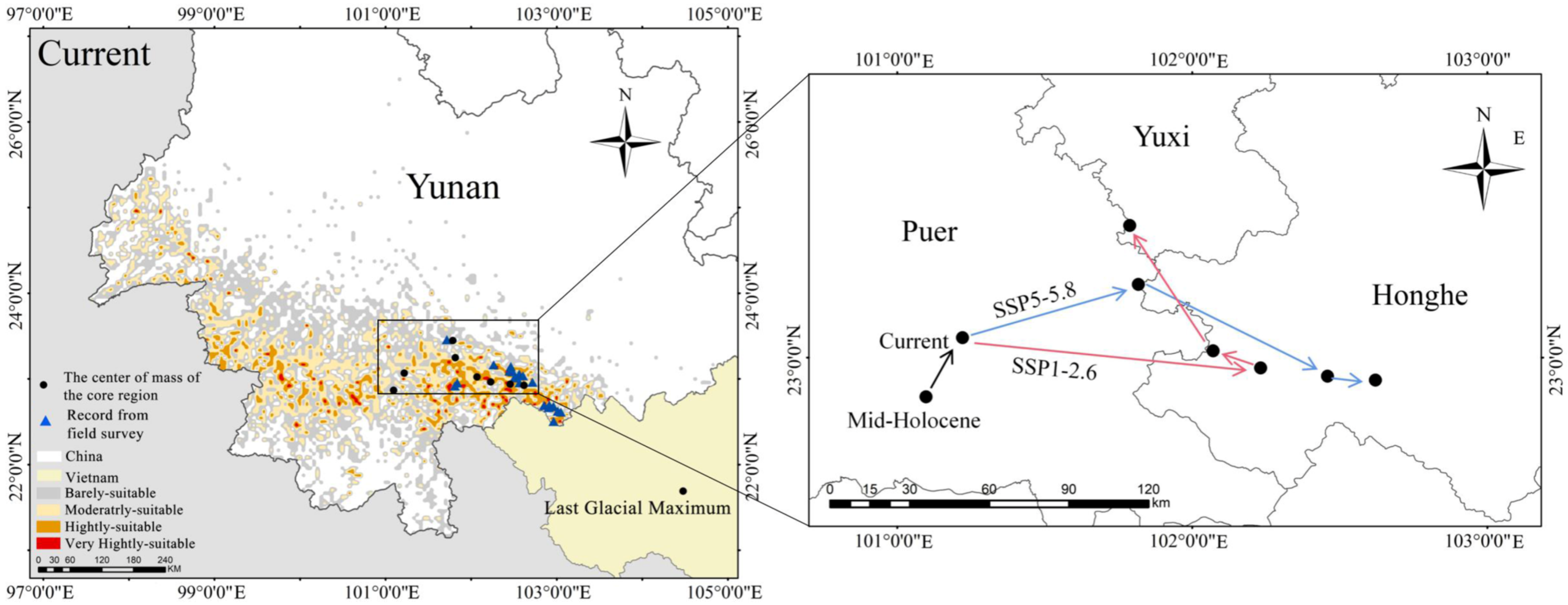
3.1. Population Status of F. strictus
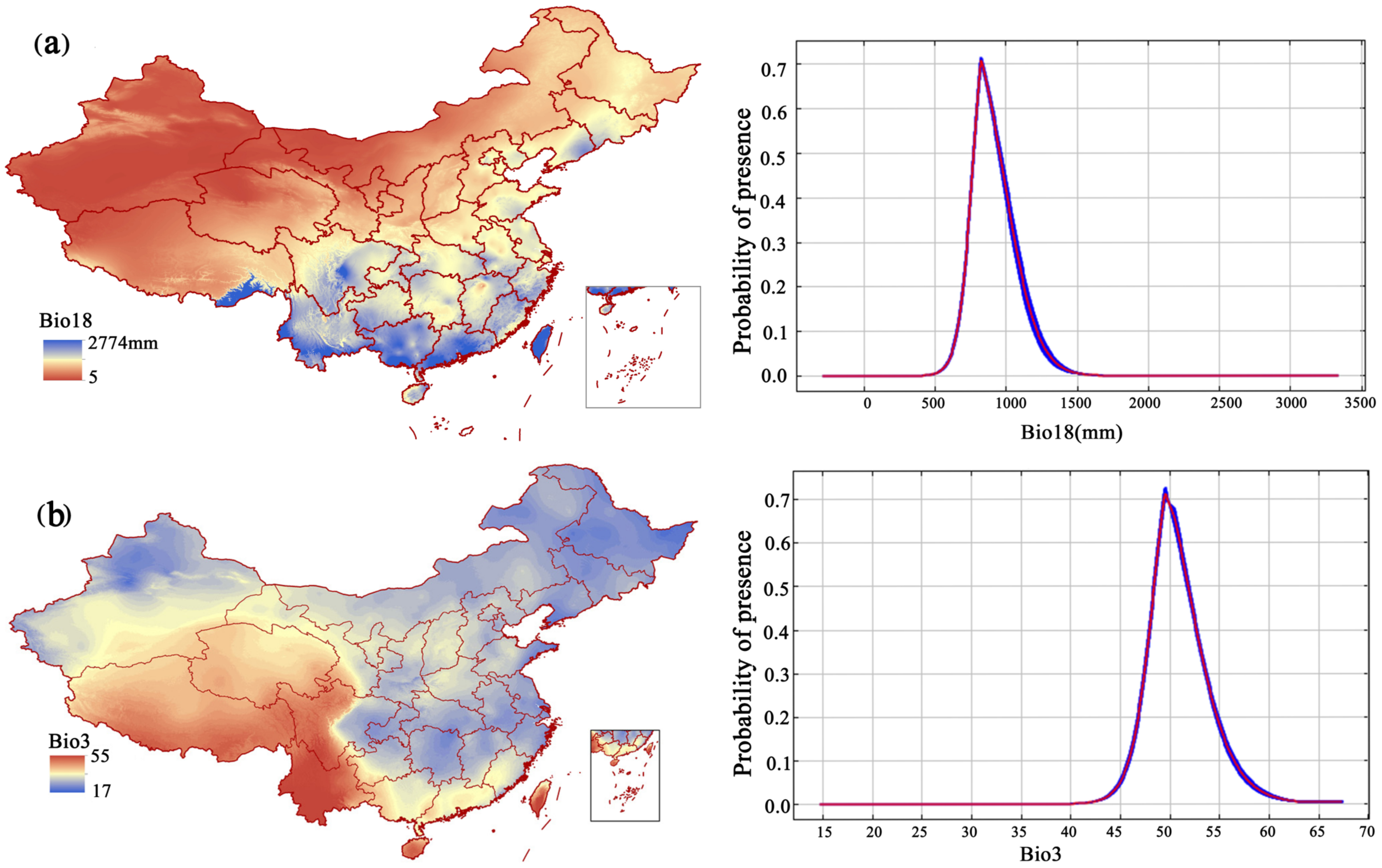
3.2. The Restriction of Environmental Factors on the Geographical Distribution of F. strictus
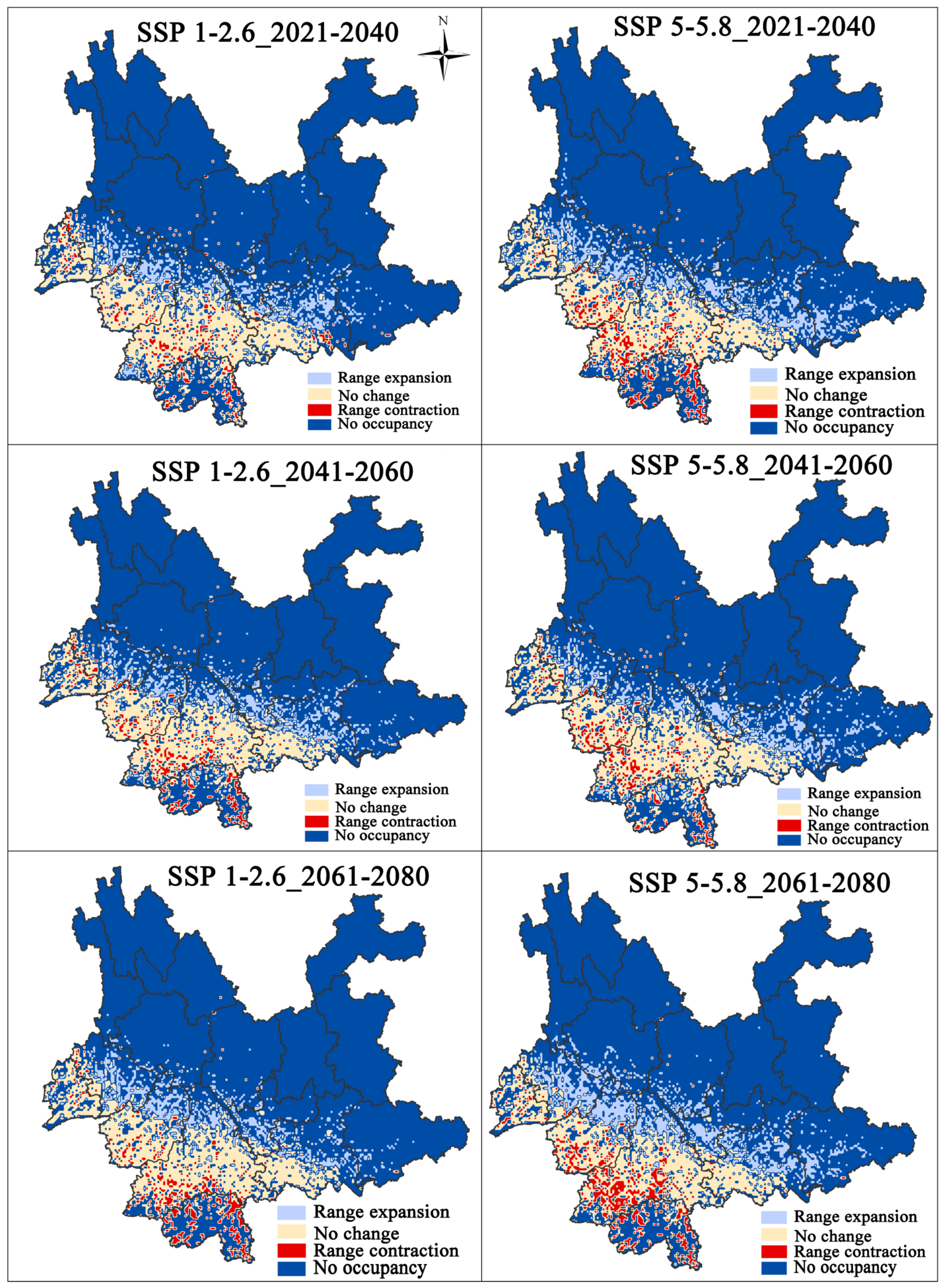
3.3. Changes in the Potential Distribution of F. strictus under Climate Change
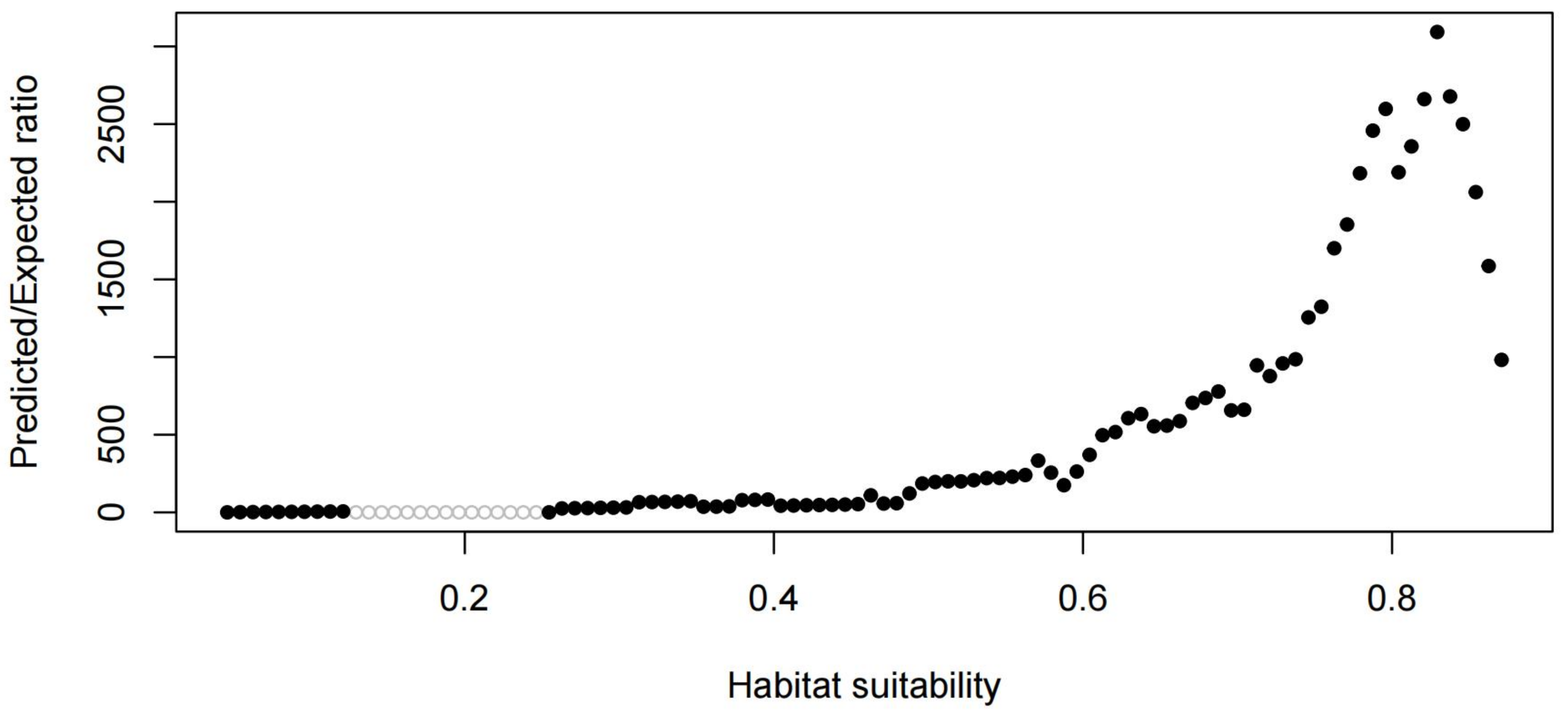
3.4. Priority Conservation Areas
4. Discussion and Conclusions
4.1. The Effect of Climate on Distribution
4.2. Refuge
4.3. Limitations and Prospects
4.4. Recommendations for Protection
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Q.E.; Zhu, G.H.; Hong, D.; Wu, Z.; Raven, P.H. World’s largest flora completed. Science 2005, 309, 2163. [Google Scholar] [CrossRef] [PubMed]
- Volis, S. How to conserve threatened Chinese plant species with extremely small populations? Plant Divers. 2016, 38, 45–52. [Google Scholar] [PubMed] [Green Version]
- Ma, Y.; Gao, C.; Grumbine, R.E.; Dao, Z.; Sun, W.; Guo, H. Conserving plant species with extremely small populations (PSESP) in China. Biodivers. Conserv. 2013, 22, 803–809. [Google Scholar] [CrossRef]
- Ren, H.; Zhang, Q.m.; Lu, H.f.; Liu, H.x.; Guo, Q.F.; Wang, J.; Jian, S.g.; Bao, H. Wild plant species with extremely small populations require conservation and reintroduction in China. Ambio 2012, 41, 8. [Google Scholar]
- State Forestry Administration of China. The Saving and Conservation Program on Extremely Small Populations in China; State Forestry Administration of China: Beijing, China, 2012.
- Sun, W.B.; Yang, J.; Dao, Z.L. Study and Conservation of Plant Species with Extremely Small Populations (PSESP) in Yunnan Province, China; Science Press: Beijing, China, 2019. [Google Scholar]
- Wade, E.M.; Nadarajan, J.; Yang, X.; Ballesteros, D.; Sun, W.; Pritchard, H.W. Plant species with extremely small populations (psesp) in China: A seed and spore biology perspective. Plant Divers. Engl. Ed. 2016, 38, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crane, P. Conserving our global botanical heritage: The PSESP plant conservation program. Plant Divers. 2020, 42, 319–322. [Google Scholar]
- Sun, W.B.; Ma, Y.P.; Blackmore, S. How a new conservation action concept has accelerated plant conservation in china. Trends Plant Sci. 2019, 24, 4–6. [Google Scholar]
- Yang, J.; Muhammad, W.H.; Tao, L.; Zhang, R.; Yun, Q.; Peter, H.; Dao, Z.L.; Luo, G.F.; Guo, H.J.; Ma, Y.P.; et al. De novo genome assembly of the endangered acer yangbiense, a plant species with extremely small populations endemic to Yunnan province, China. GigaScience 2019, 8, giz085. [Google Scholar] [CrossRef] [Green Version]
- Bachman, S.P.; Nic Lughadha, E.M.; Rivers, M.C. Quantifying progress toward a conservation assessment for all plants. Conserv. Biol. J. Soc. Conserv. Biol. 2018, 32, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Fu, J.; Wang, W.; Li, j. Development of China’s nature reserves over the past 60 years: An overview. Land Use Policy 2019, 80, 224–232. [Google Scholar] [CrossRef]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofrío-Dillon, G.; De Ruffray, P.; Vidal, C.; Pierrat, J.C.; Gégoutet, J.C. Changes in plant community composition lag behind climate warming in lowland forests. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef]
- Zheng, Y.; Ji, N.N.; Wu, B.W.; Wang, J.T.; Hu, H.W.; Guo, L.D.; He, J.Z. Climatic factors have unexpectedly strong impacts on soil bacterial β-diversity in 12 forest ecosystems. Soil Biol. Biochem. 2020, 142, 107699. [Google Scholar] [CrossRef]
- Moreno, J.L.; Bastida, F.; Díaz-López, M.; Li, Y.; Zhou, Y.; López-Mondéjar, R.; Benavente-Ferraces, I.; Rojas, R.; Rey, A.; García-Gil, J.C.; et al. Response of soil chemical properties, enzyme activities and microbial communities to biochar application and climate change in a mediterranean agroecosystem. Geoderma 2022, 407, 115536. [Google Scholar]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Williams, S.E. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Segan, D.B.; Murray, K.A.; Watson, J.E. A global assessment of current and future biodiversity vulnerability to habitat loss–climate change interactions. Glob. Ecol. Conserv. 2016, 5, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Y.; Zhang, M.; Ma, K. Climate change threats to protected plants of china: An evaluation based on species distribution modeling. Chin. Sci. Bull. 2014, 59, 4652–4659. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, Y.; Zang, Z.; Deng, S.; Shen, G. Climate warming has changed phenology and compressed the climatically suitable habitat of metasequoia glyptostroboides over the last half century. Glob. Ecol. Conserv. 2020, 23, e01140. [Google Scholar] [CrossRef]
- Cogoni, D.; Fenu, G.; Dessì, C.; Deidda, A.; Giotta, C.; Piccitto, M.; Bacchetta, G. Importance of Plants with Extremely Small Populations (PSESPs) in Endemic-Rich Areas, Elements Often Forgotten in Conservation Strategies. Plants 2021, 10, 1504. [Google Scholar] [CrossRef]
- Teixeira, T.M.; Nazareno, A.G. One Step Away from Extinction: A Population Genomic Analysis of A Narrow Endemic, Tropical Plant Species. Front. Plant Sci. 2021, 12, 1876. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.E.; Iwamura, T.; Butt, N. Mapping vulnerability and conservation adaptation strategies under climate change. Nat. Clim. Change 2013, 3, 989. [Google Scholar] [CrossRef]
- Butt, N.; Possingham, H.P.; De Los Rios, C.; Maggini, R.; Fuller, R.A.; Maxwell, S.L.; Watson, J.E.M. Challenges in assessing the vulnerability of species to climate change to inform conservation actions. Biol. Conserv. 2016, 199, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Sun, J.; El-Kassaby, Y.A.; Yang, X.; Tian, X.; Wang, T. Predicting Potential Habitat of a Plant Species with Small Populations under Climate Change: Ostryarehderiana. Forests 2022, 13, 129. [Google Scholar] [CrossRef]
- Yang, J.; Cai, L.; Liu, D.; Chen, G.; Sun, W. China’s conservation program on plant species with extremely small populations (psesp): Progress and perspectives. Biol. Conserv. 2020, 244, 108535. [Google Scholar] [CrossRef]
- Sun, W.B. Special issue for plant species with extremely small populations. Plant Divers. 2016, 38, 207–258. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar]
- Hirzel, A.H.; Le Lay, G.; Helfer, V.; Randin, C.; Guisan, A. Evaluating the ability of habitat suitability models to predict species presences. Ecol. Model. 2006, 199, 142–152. [Google Scholar] [CrossRef]
- Wan, J.; Wang, C.; Yu, F. Spatial conservation prioritization for dominant tree species of Chinese forest communities under climate change. Clim. Change 2017, 144, 303–316. [Google Scholar] [CrossRef]
- Lu, Y.; Xu, P.; Li, Q.; Wang, Y.; Wu, C. Planning priority conservation areas for biodiversity under climate change in topographically complex areas: A case study in sichuan province, china. PLoS ONE 2020, 15, e0243425. [Google Scholar]
- Cable, A.B.; O’Keefe, J.M.; Deppe, J.L.; Hohoff, T.C.; Taylor, S.J.; Davis, M.A. Habitat suitability and connectivity modeling reveal priority areas for indiana bat (Myotis sodalis) conservation in a complex habitat mosaic. Landsc. Ecol. 2021, 36, 119–137. [Google Scholar] [CrossRef]
- Qu, H.; Wang, C.J.; Zhang, Z.X. Planning priority conservation areas under climate change for six plant species with extremely small populations in China. Nat. Conserv. 2018, 25, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Sun, J. Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt model. BMC Ecol. 2018, 18, 10. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Wu, D.; Zhu, C. Prediction of the spatial distribution of Alternanthera philoxeroides in China based on ArcGIS and MaxEnt. Glob. Ecol. Conserv. 2020, 21, e00856. [Google Scholar] [CrossRef]
- West, A.M.; Kumar, S.; Brown, C.S.; Stohlgren, T.J.; Bromberg, J. Field validation of an invasive species Maxent model. Ecol. Inform. 2016, 36, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2015, 17, 43–57. [Google Scholar] [CrossRef]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (h. riparia lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Ranaei, F.; Ahmad, Z. Application of Bamboo Plants in Nine Aspects. Sci. World J. 2020, 2020, 7284203. [Google Scholar] [CrossRef]
- Sohel, M.S.I.; Alamgir, M.; Akhter, S.; Rahman, M. Carbon storage in a bamboo (Bambusa vulgaris) plantation in the degraded tropical forests: Implications for policy development. Land Use Policy 2015, 49, 142–151. [Google Scholar] [CrossRef]
- Nguyen, H.N.; Tran, V.T.; Nguyen, N.T. Ferrocalamus fibrillosus (poaceae: Bambusoideae), a new species from vietnam. Ann. Bot. Fenn. 2012, 49, 206–208. [Google Scholar] [CrossRef]
- Keng, P.J.; Xue, J.R. Ferrocalamus, Bambusoideae, Trib. Arundinarieae, Sasinae. J. Bamboo Res. 1982, 1, 1–5. [Google Scholar]
- Wen, T.H. New taxa of Bambusoideae in China (I). J. Bamboo Res. 1984, 1, 23–47. [Google Scholar]
- Volis, S.; Deng, T. Importance of a single population demographic census as a first step of threatened species conservation planning. Biodivers. Conserv. 2020, 29, 527–543. [Google Scholar] [CrossRef]
- Beckmann, M.; Václavík, T.; Manceur, A.M.; Šprtová, L.; Von Wehrden, H.; Welk, E.; Cord, A.F. glUV: A global UV-B radiation dataset for macroecological studies. Methods Ecol. Evol. 2014, 5, 372–383. [Google Scholar] [CrossRef]
- Fischer, G.; Nachtergaele, F.; Prieler, S.; Van Velthuizen, H.T.; Verelst, L.; Wiberg, D. Global Agro-Ecological Zones Assessment for Agriculture (GAEZ 2008); IIASA: Laxenburg, Austria; FAO: Rome, Italy, 2008. [Google Scholar]
- Xu, W.; Du, Q.; Yan, S.; Cao, Y.; Liu, X.; Guan, D.X.; Ma, L.Q. Geographical distribution of as-hyperaccumulator pteris vittata in China: Environmental factors and climate changes. Sci. Total Environ. 2022, 803, 149864. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distribution. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENM eval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Kass, J.M.; Muscarella, R.; Galante, P.J.; Bohl, C.L.; Pinilla-Buitrago, G.E.; Boria, R.A. ENMeval 2.0: Redesigned for customizable and reproducible modeling of species’ niches and distributions. Methods Ecol. Evol. 2021, 12, 1602–1608. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar]
- Brown, J.L.; Bennett, J.R.; French, C.M. Sdmtoolbox 2.0: The next generation python-based gis toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Boyce, M.S.; Vernier, P.R.; Nielsen, S.E.; Schmiegelow, F.K.A. Evaluating resource selection functions. Ecol. Model. 2002, 157, 281–300. [Google Scholar] [CrossRef] [Green Version]
- Janitza, S.; Strobl, C.; Boulesteix, A.-L. An AUC-based permutation variable importance measure for random forests. BMC Bioinform. 2013, 14, 119. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Liao, J.; Wang, H.; Xiao, S.; Guan, Z.; Zhang, H.; Dumont, H.J.; Han, B.-P. Modeling and Prediction of the Species’ Range of Neurobasis chinensis (Linnaeus, 1758) under Climate Change. Biology 2022, 11, 868. [Google Scholar] [CrossRef]
- Boisvert-Marsh, L.; De Blois, S. Unravelling potential northward migration pathways for tree species under climate change. J. Biogeogr. 2021, 48, 1088–1100. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.P.; Zhang, Y.F.; Hu, R.; Chen, N. Responses of plant growth of different life forms to rainfall amount changes in an arid desert area. Chin. J. Ecol. 2015, 34, 1847–1853. [Google Scholar]
- Wang, J.J.; Cao, B.; Bai, C.K.; Zhang, L.L.; Che, L. Potential distribution prediction and suitability evaluation of Fritillaria cirrhosa D. Don based on Maxent modeling and GIS. Bull. Bot. Res. 2014, 34, 642–649. [Google Scholar]
- Feeley, K.J.; Malhi, Y.; Zelazowski, P.; Silman, M.R. The relative importance of deforestation, precipitation change, and temperature sensitivity in determining the future distributions and diversity of amazonian plant species. Glob. Change Biol. 2012, 18, 2636–2647. [Google Scholar] [CrossRef]
- Enquist, B.J.; Feng, X.; Boyle, B.; Maitner, B.; Newman, E.A.; Jørgensen, P.M.; Roehrdanz, P.R.; Thiers, B.M.; Burger, J.R.; Corlett, R.T.; et al. The commonness of rarity: Global and future distribution of rarity across land plants. Sci. Adv. 2019, 5, eaaz0414. [Google Scholar] [PubMed]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.F.; Momohara, A.; Herrando, M.S.; Qian, S.H.; Yang, Y.C.; Ohsawa, M.; Luu, H.T.; et al. Identifying long-term stable refugia for relict plant species in East Asia. Nat. Commun. 2018, 9, 4488. [Google Scholar] [CrossRef] [Green Version]
- Breiner, F.T.; Nobis, M.P.; Bergamini, A.; Guisan, A. Optimizing ensembles of small models forpredicting the distribution of species with few occurrences. Methods Ecol. Evol. 2018, 9, 802–808. [Google Scholar]
- Hao, T.; Elith, J.; Lahoz-Monfort, J.J.; Guillera-Arroita, G. Testing whether ensemble modelling is advantageous for maximising predictive performance of species distribution models. Ecography 2020, 43, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Hastings, R.; Cummins, V.; Holloway, P. Assessing the Impact of Physical and Anthropogenic Environmental Factors in Determining the Habitat Suitability of Seagrass Ecosystems. Sustainability 2020, 12, 8302. [Google Scholar] [CrossRef]
- Engler, R.; Guisan, A. MigClim: Predicting plant distribution and dispersal in a changing climate. Divers. Distrib. 2009, 15, 590–601. [Google Scholar]
- Engler, R.; Hordijk, W.; Guisan, A. The MIGCLIM R package—Seamless integration of dispersal constraints into projections of species distribution models. Ecography 2012, 35, 872–878. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description | Percent Contribution (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Current | SSP c 1-2.6 | SSP5-8.5 | ||||||
| 1 d | 2 | 3 | 1 | 2 | 3 | |||
| bio1 (°C) a | Annual mean temperature | 0.1 | 0.8 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| bio2 (°C) b | Mean diurnal range (mean of monthly (max temp–min temp) | - | - | - | - | - | - | - |
| bio3 | Isothermality (bio2/bio7) (×100) | 31.8 | 34 | 27.3 | 26.2 | 17 | 22.5 | 24.8 |
| bio4 | Temperature seasonality (standard deviation × 100) | 1.9 | 1.8 | 4.1 | 3.8 | 5.7 | 3.3 | 4.1 |
| bio5 (°C) | Maximum temperature of warmest month | - | - | - | - | - | - | - |
| bio6 (°C) | Minimum temperature of coldest month | - | - | - | - | - | - | - |
| bio7 (°C) | Temperature annual range (bio5-bio6) × 10 | - | - | - | - | - | - | - |
| bio8 (°C) | Mean temperature of wettest quarter | - | - | - | - | - | - | - |
| bio9 (°C) | Mean temperature of driest quarter × 10 | - | - | - | - | - | - | - |
| bio10 (°C) | Mean temperature of warmest quarter | - | - | - | - | - | - | - |
| bio11 (°C) | Mean temperature of coldest quarter | - | - | - | - | - | - | - |
| bio12 (mm) | Annual precipitation | - | - | - | - | - | - | - |
| bio13 (mm) | Precipitation of wettest month | - | - | - | - | - | - | - |
| bio14 (mm) | Precipitation of driest month | 10.2 | 3.5 | 3.5 | 6.6 | 2.5 | 4.9 | 4.1 |
| bio15 (mm) | Precipitation seasonality (coefficient of variation) | - | - | - | - | - | - | - |
| bio16 (mm) | Precipitation of wettest quarter | - | - | - | - | - | - | - |
| bio17 (mm) | Precipitation of driest quarter | - | - | - | - | - | - | - |
| bio18 (mm) | Precipitation of warmest quarter | 31.5 | 34 | 35.5 | 36.3 | 45.8 | 38.3 | 37 |
| bio19 (mm) | Precipitation of coldest quarter | - | - | - | - | - | - | - |
| aspect | (Uphill height/horizontal distance) × 100% | 3.2 | 2.5 | 3.7 | 3.6 | 3.5 | 3.8 | 3.4 |
| slope | (height difference/distance) × 100% | 10.5 | 12.4 | 13.6 | 12.3 | 11.2 | 13.7 | 12.9 |
| elev (m) | Elevation | 0.7 | 1.6 | 2.7 | 2.5 | 6 | 3.1 | 2.5 |
| T-ph | Soil pH | 1.1 | 2.3 | 1.8 | 1.6 | 2 | 2.1 | 2.2 |
| T-oc | Organic carbon (g/kg × 100%) | 8.3 | 7.4 | 6.3 | 5.8 | 5.5 | 6.7 | 7.1 |
| Uvb3 (J/m2/d) | Mean UV-B radiation of highest month | 0.6 | 2.1 | 1.3 | 1.2 | 0.8 | 1.6 | 1.6 |
| Area | Last Glacial Maximum | Mid-Holocene | Current | SSP 1-2.6 | SSP 5-5.8 | ||||
|---|---|---|---|---|---|---|---|---|---|
| 2021–2040 | 2041–2060 | 2061–2080 | 2021–2040 | 2041–2060 | 2061–2080 | ||||
| Barely suitable a | −28% b | +63% | 3.5229 (km2) | +34% | +28% | +23% | +40% | +43% | +48% |
| Moderately suitable | −51% | +32% | 1.8907 (km2) | +17% | +21% | +24% | +35% | +16% | +25% |
| Highly suitable | −41% | +18% | 8586 (km2) c | −12.8% | −8.19% | +11.6% | +14.3% | −15% | −16.4% |
| Very highly suitable | +377% | +42% | 1162 (km2) | −34% | −46% | −23% | −26% | −44% | −58% |
| Total suitable | −37% | +47% | 6.3883 (km2) | +21.6% | +19.9% | +21.1% | +34% | +26% | +30.9% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, H.; Zheng, X.; Wang, Y.; Wang, W.; Li, M.; Wang, S.; Wang, J.; Wang, C.; Zhan, H. Effects of Climate Change and Environmental Factors on Bamboo (Ferrocalamus strictus), a PSESP Unique to China. Forests 2022, 13, 2108. https://doi.org/10.3390/f13122108
He H, Zheng X, Wang Y, Wang W, Li M, Wang S, Wang J, Wang C, Zhan H. Effects of Climate Change and Environmental Factors on Bamboo (Ferrocalamus strictus), a PSESP Unique to China. Forests. 2022; 13(12):2108. https://doi.org/10.3390/f13122108
Chicago/Turabian StyleHe, Honglan, Xiaofeng Zheng, Yingqiong Wang, Wenquan Wang, Maobiao Li, Shuguang Wang, Jin Wang, Changming Wang, and Hui Zhan. 2022. "Effects of Climate Change and Environmental Factors on Bamboo (Ferrocalamus strictus), a PSESP Unique to China" Forests 13, no. 12: 2108. https://doi.org/10.3390/f13122108