1. Introduction
Biodiversity is an important feature of life, and species composition and community construction have long been core issues in ecological research [
1,
2]. Indeed, the biodiversity and structural composition and distribution of communities are closely related to ecosystem multifunctionality [
2]. Topography is one of the most major natural factors that affects biodiversity. Topography belongs to biogeography and affects numerous ecological functions and processes, such as soil erosion and water storage [
3], weathering and nutrient leaching [
4], and migration and storage of soil organic matter [
5].
Aspect (shady and sunny aspects) is a feature of natural geographical mountain landscapes [
6]. Shady and sunny aspects are different habitats with climate and microenvironment differences that form a natural environmental gradient [
7]. The difference in climate conditions between shady and sunny aspects leads to profound changes in vegetation, soil, landform, hydrology, and other factors [
8,
9]. The shady aspects are darker, wetter, and colder, with less light radiation, and the climate characteristics on sunny aspects are the opposite [
10]. Changes in biodiversity distribution and community construction on shady and sunny aspect gradients, as well as the relationships between environmental factors and biological communities, can reflect the roles and contributions of various biological, physical, and chemical factors.
Most ecological studies on different aspects have predominantly focused on soil stoichiometry characteristics, vegetation distribution and succession, plant community diversity and structure, aboveground plant biomass changes, and plant functional traits [
11,
12,
13]. Zhao et al. [
14] focused on vegetation–microorganism–aspects complexes and concluded that soil microbial activity in mountain ecosystems was profoundly affected by these complexes. However, previous studies have not fully solved the problem of how aspects affect biodiversity directly or indirectly, and little is known about the coexistence of species and mechanisms of community construction. Therefore, it is necessary to study different aspects in typical mountain ecosystems and reveal the relationships among environmental factors (e.g., light, temperature, and soil’s physical and chemical properties), biological factors (e.g., plant community composition, stoichiometric characteristics, and functional traits), and community composition and distribution rules of specific biotas.
Toona ciliata Roem. var.
pubescens (Franch.) Hand.-Mazz. (
Tc) is regarded as a national secondary protected plant in China. Moreover,
Tc is an endangered species and a valuable woody plant of the
Toona genus in the Meliaceae family [
15]. Owing to a long period of exhaustive utilization and exploitation, habitats of
Tc forest are affected by these effects and have disadvantageous factors for natural regeneration, meaning that the distribution area of natural
Tc forests is reduced and fragmented [
16]. Wild
Tc is predominantly distributed on both sides of valleys and streams in virgin forests in subtropical China, breeding predominantly by seeds and clonal tillers [
15]. Their special habitat preferences and breeding strategies may be the dominant cause of this endangerment. In recent years, scientists have addressed the problems of the natural regeneration of
Tc forests [
17], and found that transmission barriers [
18] and microhabitat restrictions [
19] might be major causes of failure of
Tc regeneration. Sensitive habitat selection and lower forest regeneration capacity have caused difficulties in the protection and restoration of wild
Tc forests [
20]. Microhabitats involve many environmental factors, such as water, microorganisms, light, and soil. The growth of little trees is affected by these environmental factors, and there are large variations in the key factors of various plants. Soil microorganisms play a major role in seedling growth, improving soil quality, and promoting natural regeneration in
Tc forests [
20]. Huang et al. [
21] found that soil pathogen infection was the dominant reason for seed rot and the natural regeneration failure of
Tc. As an important soil microorganism, arbuscular mycorrhizal (AM) fungi has a symbiotic association with over 80% of terrestrial plants [
22], and can effectively promote plant growth and natural regeneration of
Tc [
23,
24]. The symbiotes formed by AM fungi and plants give rise to vital roles in plant nutrient absorption and resistance improvement to external adverse environments at the ecosystem or individual scale [
22,
25]. Wild
Tc forests are largely distributed in habitats which have poor light and soil conditions, and
Tc seedlings are often hindered by other plants during the initial growth stage [
21]. Considering this, AM fungi may be important in wild
Tc forests, as AM fungi can promote plant development and growth by increasing the chlorophyll content in plant leaves, improving photosynthesis, promoting C assimilation and metabolism of plants [
26], and improving the competitiveness of plants in natural habitats.
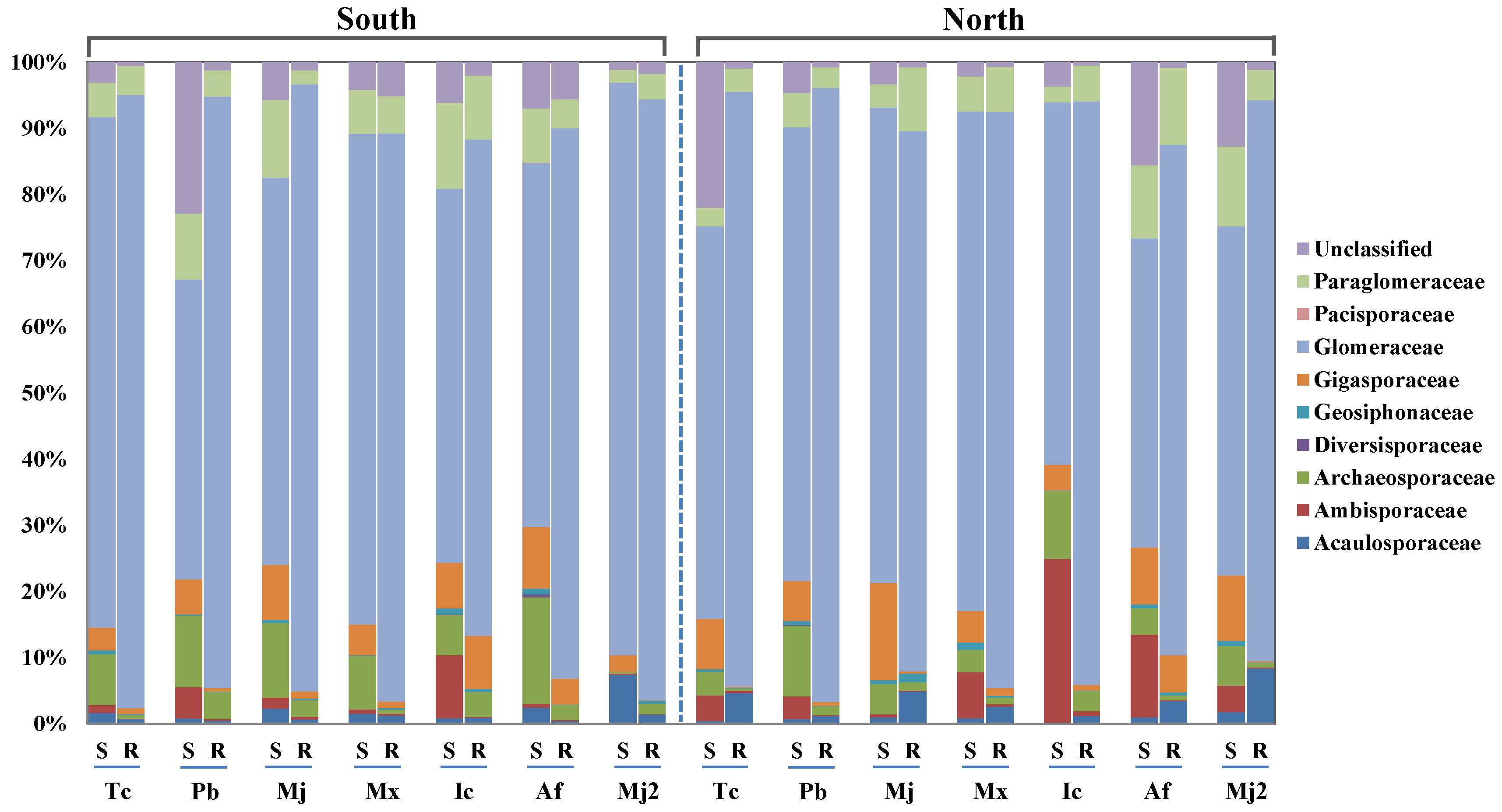
Currently, the study of different aspect-related soil microbial community compositions and their distribution characteristics can only provide some sporadic information [
27,
28] and only a few studies have investigated the impact of aspect on AM fungal community structure. Chu et al. [
28] reported the community composition of AM fungi on sunny and shady aspects in boreal forests of the Greater Khingan Mountains. Similarly, one study reported the distribution of AM fungi on sunny and shady aspects in the arid ecosystem of Inner Mongolia [
29]. However, the above studies covered only a few ecosystem types, and studies on particular species and habitats are lacking, limiting our insight into the community-building mechanisms associated with AM fungi. Shady and sunny aspects are comprehensive natural environmental gradients, and the AM diversity and relationship among AM fungi, environmental factors, and plant communities can reveal the comprehensive effects in natural
Tc forests.
Considering the particularity of wild Tc forest distribution regions and the important role of AM fungi in natural regeneration and growth of wild Tc forests, this study aimed to reveal the community characteristics and biogeographic distribution rules (sunny and shady aspects) of AM fungi in a wild Tc forests. A gradient of shady and sunny aspects in the Guanshan National Nature Reserve in Jiangxi Province was selected as the research site. Field investigations of AM fungal community and diversity distribution patterns in the roots and rhizosphere soils of seven widespread plants in natural Tc forest were analyzed using the Illumina PE250 high-throughput sequencing method. The rich, continuous environmental (biotic and abiotic) gradients and distinct vegetation types in this region provide an excellent opportunity to discover the dynamics of AM fungi on different aspects in natural Tc forests. Our results will provide important information on the ecological processes of AM fungal community construction in natural Tc forests and a reference for discussing the microbiological mechanism of adaptation of Tc to microhabitats. The null hypothesis was that Tc in shady and sunny aspects would be associated with similar AM fungi diversity and composition, but that Tc recruited significantly different AM fungi compared to other plants. Changes in soil chemical properties and tree species may be responsible for these variations. The findings of this study will improve our ability to understand ecosystem dynamics and function in wild Tc forests and facilitate scientific protection and cultivation strategy decisions on subtropical mountain ecosystems in China.
2. Materials and Methods
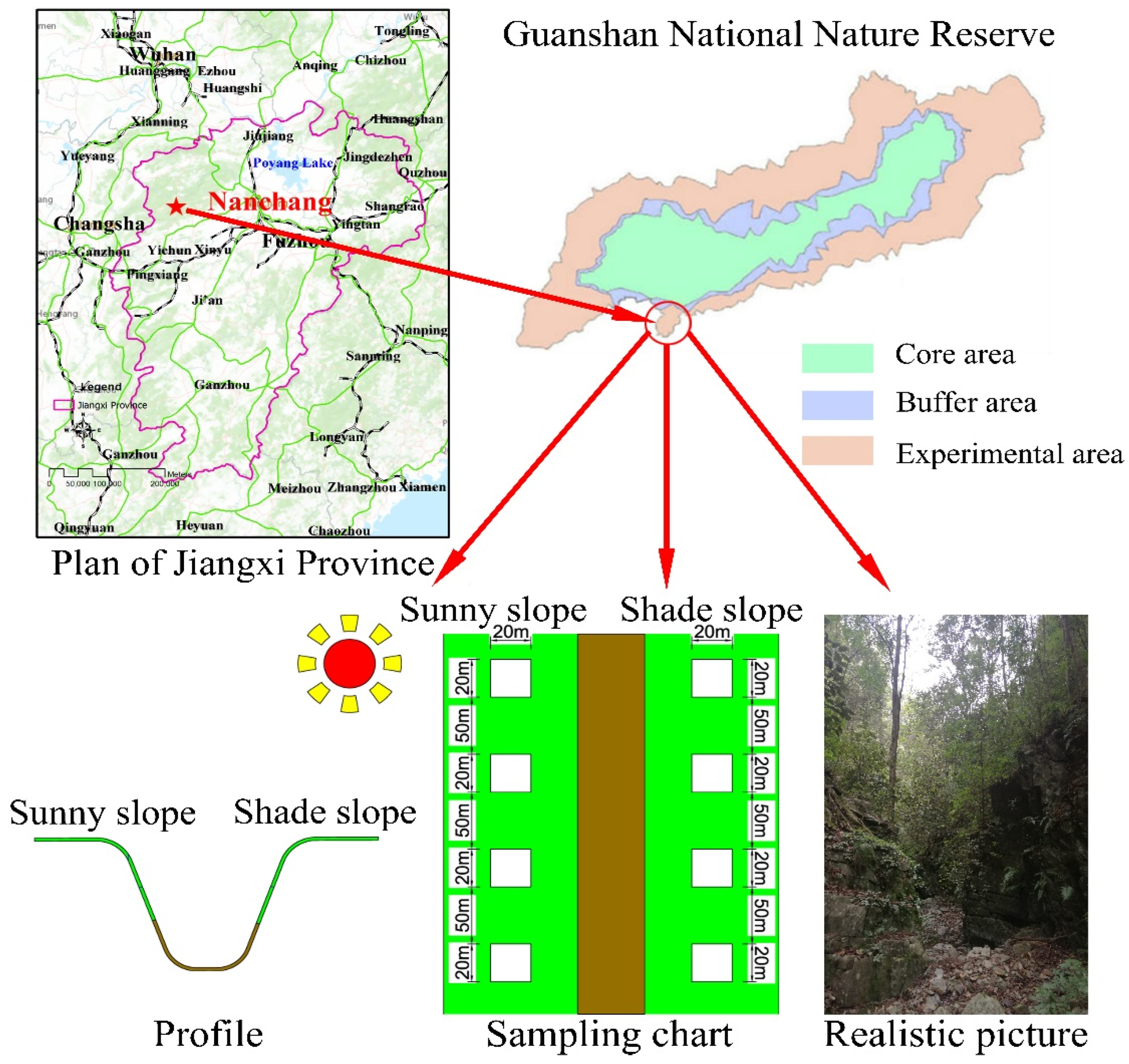
2.1. Study Site
The Guanshan National Nature Reserve (28°30′–28°40′ N, 114°29′–114°45′ E) is located in the western part of the Jiuling Mountains in the northwest Jiangxi Province, China. It was one of the first national nature reserves in Jiangxi Province and belongs to the subtropical evergreen broad-leaved forest ecosystem. The mean annual temperature is 17.2 °C. The mean annual precipitation is 1680.2 mm. The total area is 11,500.5 ha, comprising 3621.1 ha in the core area, 1466.4 ha in the buffer zone, and 6413.0 ha in the experimental area. The soil type of this study area was typical red soil.
2.2. Experimental Design and Sampling
The natural
Tc forest community is predominantly distributed in a subtropical evergreen broad-leaved forest (
Figure 1). The
Tc is dominantly distributed sporadically on both sides of valleys and streams and on both sides of the aspects with different orientations and illumination and hydrothermal conditions. The photophilous tree species is mainly distributed in the arborous layer. A field survey was started at 2017, and eight experimental sample plots of 20 × 20 m were set up in a natural
Tc forest (north-facing [NF] aspect—4, and south-facing [SF] aspect—4). In the sample plot, we selected seven dominant plants that were distributed in NF and SF, as follows:
Toona ciliates (
Tc),
Padus buergeriana (
Pb),
Maesa japonica (
Mj),
Melliodendron xylocarpum (
Mx),
Ilex chinensis (
Ic),
Alniphyllum fortunei (
Af), and
Mallotus japonicas (
Mj2). At least one individual sample from each plant was selected in each plot. Five individual root samples and five individual rhizosphere soil samples were randomly selected from each plant, respectively. Fine fibrous roots in the 0–20 cm soil depth were collected from the root samples, and 1–2 kg of soil around the root was collected with a clean small steel shovel to extract AM fungal spores. Finally, 70 root samples and 70 rhizosphere soil samples per plot (2 aspects × 7 plants × 5 individuals) were collected. The collected fine roots were rinsed with water and stored at −80 °C for DNA extraction. The collected soil samples were passed through a 2 mm sieve to eliminate stones, roots, and litter for DNA extraction (−80 °C) and soil experiments (air-dried soil, 0.149 mm mesh).
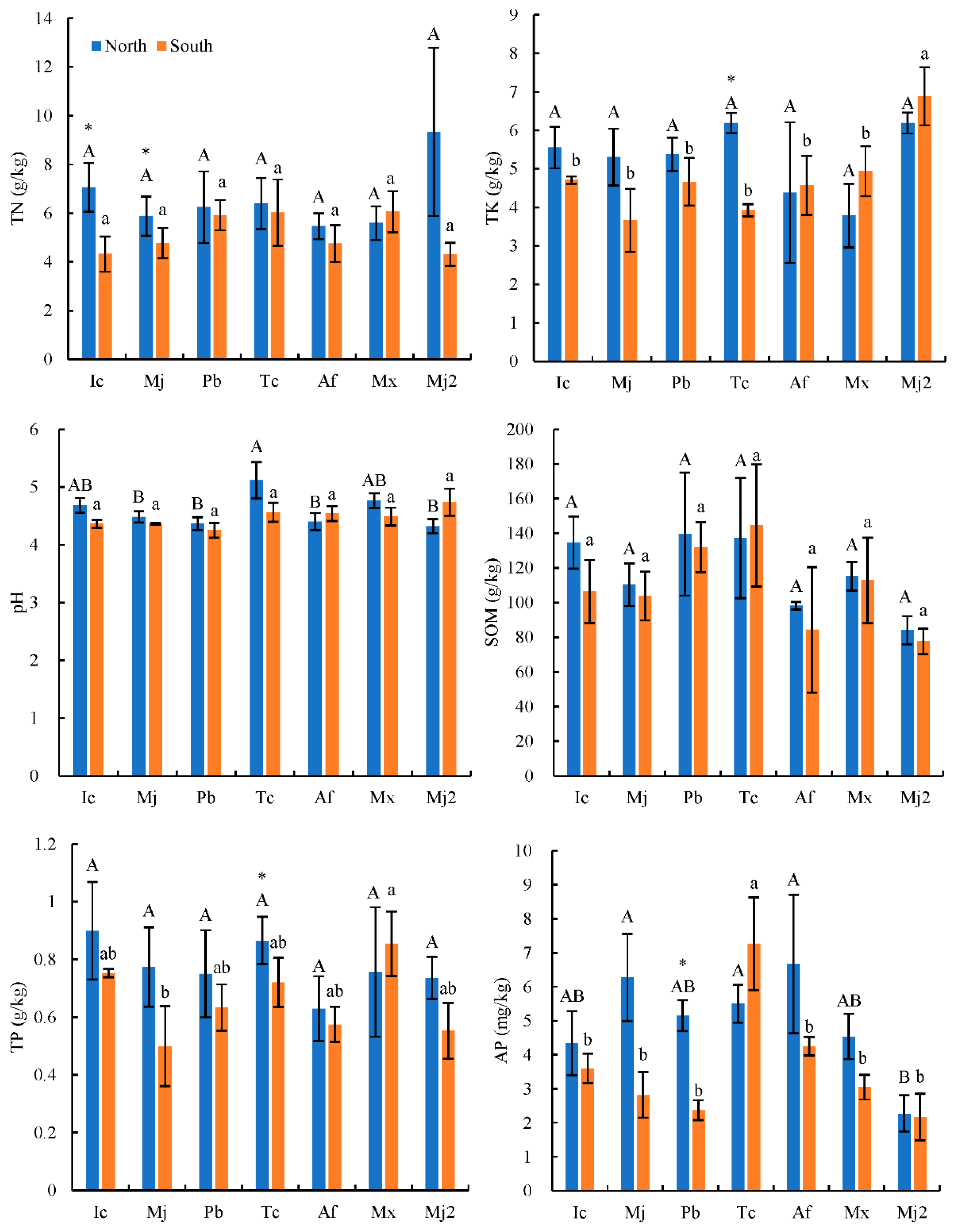
2.3. Soil Chemical Property Determination
The soil pH was measured using a glass electrode at 1:2.5 (
w/
v). The soil organic matter (SOM, g·kg
−1) content was measured using the K
2Cr
2O
7 heating method. Soil available phosphorus (AP) content was determined after extraction with 0.5 mol·L
−1 NaHCO
3. Total potassium (TK) content was extracted using a 1 M NH
4OAc solution (pH 7.0) and measured by flame photometry. Total phosphorus (TP) content was determined using the H
2SO
4-HClO
4 heating digestion method, and the total nitrogen (TN) content was measured using the Kjeldahl method. The detailed steps can be found in SD Bao [
30].
2.4. DNA Extraction Method
Soil DNA was extracted using a Fast DNA SPIN Kit (MP Biomedicals LLC, Santa Ana, CA, USA) according to the instructions and using 0.5 g fresh soil samples. Root DNA was extracted using a Fast Plant Kit (Beijing Tiangen) according to the instructions and using 0.05 g frozen root samples. The total concentration of the extracted soil/root DNA in each soil or root sample was determined using a NanoDrop ND-8000 spectrophotometer (NanoDrop, Wilmington, DE, USA). The DNA quality was checked by 1% agarose gel electrophoresis, diluted to 10–20 ng·μL−1 with ultrapure water, and preserved at −20 °C in a refrigerator for molecular biological analysis.
2.5. PCR Reaction Method
To guarantee the reliability and accuracy of the further data analysis, the PCR reaction needs to meet the following two requirements: (1) use a small number of amplification cycles and (2) ensure that the number of cycles amplified by each sample is consistent. The SSU rDNA of AM fungi in soil and plant roots was amplified using AM FUNGI-specific primers AMV4.5NF-F (5′-AAGCTCGTAGTTGAATTTCG-3′) and AMDGR-R (5′-CCCAACTATCCCTATTAATCAT-3′). Then, PCR was performed using TransStart Fastpfu DNA Polymerase, with a 20 μL reaction system. Each sample was repeated three times. The reaction system is shown in
Table 1.
The PCR reaction system was mixed mildly and put into the PCR instrument (ABI GeneAmp® 9700 type), and the reaction conditions were as follows: (1) 5 min of incipient denaturation at 95 °C; (2) 30 s of denaturation at 95 °C; (3) 30 s of annealing at 55 °C; (4) 45 s of elongation at 72 °C; (5) 27 cycles from step 4 to step 2; and (6) incubation at 72 °C for 10 min. Each sample was subjected to three replicates, and the PCR products of the same sample were blended and detected using 2% agarose gel electrophoresis.
2.6. Illumina High-Throughput Sequencing and Bioinformatics
The PCR products were subjected to 1% agarose gel electrophoresis in 1 × TAE buffer (added with Biotake producing a green as blue nucleic acid dye). After electrophoresis at 125 V for 30 min, the PCR results of the gel were observed under UV light. Then, the target band for recovering the PCR products was cut off using a AxyPrepDNA gel recovery kit (AXYGEN), which was eluted with Tris-HCl and detected by 2% agarose electrophoresis. The PCR products were observed and identified using 2% agarose gel, while the electrophoresis condition was 120 V for 40 min. The target bands were separated by electrophoresis, cut, and purified (Aidlab Biotechnologies Co., Ltd., Tianjin, China). The purified product was quantified, homogenized using a TBS-380 fluorescence spectrometer, and sequenced using the Illumina PE250 high-throughput sequencing platform (Shanghai BIOZERON Co., Ltd., Shanghai, China) referring to standard methods. A 97% identity threshold was used to group sequences into operational taxonomic units (OTUs) [
31]. The largest ample sequence from each OTU was regarded as the typical sequence for the OTU. The Usearch (version 7.1
http://drive5.com/uparse/, accessed on 25 December 2017) software was used to cluster sequences. Taxonomy was allotted to the fungal OTUs against a subset of the Silva 104 database (
http://www.arb-silva.de/download/archive/qiime/, accessed on 28 December 2017). The GenBank (
http://www.ncbi.nlm.nih.gov/, accessed on 31 December 2017) for the OTUs was used to confirm the typical sequences that could not be recognized at the family or class level in the fungal database. The original sequences described here can be accessed through GenBank with SRP277481 and accession numbers SAMN15815414 to SAMN15815425 and SAMN15815492 to SAMN15815503.
2.7. Calculation Methods of Diversity Correlation Indexes Data Analysis
We used the Chao index to calculate the total number of species in an ecosystem [
32]. Here, Chao 1 was used to calculate the number of OTUs, as follows:
where
Schao1 is the number of OTUs,
Sobs is the number of observed OTUs,
n1 is the number of OTUs containing only one sequence (singletons), and
n2 is the number of OTUs containing only two sequences (doubletons).
The Simpson index was used to quantitatively calculate the microbial diversity [
33]. The larger the Simpson index, the lower the community diversity, as follows:
where
Sobs is the number of observed OTUs,
ni is the sequence number of OTU-i, and
N is the total number of sequences.
The Shannon index (i.e., the Shannon–Weiner index) was also applied to quantitatively calculate the microbial diversity. The larger the Shannon index, the greater the community diversity, as follows:
where
Sobs is the number of observed OTUs,
ni is the sequence number of OTU-i, and
N is the total number of sequences.
Subsequently, the coverage index was calculated as follows:
where
C is the coverage of OTUs,
n1 is the number of OTUs containing only one sequence (singletons), and
N is the total sequence.
2.8. Data Analysis
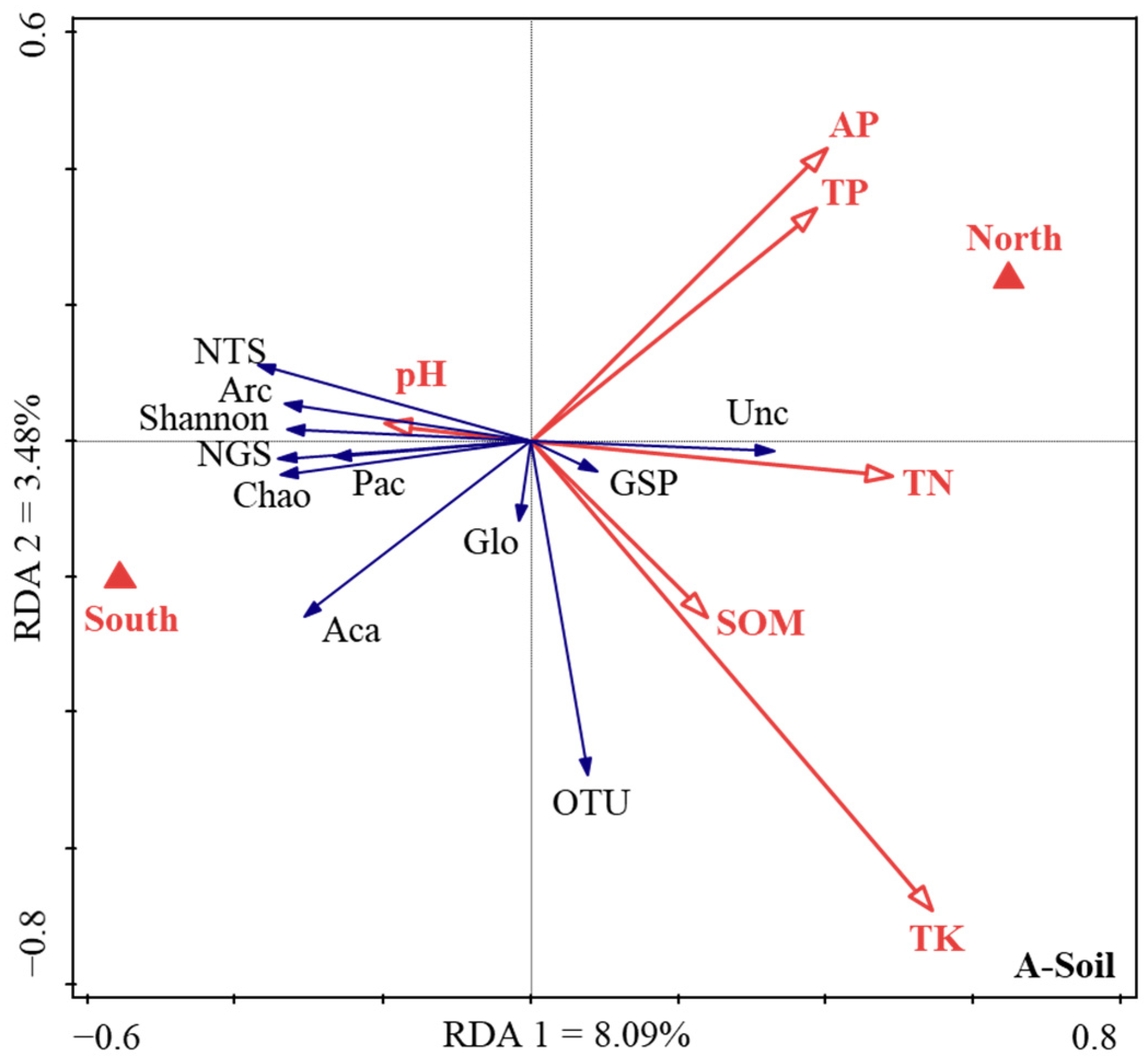
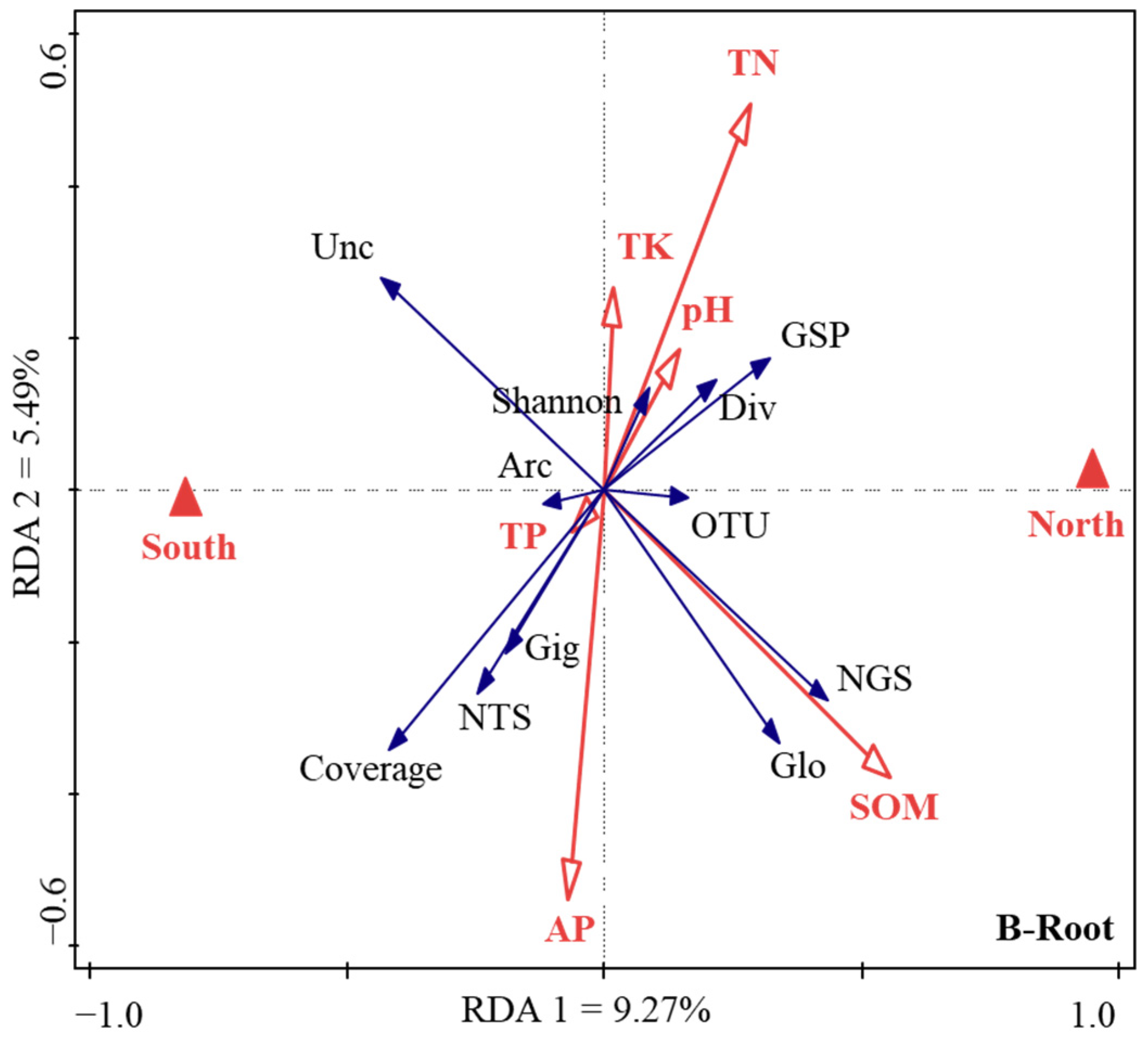
Firstly, the normal distribution test is carried out on all the data used in the analysis. If the data does not conform to the normal distribution, the natural logarithm or trigonometric function is used for data conversion. Secondly, the Duncan’s multiple comparative analysis (version 26.0; IBM, Armonk, NY, USA) was conducted to analyze the variations in soil chemical properties (pH, SOM, AP, TK, TP, and TN), soil and root AM fungi community diversity (the Chao, Shannon, Simpson, and coverage indices), and sequence parameters (the total sequence number, the number of Glomeromycota sequences, the proportion of Glomeromycota sequences, the number of OTUs, and the relative abundance of the belowground AM fungi community sequence) in Tc natural forest in different aspects. Pairwise comparative analysis was used to compare the difference of the corresponding value between the north and south aspects. Thirdly, there were, respectively, 18 response factors for AM fungal community characteristics in soil and root. Through factor analysis, 6 types of comprehensive factors (which mainly reflected the information of 11 key factors) were used to reflect the information of 18 response factors, so as to reduce the dimension, and the key response factors were determined for further analysis. Therefore, the redundancy analysis (RDA) was applied to analyze the relationships between soil environmental variables and the key characteristics of the AM fungal communities by using the standardized data (Canoco 5.0, Biometrics Wageningen, The Netherlands).

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




