Observed and Predicted Geographic Distribution of Acer monspessulanum L. Using the MaxEnt Model in the Context of Climate Change

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
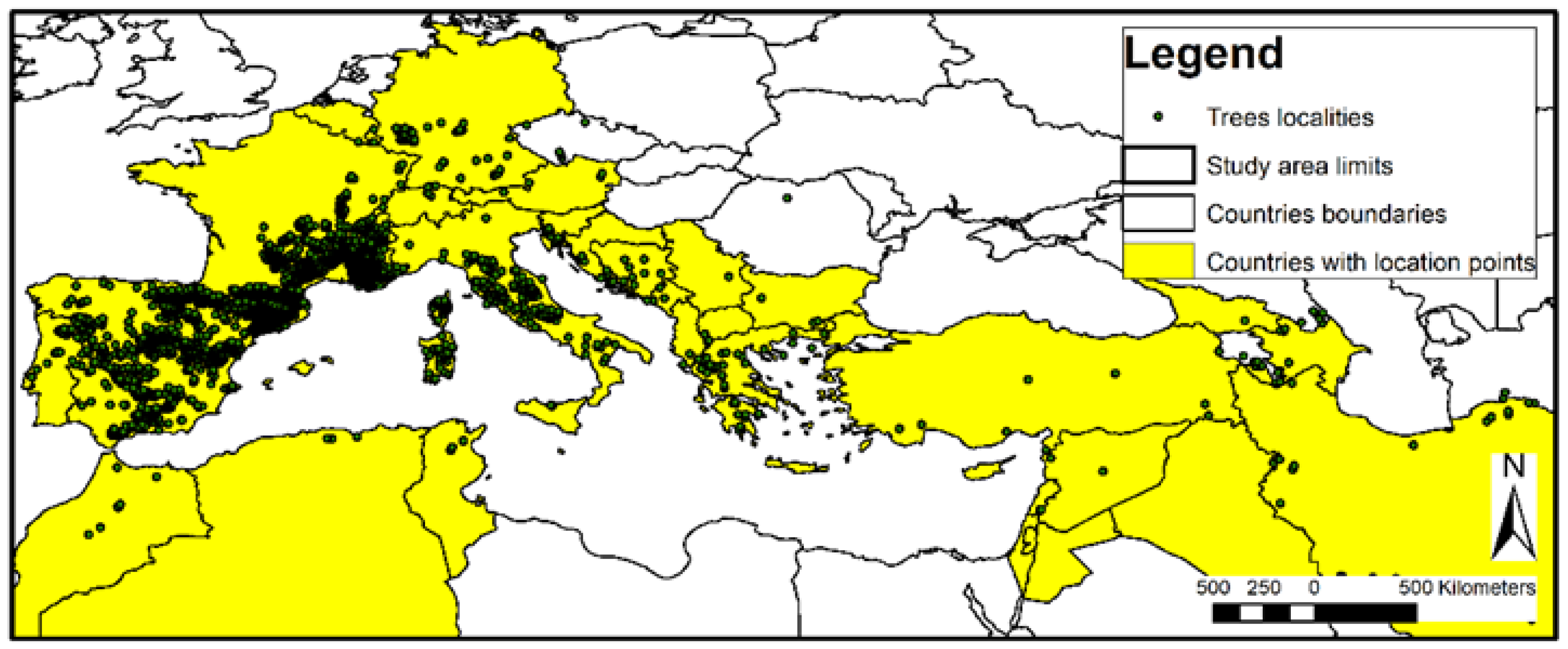
2.1. Species Description and Study Area
2.2. Data
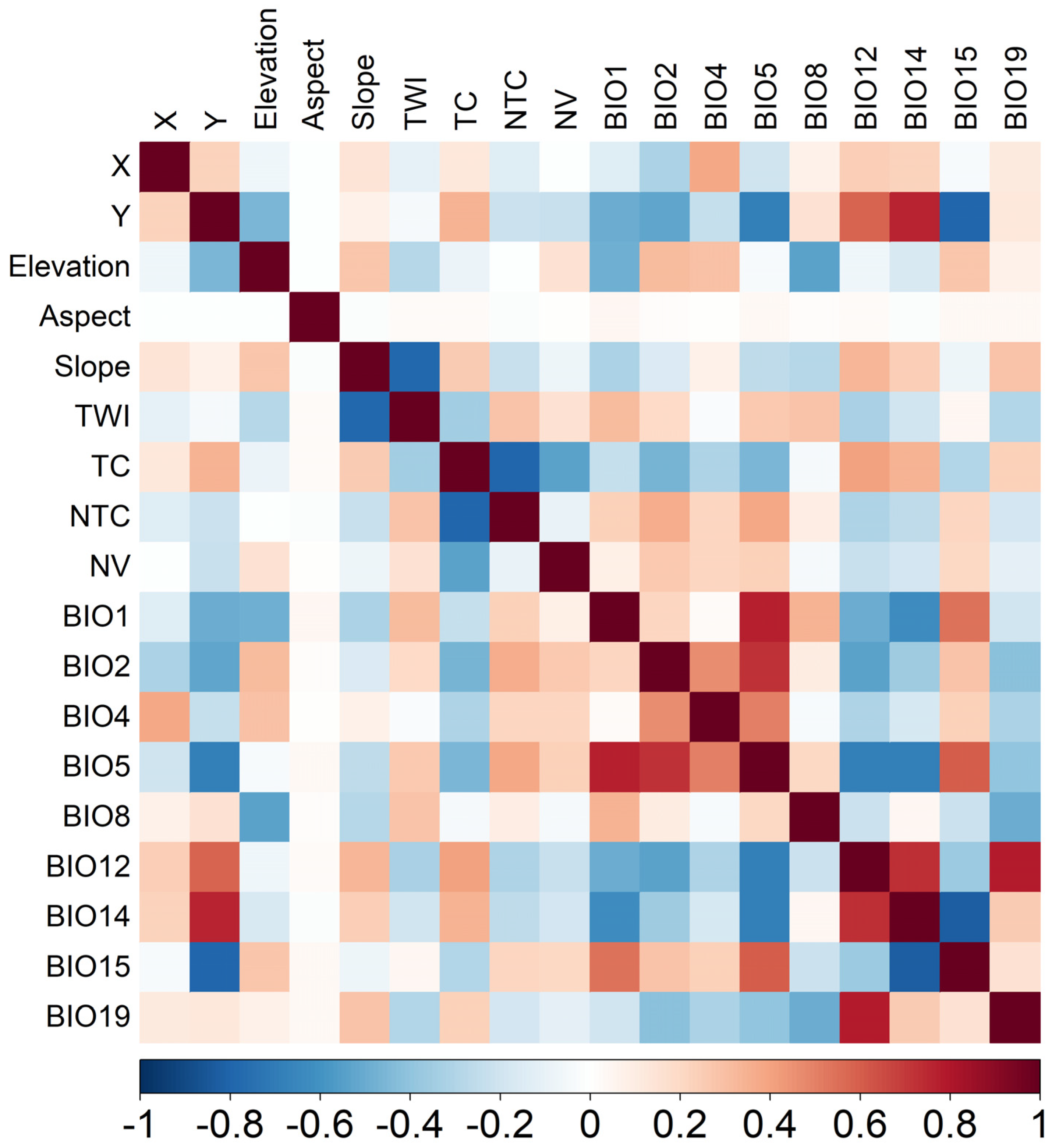
2.3. Variables Selection
2.4. MaxEnt Modeling
2.5. Model Application and Data Analysis
3. Results
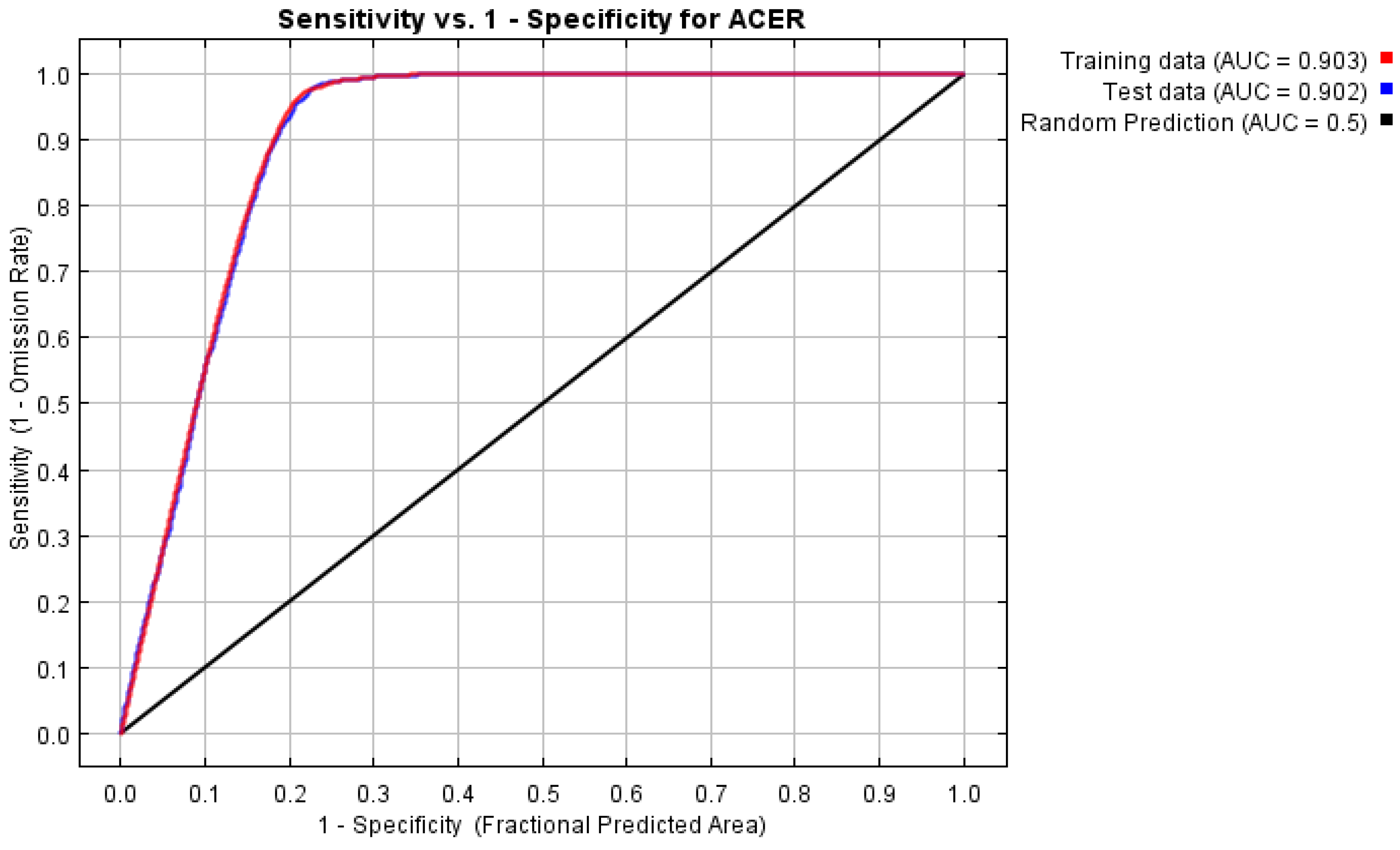
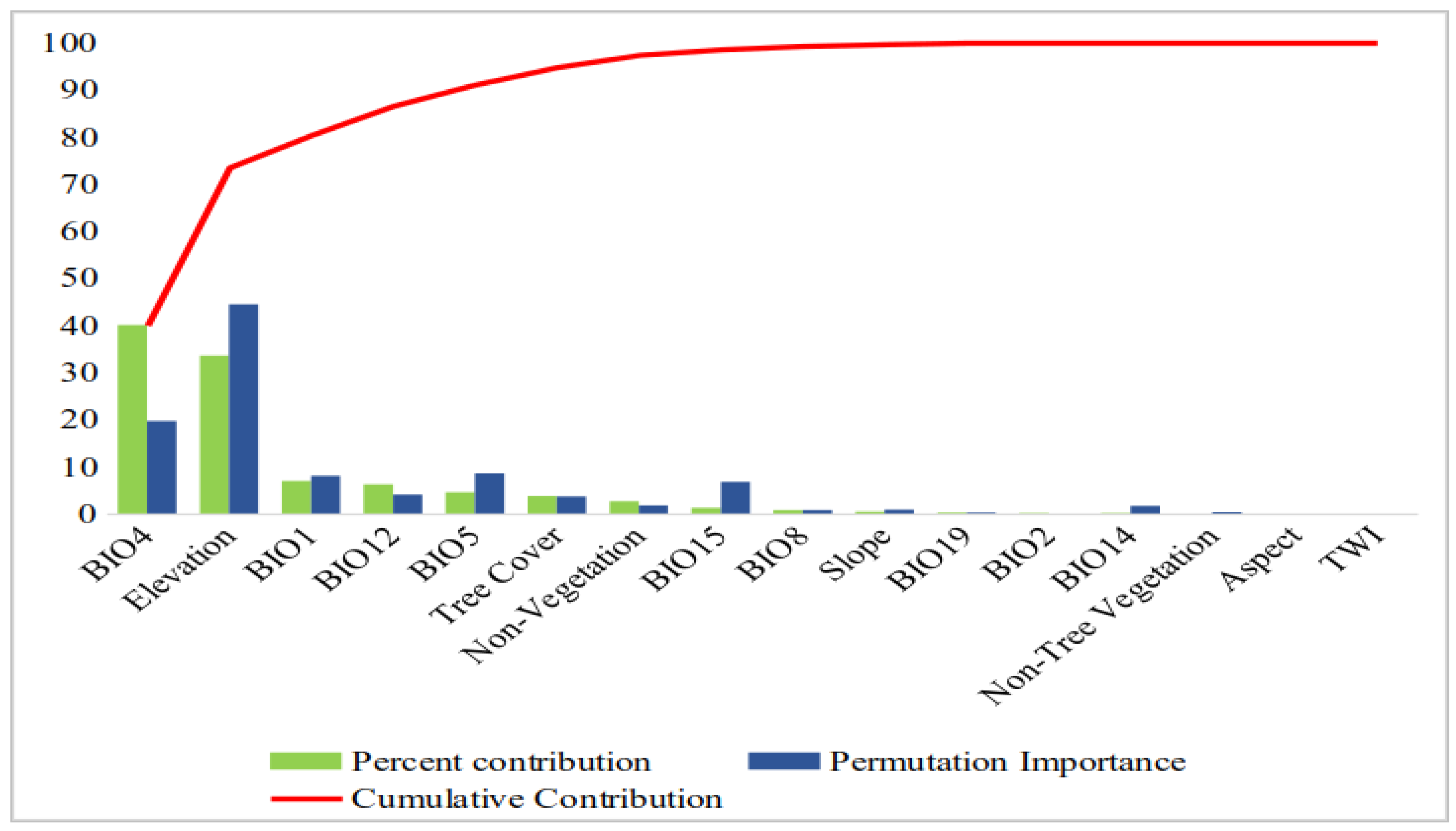
3.1. Model Fitting Results
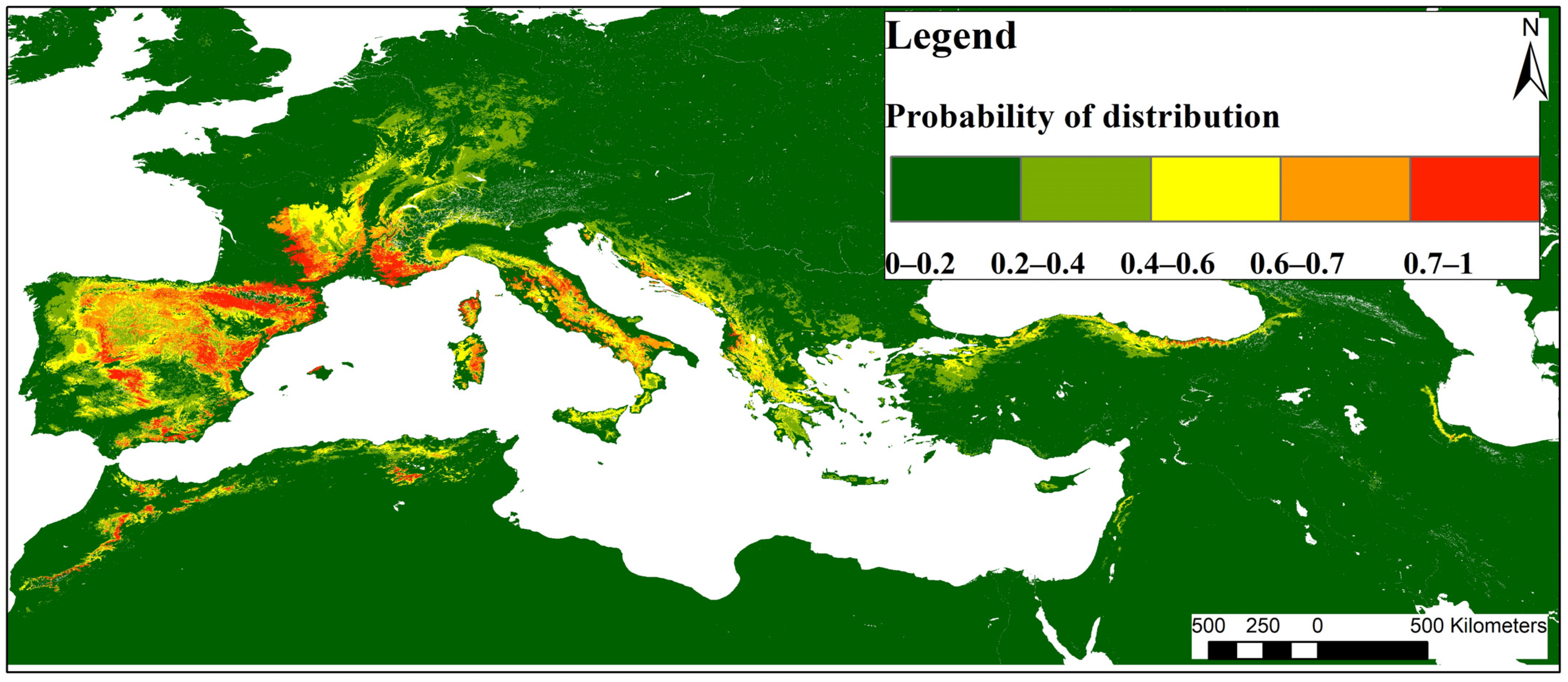
3.2. Current Potential Distribution
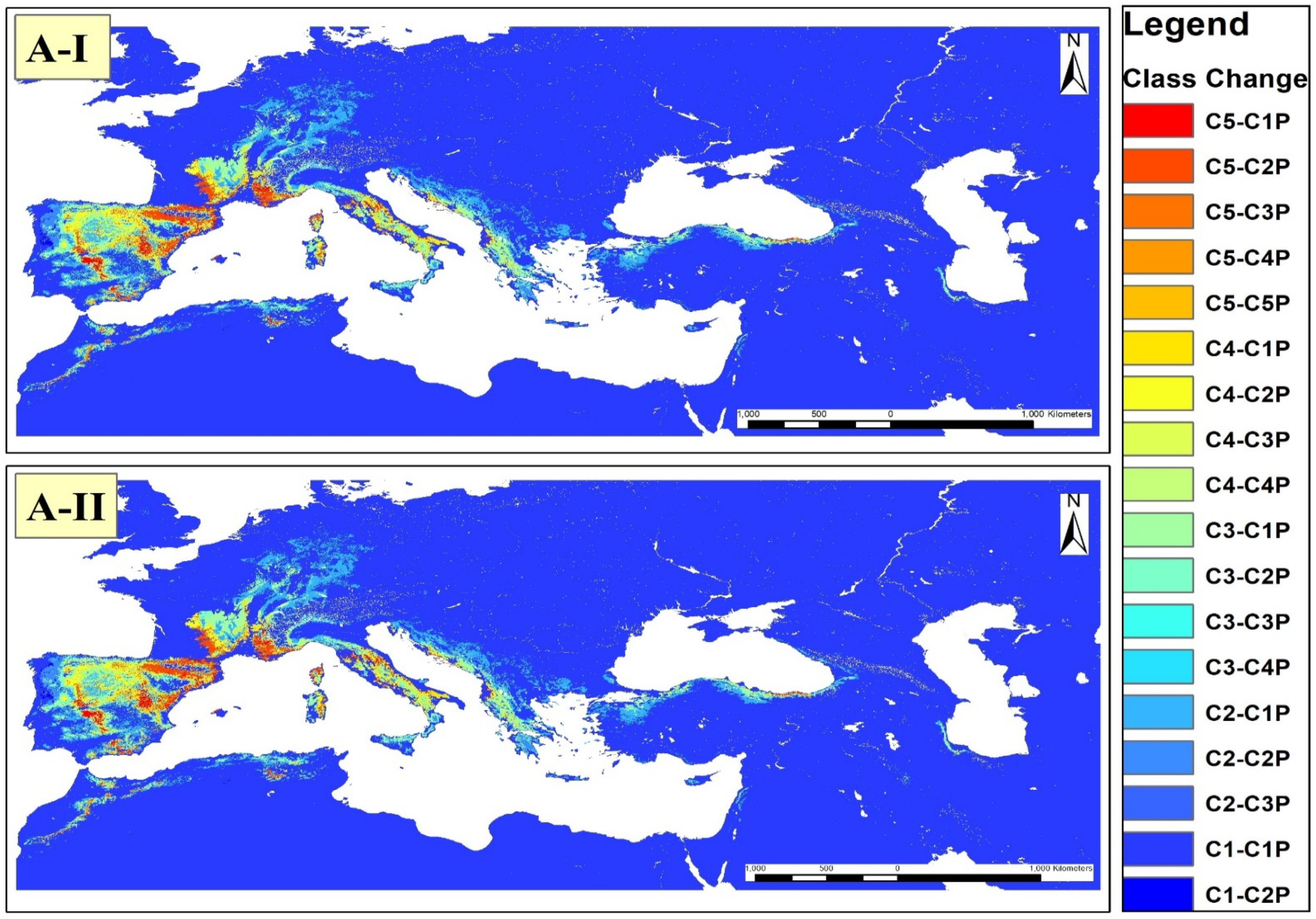
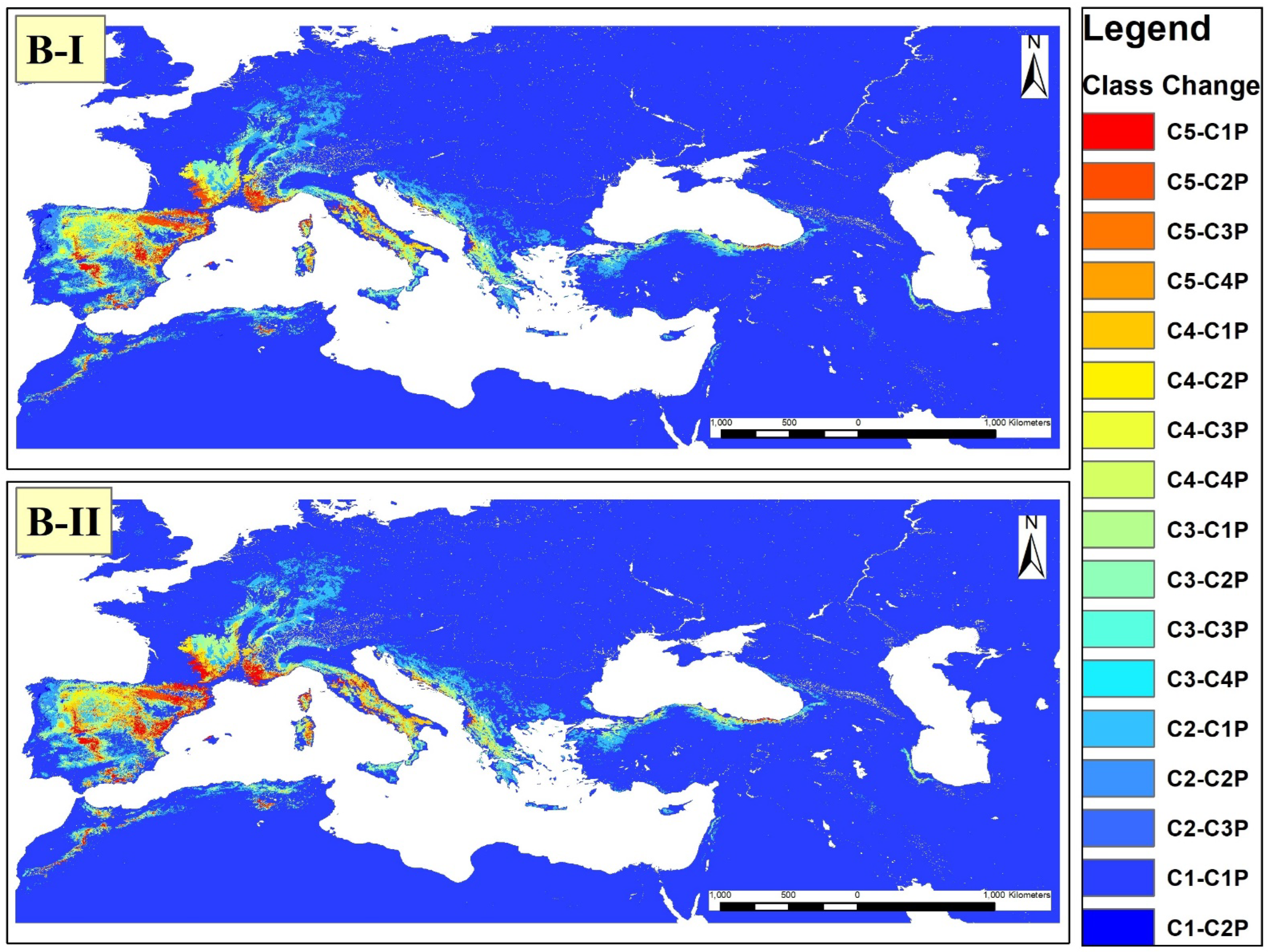
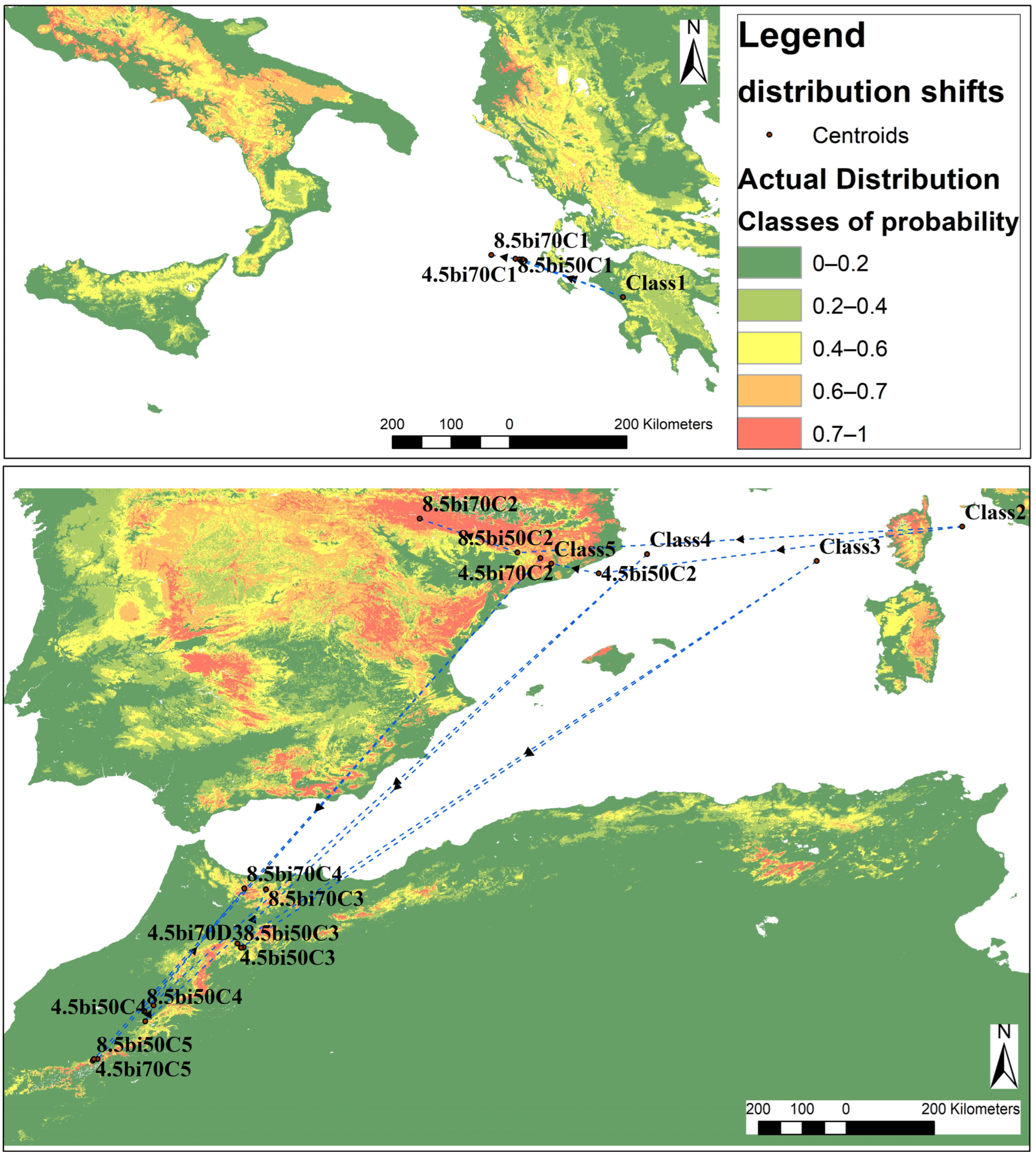
3.3. Suitability Distribution in the Future
4. Discussion
4.1. Current Potential Distribution
4.2. Suitability Distribution in the Future
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Milligan, S.R.; Holt, W.V.; Lloyd, R. Impacts of Climate Change and Environmental Factors on Reproduction and Development in Wildlife. Phil. Trans. R. Soc. B 2009, 364, 3313–3319. [Google Scholar] [CrossRef] [Green Version]
- Mott, C.L. Environmental Constraints to the Geographic Expansion of Plant and Animal Species. Nat. Educ. Knowl. 2010, 3, 72. [Google Scholar]
- Yi, Y.; Cheng, X.; Yang, Z.-F.; Zhang, S.-H. Maxent Modeling for Predicting the Potential Distribution of Endangered Medicinal Plant (H. Riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Çoban, H.O.; Örücü, Ö.K.; Arslan, E.S. MaxEnt Modeling for Predicting the Current and Future Potential Geographical Distribution of Quercus Libani Olivier. Sustainability 2020, 12, 2671. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.S.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.C.; D’Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. The Impacts of Increasing Drought on Forest Dynamics, Structure, and Biodiversity in the United States. Glob. Chang. Biol. 2016, 22, 2329–2352. [Google Scholar] [CrossRef] [Green Version]
- Gazol, A.; Camarero, J.J.; Vicente-Serrano, S.M.; Sánchez-Salguero, R.; Gutiérrez, E.; de Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; Tíscar, P.A.; et al. Forest Resilience to Drought Varies across Biomes. Glob. Chang. Biol. 2018, 24, 2143–2158. [Google Scholar] [CrossRef]
- Okin, G.S.; Dong, C.; Willis, K.S.; Gillespie, T.W.; MacDonald, G.M. The Impact of Drought on Native Southern California Vegetation: Remote Sensing Analysis Using MODIS-Derived Time Series. J. Geophys. Res. Biogeosciences 2018, 123, 1927–1939. [Google Scholar] [CrossRef]
- Rodriguez-Caballero, E.; Belnap, J.; Büdel, B.; Crutzen, P.J.; Andreae, M.O.; Pöschl, U.; Weber, B. Dryland Photoautotrophic Soil Surface Communities Endangered by Global Change. Nat. Geosci. 2018, 11, 185–189. [Google Scholar] [CrossRef]
- Wieneke, S.; Burkart, A.; Cendrero-Mateo, M.P.; Julitta, T.; Rossini, M.; Schickling, A.; Schmidt, M.; Rascher, U. Linking Photosynthesis and Sun-Induced Fluorescence at Sub-Daily to Seasonal Scales. Remote Sens. Environ. 2018, 219, 247–258. [Google Scholar] [CrossRef]
- Guimarães, P.R. The Structure of Ecological Networks Across Levels of Organization. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 433–460. [Google Scholar] [CrossRef]
- Bhat, J.A.; Kumar, M.; Pala, N.A.; Shah, S.; Dayal, S.; Gunathilake, C.; Negi, A.K. Influence of Altitude on the Distribution Pattern of Flora in a Protected Area of Western Himalaya. Acta Ecol. Sin. 2020, 40, 30–43. [Google Scholar] [CrossRef]
- Bazzaz, F.A. The Response of Natural Ecosystems to the Rising Global CO2 Levels. Annu. Rev. Ecol. Syst. 1990, 21, 167–196. [Google Scholar] [CrossRef]
- Malcolm, J.R.; Markham, A.; Neilson, R.P.; Garaci, M. Estimated Migration Rates under Scenarios of Global Climate Change. J. Biogeogr. 2002, 29, 835–849. [Google Scholar] [CrossRef] [Green Version]
- Malcolm, J.R.; Liu, C.; Neilson, R.P.; Hansen, L.; Hannah, L. Global Warming and Extinctions of Endemic Species from Biodiversity Hotspots. Conserv. Biol. 2006, 20, 538–548. [Google Scholar] [CrossRef]
- Dawson, T.P.; Jackson, S.T.; House, J.I.; Prentice, I.C.; Mace, G.M. Beyond Predictions: Biodiversity Conservation in a Changing Climate. Science 2011, 332, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Ducrocq, V.; Drobinski, P.; Lambert, D.; Molinié, G.; Llasat, C. Preface: Forecast and Projection in Climate Scenario of Mediterranean Intense Events: Uncertainties and Propagation on Environment (the MEDUP Project). Nat. Hazards Earth Syst. Sci. 2013, 13, 3043–3047. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.-U.; Seo, K.-H.; Chen, D. Climate Change over the Mediterranean and Current Destruction of Marine Ecosystem. Sci. Rep. 2019, 9, 18813. [Google Scholar] [CrossRef] [Green Version]
- Angert, A.L.; Bontrager, M.G.; Ågren, J. What Do We Really Know About Adaptation at Range Edges? Annu. Rev. Ecol. Evol. Syst. 2020, 51, 341–361. [Google Scholar] [CrossRef]
- Martínez-López, J.; Bagstad, K.J.; Balbi, S.; Magrach, A.; Voigt, B.; Athanasiadis, I.; Pascual, M.; Willcock, S.; Villa, F. Towards Globally Customizable Ecosystem Service Models. Sci. Total Environ. 2019, 650, 2325–2336. [Google Scholar] [CrossRef]
- Vilà-Cabrera, A.; Premoli, A.C.; Jump, A.S. Refining Predictions of Population Decline at Species’ Rear Edges. Glob. Chang. Biol. 2019, 25, 1549–1560. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Matesanz, S.; Guilhaumon, F.; Araújo, M.B.; Balaguer, L.; Benito-Garzón, M.; Cornwell, W.; Gianoli, E.; Kleunen, M.; Naya, D.E.; et al. The Effects of Phenotypic Plasticity and Local Adaptation on Forecasts of Species Range Shifts under Climate Change. Ecol. Lett. 2014, 17, 1351–1364. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stohlgren, T.J. Maxent Modeling for Predicting Suitable Habitat for Threatened and Endangered Tree Canacomyrica Monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 94–98. [Google Scholar]
- Gebrewahid, Y.; Abrehe, S.; Meresa, E.; Eyasu, G.; Abay, K.; Gebreab, G.; Kidanemariam, K.; Adissu, G.; Abreha, G.; Darcha, G. Current and Future Predicting Potential Areas of Oxytenanthera Abyssinica (A. Richard) Using MaxEnt Model under Climate Change in Northern Ethiopia. Ecol. Process. 2020, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Hemati, T.; Pourebrahim, S.; Monavari, M.; Baghvand, A. Species-Specific Nature Conservation Prioritization (a Combination of MaxEnt, Co$ting Nature and DINAMICA EGO Modeling Approaches). Ecol. Model. 2020, 429, 109093. [Google Scholar] [CrossRef]
- Mahatara, D.; Acharya, A.; Dhakal, B.; Sharma, D.; Ulak, S.; Paudel, P. Maxent Modelling for Habitat Suitability of Vulnerable Tree Dalbergia Latifolia in Nepal. Silva Fennica 2021, 55. [Google Scholar] [CrossRef]
- Spatial Modeling in Forest Resources Management; Shit, P.K.; Pourghasemi, H.R.; Das, P.; Bhunia, G.S. (Eds.) Environmental Science and Engineering; Springer International Publishing: Cham, Switzerland, 2021; ISBN 978-3-030-56541-1. [Google Scholar]
- Wei, Y.; Zhang, L.; Wang, J.; Wang, W.; Niyati, N.; Guo, Y.; Wang, X. Chinese Caterpillar Fungus (Ophiocordyceps Sinensis) in China: Current Distribution, Trading, and Futures under Climate Change and Overexploitation. Sci. Total Environ. 2021, 755, 142548. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A Review of Methods for the Assessment of Prediction Errors in Conservation Presence/Absence Models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A Practical Guide to MaxEnt for Modeling Species’ Distributions: What It Does, and Why Inputs and Settings Matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent Modeling for Predicting Impacts of Climate Change on the Potential Distribution of Thuja Sutchuenensis Franch., an Extremely Endangered Conifer from Southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt Modeling to Predict the Potential Distribution of the Endemic Plant Rosa Arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Zhang, K.; Sun, L.; Tao, J. Impact of Climate Change on the Distribution of Euscaphis Japonica (Staphyleaceae) Trees. Forests 2020, 11, 525. [Google Scholar] [CrossRef]
- Du, Z.; He, Y.; Wang, H.; Wang, C.; Duan, Y. Potential Geographical Distribution and Habitat Shift of the Genus Ammopiptanthus in China under Current and Future Climate Change Based on the MaxEnt Model. J. Arid Environ. 2021, 184, 104328. [Google Scholar] [CrossRef]
- Wei, B.; Wang, R.; Hou, K.; Wang, X.; Wu, W. Predicting the Current and Future Cultivation Regions of Carthamus Tinctorius L. Using MaxEnt Model under Climate Change in China. Glob. Ecol. Conserv. 2018, 16, e00477. [Google Scholar] [CrossRef]
- Willcock, S.; Hooftman, D.A.P.; Balbi, S.; Blanchard, R.; Dawson, T.P.; O’Farrell, P.J.; Hickler, T.; Hudson, M.D.; Lindeskog, M.; Martinez-Lopez, J.; et al. A Continental-Scale Validation of Ecosystem Service Models. Ecosystems 2019, 22, 1902–1917. [Google Scholar] [CrossRef] [Green Version]
- IPCC Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Crowley, D.; Rivers, M.C.; Barstow, M. 2018. Acer Monspessulanum (Errata Version Published in 2018). The IUCN Red List of Threatened Species: 2018, e.T193835A135202094. Available online: https://dx.doi.org/10.2305/IUCN.UK.2018-1.RLTS.T193835A124731677.en (accessed on 10 January 2021).
- Lionello, P.; Planton, S.; Rodo, X. Preface: Trends and Climate Change in the Mediterranean Region. Glob. Planet. Chang. 2008, 63, 87–89. [Google Scholar] [CrossRef]
- Lionello, P.; Abrantes, F.; Gacic, M.; Planton, S.; Trigo, R.; Ulbrich, U. The Climate of the Mediterranean Region: Research Progress and Climate Change Impacts. Reg. Environ. Chang. 2014, 14, 1679–1684. [Google Scholar] [CrossRef]
- Planton, S.; Driouech, F.; Rhaz, K.E.; Lionello, P. Sub-Chapter 1.2.2. The Climate of the Mediterranean Regions in the Future Climate Projections. In The Mediterranean Region under Climate Change, IRD ed.; Recherche pour le Développement or IRD: Marseille, France, 2016; pp. 83–91. [Google Scholar]
- DGF Filière Des Semences Forestières et Pastorales En Tunisie. Projet de Gestion Intégrée des Forêts (Phase II); DGF: Tunis, Tunisia, 2015. [Google Scholar]
- Van Gelderen, D.M.; Oterdoom, H.J.; de Jong, P.C. Maples of the World; Timber Press: Portland, OR, USA, 1995; Volume 32. [Google Scholar]
- D’Ambrosio, C. La Biosistematica Molecolare Del Genere Acer: Applicazione Del Método Barcoding Alle Specie Italiane. Ph.D. Thesis, Università degli Studi della Tuscia di Viterbo, Viterbo, Italy, 2009. [Google Scholar]
- Coombes, A.J.; Debreczy, Z. Arbres:L’encyclopedie Des 600 Plus Beaux Arbres Du Monde; Flammarion: Paris, France, 2011. [Google Scholar]
- Zare, A. A Study of Different Treatment Effect on Seed Germination Characteristics and Seedling Survival Montpellier Maple (Acer Menspessolanum Subsp. Turcomanicum Rech. F.). Indian J. Fundam. Appl. Life Sci. 2014, 4, 455–464. [Google Scholar]
- Wieczorek, K.; Kanturski, M.; Junkiert, Ł.; Bugaj-Nawrocka, A. A Comparative Morphometric Study of the Genus Drepanosiphoniella Davatchi, Hille Ris Lambers and Remaudière (Hemiptera: Aphididae: Drepanosiphinae). Zool. Anz. A J. Comp. Zool. 2015, 257, 39–53. [Google Scholar] [CrossRef]
- Pottier-Alapetite, G. Flore De La Tunisie: Angiospermes-Dicotyledones: * Apetales-Dialypetales; Le Ministère de l’Enseignement Supérieur et de la Recherche Scientifique et le Ministère de l’Agriculture: Tunis, Tunisia, 1979. [Google Scholar]
- Bi, W.; Gao, Y.; Shen, J.; He, C.; Liu, H.; Peng, Y.; Zhang, C.; Xiao, P. Traditional Uses, Phytochemistry, and Pharmacology of the Genus Acer (Maple): A Review. J. Ethnopharmacol. 2016, 189, 31–60. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Jia, D.; Tao, J. Species Distribution Modeling of Sassafras Tzumu and Implications for Forest Management. Sustainability 2020, 12, 4132. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- FAO. Harmonized World Soil Database (Version 1.1); FAO: Rome, Italy, 2009. [Google Scholar]
- Kopecký, M.; Macek, M.; Wild, J. Topographic Wetness Index Calculation Guidelines Based on Measured Soil Moisture and Plant Species Composition. Sci. Total Environ. 2021, 757, 143785. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Khan, A.M.; Li, Q.; Saqib, Z.; Khan, N.; Habib, T.; Khalid, N.; Majeed, M.; Tariq, A. MaxEnt Modelling and Impact of Climate Change on Habitat Suitability Variations of Economically Important Chilgoza Pine (Pinus gerardiana Wall.) in South Asia. Forests 2022, 13, 715. [Google Scholar] [CrossRef]
- Jiang, R.; Zou, M.; Qin, Y.; Tan, G.; Huang, S.; Quan, H.; Zhou, J.; Liao, H. Modeling of the potential geographical distribution of three Fritillaria species under climate change. Front. Plant Sci. 2021, 12, 749838. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Swets, J. Measuring the Accuracy of Diagnostic Systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent Modeling for Predicting the Potential Distribution of Medicinal Plant, Justicia Adhatoda L. in Lesser Himalayan Foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Meddour, R.; Meddour-Sahar, O.; Derridj, A.; Géhu, J.-M. Synopsis Commenté Des Groupements Végétaux Forestiers et Préforestiers de La Kabylie Djurdjuréenne (Algérie). Rev. For. Française 2010, 62, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Klausmeyer, K.R.; Shaw, M.R. Climate Change, Habitat Loss, Protected Areas and the Climate Adaptation Potential of Species in Mediterranean Ecosystems Worldwide. PLoS ONE 2009, 4, e6392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, M.; Li, C.; Liu, Z. Optimized Maxent Model Predictions of Climate Change Impacts on the Suitable Distribution of Cunninghamia Lanceolata in China. Forests 2020, 11, 302. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Xiong, Q.; Yu, L.; Yan, W.; Qu, X. Impact of Climate Change on Potential Distribution Patterns of Alpine Vegetation in the Hengduan Mountains Region, China. Mt. Res. Dev. 2020, 40, R48–R54. Available online: https://www.jstor.org/stable/27003481 (accessed on 14 February 2021). [CrossRef]
- Fyllas, N.M.; Koufaki, T.; Sazeides, C.I.; Spyroglou, G.; Theodorou, K. Potential Impacts of Climate Change on the Habitat Suitability of the Dominant Tree Species in Greece. Plants 2022, 11, 1616. [Google Scholar] [CrossRef] [PubMed]
- Cuttelod, A.; Garcia, N.; Malak, D.A.; Temple, H.J.; Katarya, V. The Mediterranean: A Biodiversity Hotspot under Threat. In Wildlife in a Changing World: An Analysis of the 2008 IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2009; Volume 89. [Google Scholar]
- Portilla Cabrera, C.V.; Selvaraj, J.J. Geographic Shifts in the Bioclimatic Suitability for Aedes Aegypti under Climate Change Scenarios in Colombia. Heliyon 2020, 6, e03101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sillero, N.; Barbosa, A.M. Common Mistakes in Ecological Niche Models. Int. J. Geogr. Inf. Sci. 2021, 35, 213–226. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Source and Category | Variable | Description | Unit |
|---|---|---|---|
| FAO global Harmonized World Soil Database Topographic data | Elevation | Elevation | m |
| Slope | Slope | Degree | |
| Aspect | Aspect | Degree | |
| TWI | Topographic Wetness Index | ||
| MODIS Vegetation data | TC | Tree Cover | % |
| NTC | Non-Tree Vegetation | % | |
| NV | Non-Vegetation | % | |
| Worldclim Bioclimatic data | BIO1 | Annual Mean Temperature | °C |
| BIO2 | Mean Diurnal Range (Mean of monthly (max temp–min temp)) | °C | |
| BIO4 | Temperature Seasonality (standard deviation × 100) | ||
| BIO5 | Max Temperature of the Warmest Month | °C | |
| BIO8 | Mean Temperature of the Wettest Quarter | °C | |
| BIO12 | Annual Precipitation | mm | |
| BIO14 | Precipitation of the Driest Month | mm | |
| BIO15 | Precipitation Seasonality (Coefficient of Variation) | mm | |
| BIO19 | Precipitation of Coldest Quarter | mm |
| Habitat Suitability Class | Probability of Distribution | Combined Classes |
|---|---|---|
| Class 1: Unsuitable | [0.0–0.2] | Unsuitable classes |
| Class 2: Barely suitable | [0.2–0.4] | |
| Class 3: Suitable | [0.4–0.6] | Suitable classes |
| Class 4: Highly suitable | [0.6–0.7] | |
| Class 5: Extremely suitable | [0.7–1.0] |
| Mean | Observed Distribution | Mean of Unsuitable Classes | Mean of Suitable Classes | Class 1 | Class 2 | Class 3 | Class 4 | Class 5 |
|---|---|---|---|---|---|---|---|---|
| X | 2.9 ± 6.7 | 19.5 ± 13 | 4.2 ± 2.8 | 28.7 ± 19.3 | 10.2 ± 13.3 | 7.1 ± 12.6 | 3.7 ± 9.54 | 1.7 ± 6.47 |
| Y | 42.6 ± 2.5 | 42 ± 0.6 | 41.7 ± 0.1 | 41.5 ± 8.82 | 42.3 ± 4.62 | 41.6 ± 3.41 | 41.7 ± 2.49 | 41.5 ± 2.72 |
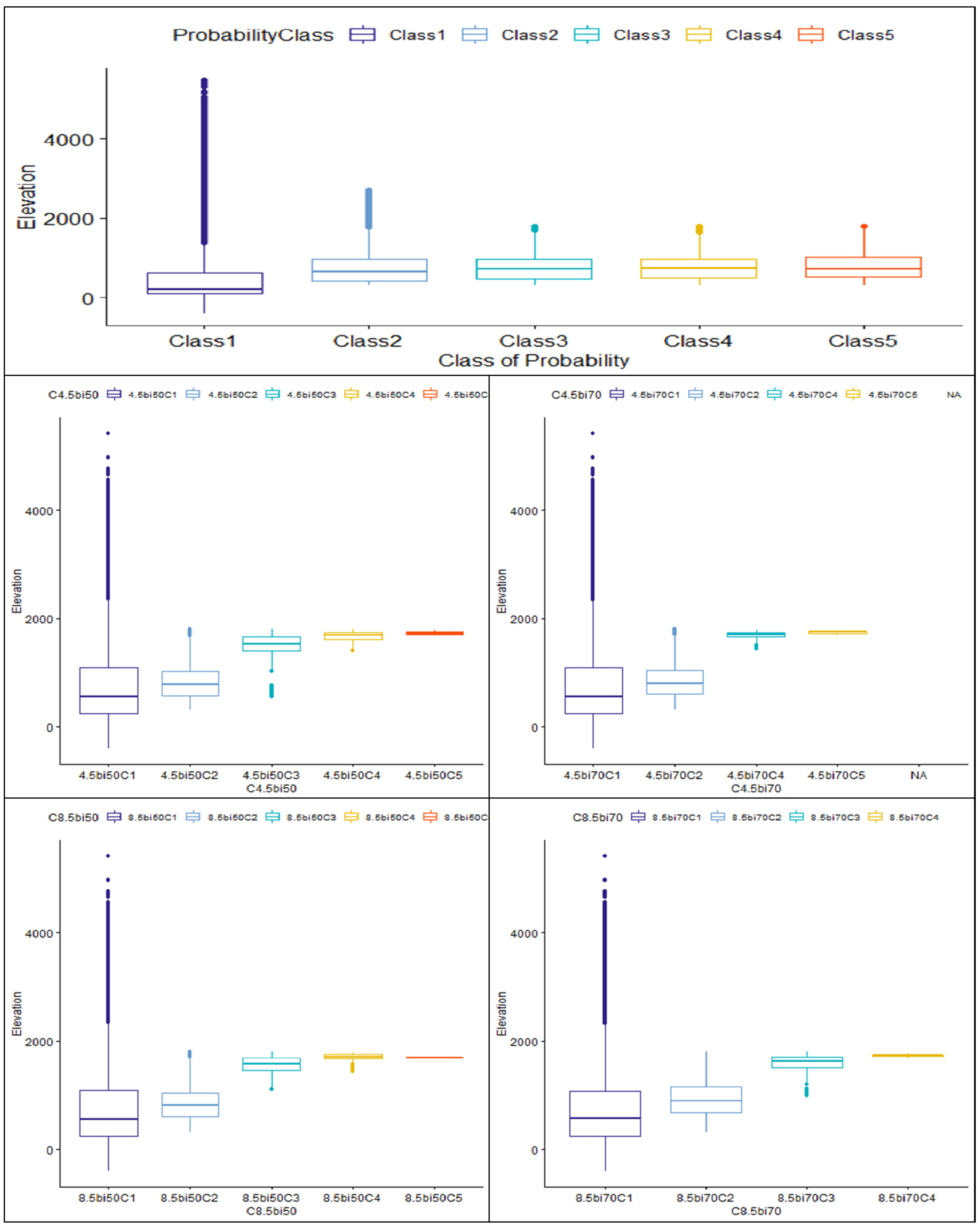
| Elevation | 739.3 ± 327 | 601.1 ± 190.4 | 776.6 ± 19.7 | 466.4 ± 566 | 735.7 ± 355 | 764.5 ± 335 | 766 ± 310 | 799.4 ± 350 |
| Slope | 14.8 ± 22 | 5 ± 4 | 10.7 ± 3.4 | 2.1 ± 10.2 | 7.8 ± 17.7 | 9.7 ± 19.7 | 7.8 ± 17.7 | 14.5 ± 21.9 |
| Aspect | 180.2 ± 103 | 177.6 ± 5.1 | 181.8 ± 2.5 | 173.9 ± 107 | 181.1 ± 108 | 181.6 ± 107 | 179.3 ± 105 | 184.3 ± 101 |
| TWI | 11.6 ± 3.4 | 13.7 ± 1.1 | 12.4 ± 0.8 | 14.4 ± 2.23 | 12.9 ± 3.07 | 12.6 ± 3.27 | 12.9 ± 3.11 | 11.5 ± 3.37 |
| TC | 28.2 ± 18.9 | 18.9 ± 11.6 | 24.7 ± 1.6 | 10.6 ± 17.2 | 27 ± 23.4 | 24.9 ± 21.7 | 23.1 ± 19 | 26.2 ± 18.2 |
| NTV | 55.6 ± 16 | 46.4 ± 9.1 | 58 ± 2 | 39.9 ± 28.8 | 52.7 ± 20.1 | 56 ± 18.3 | 59.9 ± 16.3 | 58 ± 15.1 |
| NV | 16 ± 11.7 | 34.8 ± 20.7 | 17.2 ± 1.7 | 49.4 ± 35 | 20.1 ± 16.4 | 19 ± 13.9 | 16.9 ± 11 | 15.7 ± 10.7 |
| BIO1 | 11.3 ± 1.8 | 11.9 ± 1.3 | 11.6 ± 0.1 | 12.8 ± 6.38 | 10.9 ± 3.05 | 11.5 ± 2.27 | 11.6 ± 1.7 | 11.7 ± 1.63 |
| BIO2 | 10 ± 1.6 | 10.5 ± 0.8 | 10.1 ± 0.3 | 11 ± 2.55 | 9.8 ± 1.81 | 9.8 ± 1.78 | 10 ± 1.79 | 10.4 ± 1.74 |
| BIO4 | 623 ± 56 | 751.3 ± 143.3 | 621.6 ± 7.4 | 852.6 ± 224 | 649.9 ± 65.1 | 629.4 ± 55 | 620.8 ± 44.9 | 614.6 ± 43.9 |
| BIO5 | 26.8 ± 2.6 | 28.5 ± 3 | 27.2 ± 0.3 | 30.6 ± 7.15 | 26.3 ± 4.27 | 26.8 ± 3.47 | 27.1 ± 2.6 | 27.4 ± 2.49 |
| BIO8 | 9.8 ± 3.2 | 11 ± 3.2 | 9.5 ± 0.4 | 13.2 ± 5.49 | 8.7 ± 4.48 | 9.1 ± 3.83 | 9.4 ± 3.18 | 9.9 ± 2.83 |
| BIO12 | 778.4 ± 215 | 627.2 ± 280.7 | 741.3 ± 26.9 | 428.7 ± 314 | 825.7 ± 323 | 772.3 ± 269 | 723.9 ± 222 | 727.6 ± 202 |
| BIO14 | 34.8 ± 18.4 | 26.5 ± 11.9 | 30.8 ± 0.4 | 18.1 ± 19.3 | 34.9 ± 26.1 | 30.4 ± 20.3 | 30.7 ± 17.3 | 31.3 ± 17.7 |
| BIO15 | 28.8 ± 13 | 40.6 ± 7.5 | 32.3 ± 2.6 | 45.8 ± 25.2 | 35.2 ± 19.3 | 35.1 ± 15.1 | 31.7 ± 11.8 | 29.9 ± 12.3 |
| BIO19 | 205.7 ± 70.4 | 180.7 ± 94.5 | 209.2 ± 17.8 | 113.8 ± 89.3 | 247.5 ± 116 | 229.3 ± 99 | 202.9 ± 75.9 | 195.3 ± 61 |
| Suitability Class (%) | History | 45bi50 | 45bi70 | 85bi50 | 85bi70 |
|---|---|---|---|---|---|
| Class 1 % of the total area | 91.80 | 97.27 ± 0.65 | 97.41 ± 0.92 | 97.64 ± 0.81 | 98.24 ± 1.09 |
| Class 2 % of the total area | 3.45 | 2.65 ± 0.63 | 2.51 ± 0.88 | 2.29 ± 0.78 | 1.71 ± 1.08 |
| Class 3 % of the total area | 2.40 | 0.07 ± 0.03 | 0.07 ± 0.05 | 0.06 ± 0.03 | 0.05 ± 0.04 |
| Class 4 % of the total area | 1.47 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.00 ± 0.00 |
| Class 5 % of the total area | 0.89 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Remaining Suitable areas (%) | 0.09 ± 0.02 | 0.08 ± 0.02 | 0.07 ± 0.01 | 0.05 ± 0.01 | |
| Total Suitability loss (%) | 99.91 ± 0.64 | 99.92 ± 0.90 | 99.93 ± 0.00 | 99.95 ± 1.08 | |
| Changes (%) | |||||
| Class 1 to Projected class 1 | 99.91 ± 0.10 | 99.92 ± 0.05 | 99.93 ± 0.05 | 99.85 ± 0.23 | |
| Class 1 to Projected class 2 | 0.09 ± 0.10 | 0.08 ± 0.05 | 0.07 ± 0.05 | 0.14 ± 0.23 | |
| Class 1 to Projected class 3 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Class 1 to Projected class 4 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Class 1 to Projected class 5 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Class 2 to Projected class 1 | 87.39 ± 4.11 | 87.79 ± 5.29 | 88.67 ± 5.05 | 90.13 ± 7.78 | |
| Class 2 to Projected class 2 | 12.43 ± 4.07 | 12.05 ± 5.21 | 11.20 ± 4.99 | 9.70 ± 7.79 | |
| Class 2 to Projected class 3 | 0.18 ± 0.11 | 0.16 ± 0.14 | 0.12 ± 0.11 | 0.17 ± 0.17 | |
| Class 2 to Projected class 4 | 0.00 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Class 2 to Projected class 5 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Class 3 to Projected class 1 | 66.82 ± 8.79 | 67.89 ± 12.30 | 71.43 ± 10.96 | 78.88 ± 12.4 | |
| Class 3 to Projected class 2 | 32.24 ± 8.61 | 31.25 ± 11.94 | 27.87 ± 10.66 | 20.50 ± 12.38 | |
| Class 3 to Projected class 3 | 0.87 ± 0.41 | 0.79 ± 0.49 | 0.64 ± 0.37 | 0.59 ± 0.47 | |
| Class 3 to Projected class 4 | 0.05 ± 0.05 | 0.05 ± 0.04 | 0.05 ± 0.04 | 0.02 ± 0.03 | |
| Class 3 to Projected class 5 | 0.02 ± 0.05 | 0.02 ± 0.04 | 0.01 ± 0.02 | 0.00 ± 0.01 | |
| Class 4 to Projected class 1 | 43.33 ± 13.36 | 46.93 ± 18.85 | 52.04 ± 15.89 | 68.71 ± 18.56 | |
| Class 4 to Projected class 2 | 55.05 ± 12.87 | 51.49 ± 18.20 | 46.74 ± 15.31 | 30.24 ± 18.57 | |
| Class 4 to Projected class 3 | 1.41 ± 0.54 | 1.37 ± 1.31 | 1.04 ± 0.59 | 0.98 ± 1.1 | |
| Class 4 to Projected class 4 | 0.16 ± 0.11 | 0.16 ± 0.09 | 0.13 ± 0.09 | 0.05 ± 0.07 | |
| Class 4 to Projected class 5 | 0.06 ± 0.09 | 0.06 ± 0.08 | 0.05 ± 0.07 | 0.01 ± 0.02 | |
| Class 5 to Projected class 1 | 33.16 ± 12.85 | 37.51 ± 20.56 | 41.85 ± 17.39 | 63.95 ± 23.28 | |
| Class 5 to Projected class 2 | 63.05 ± 11.81 | 58.96 ± 19.14 | 55.21 ± 16.05 | 34.58 ± 22.44 | |
| Class 5 to Projected class 3 | 2.93 ± 0.88 | 2.78 ± 1.68 | 2.31 ± 1.09 | 1.26 ± 1.01 | |
| Class 5 to Projected class 4 | 0.53 ± 0.24 | 0.48 ± 0.25 | 0.40 ± 0.22 | 0.17 ± 0.20 | |
| Class 5 to Projected class 5 | 0.33 ± 0.25 | 0.28 ± 0.27 | 0.23 ± 0.28 | 0.05 ± 0.10 | |
| Loss | |||||
| Gain | |||||
| Remaining | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aouinti, H.; Moutahir, H.; Touhami, I.; Bellot, J.; Khaldi, A. Observed and Predicted Geographic Distribution of Acer monspessulanum L. Using the MaxEnt Model in the Context of Climate Change. Forests 2022, 13, 2049. https://doi.org/10.3390/f13122049
Aouinti H, Moutahir H, Touhami I, Bellot J, Khaldi A. Observed and Predicted Geographic Distribution of Acer monspessulanum L. Using the MaxEnt Model in the Context of Climate Change. Forests. 2022; 13(12):2049. https://doi.org/10.3390/f13122049
Chicago/Turabian StyleAouinti, Hamdi, Hassane Moutahir, Issam Touhami, Juan Bellot, and Abdelhamid Khaldi. 2022. "Observed and Predicted Geographic Distribution of Acer monspessulanum L. Using the MaxEnt Model in the Context of Climate Change" Forests 13, no. 12: 2049. https://doi.org/10.3390/f13122049