
Spatial-Coherent Dynamics and Climatic Signals in the Radial Growth of Siberian Stone Pine (Pinus sibirica Du Tour) in Subalpine Stands along the Western Sayan Mountains

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
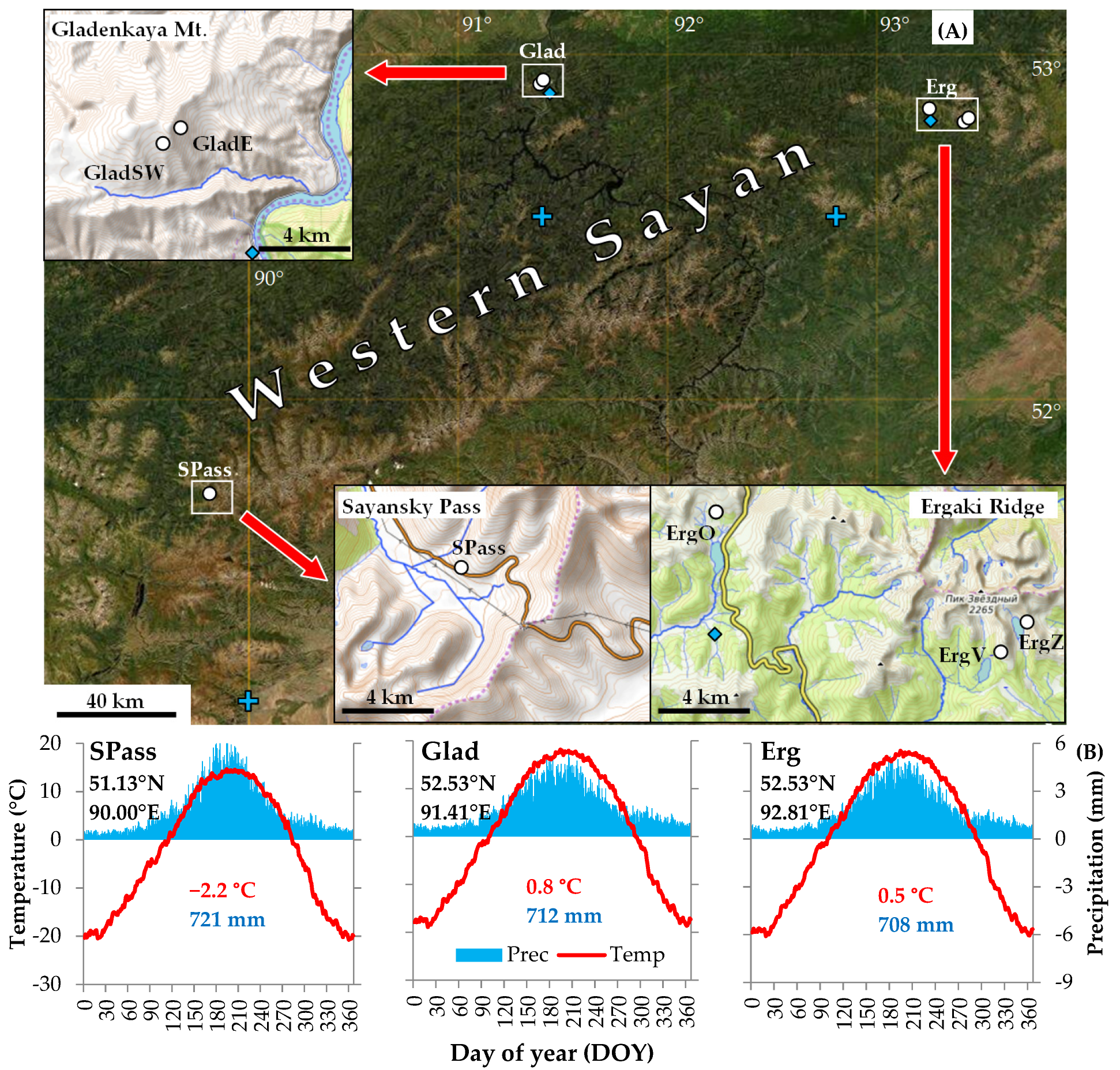
2.1. Study Area and Sampling Sites
2.2. Tree-Ring Width Chronologies and Statistical Analysis
- (i)
- (1—r), where r is a correlation coefficient between two chronologies or their groups (clusters) over the 1936–2010 period (coherence in growth dynamics);
- (ii)
- Euclidean distance, where the coordinates are correlations of chronologies with seasonal climatic variables (coherence in sign and intensity of climatic response);
- (iii)
- Euclidean distance, where the coordinates are the start and end dates of the maximum seasonal climatic responses (coherence in seasonality of climatic response).
3. Results
3.1. TRW Chronologies and Their Spatial Coherence
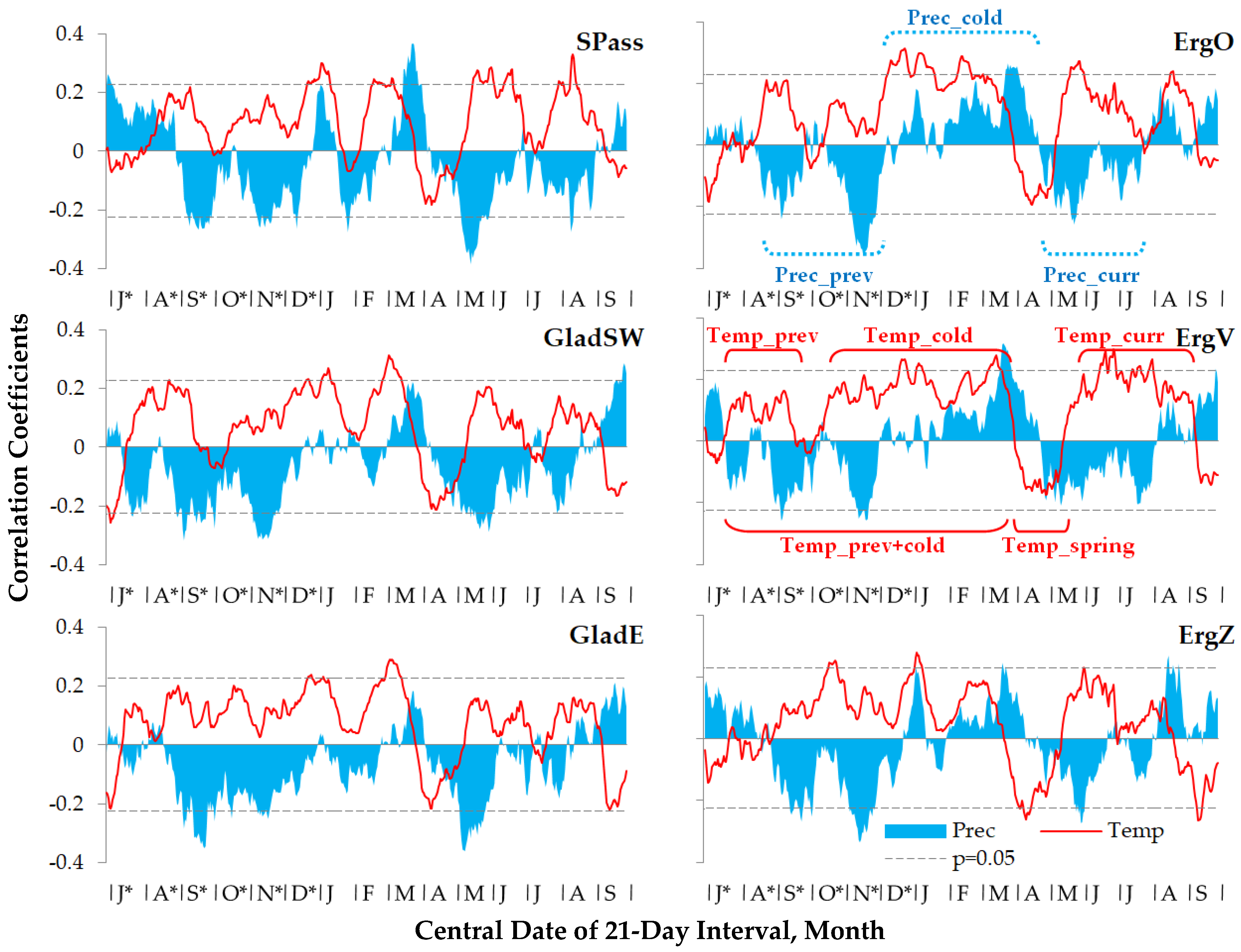
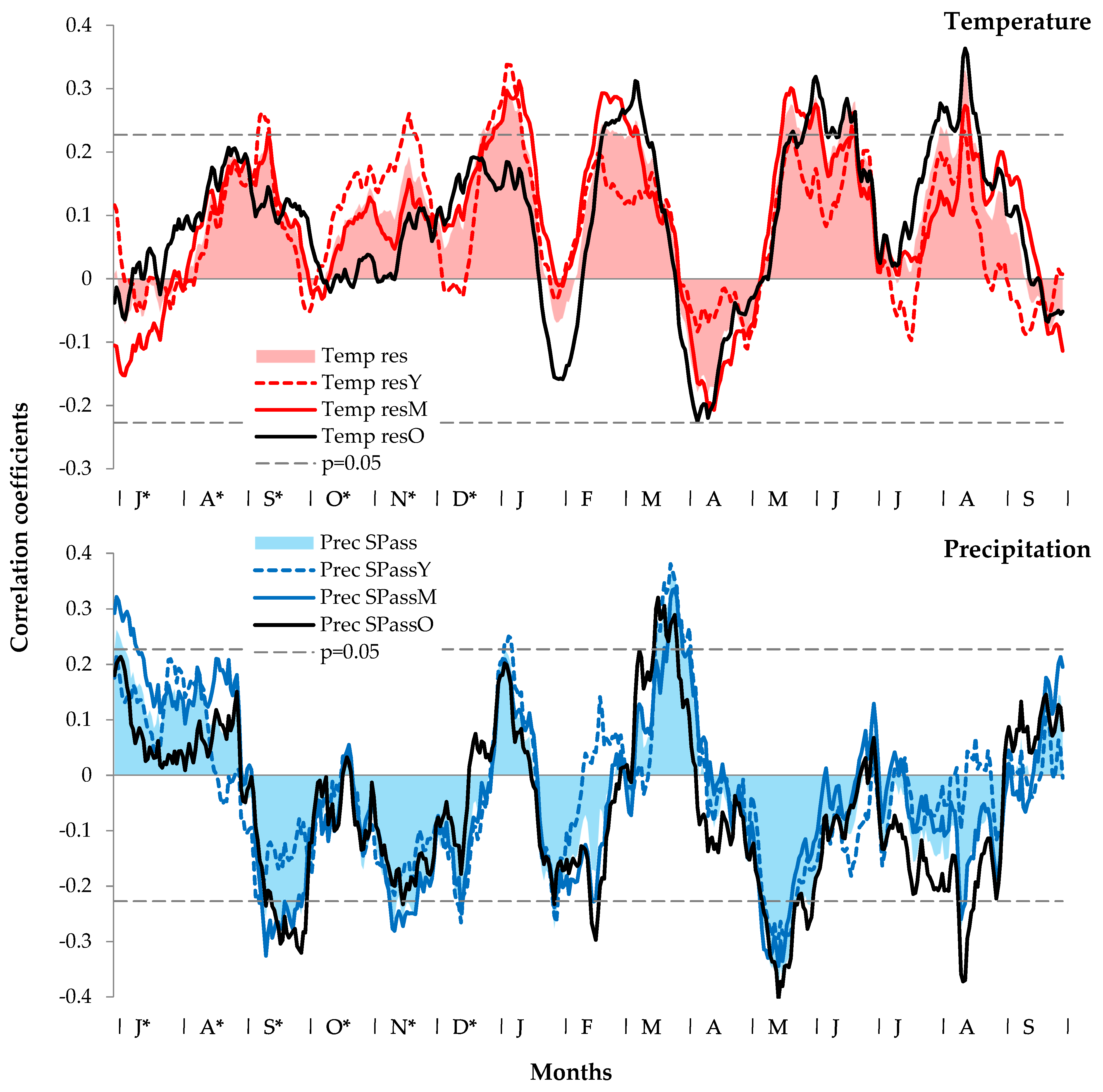
3.2. Climatic Response of Siberian Stone Pine Radial Growth


3.3. Age-Related Patterns of Siberian Stone Pine Growth Dynamics and Its Climatic Response
4. Discussion
4.1. Developed TRW Chronologies, Their Characteristics and Spatial Patterns

4.2. Influence of Climatic Factors on the Radial Growth of Siberian Stone Pine
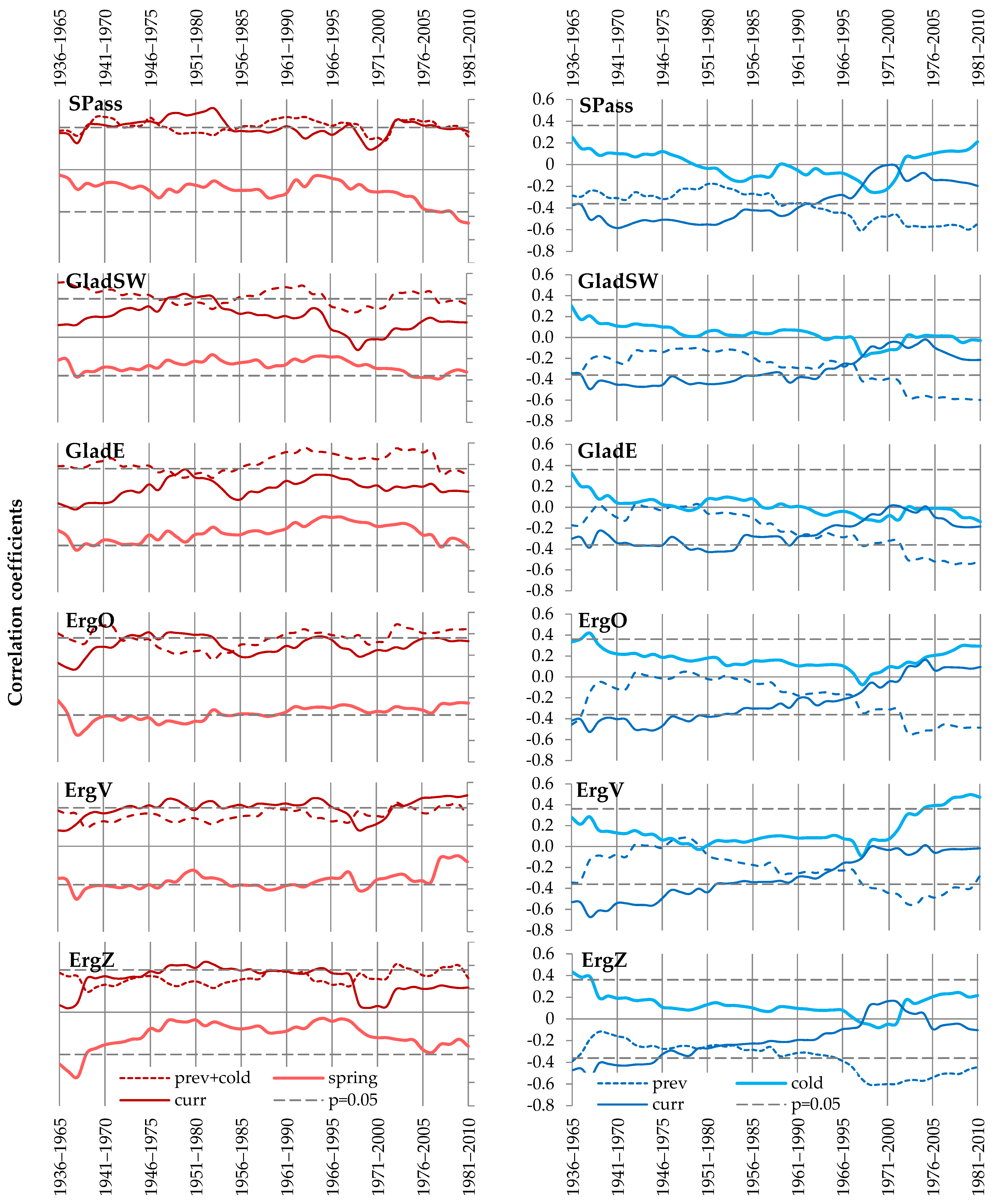
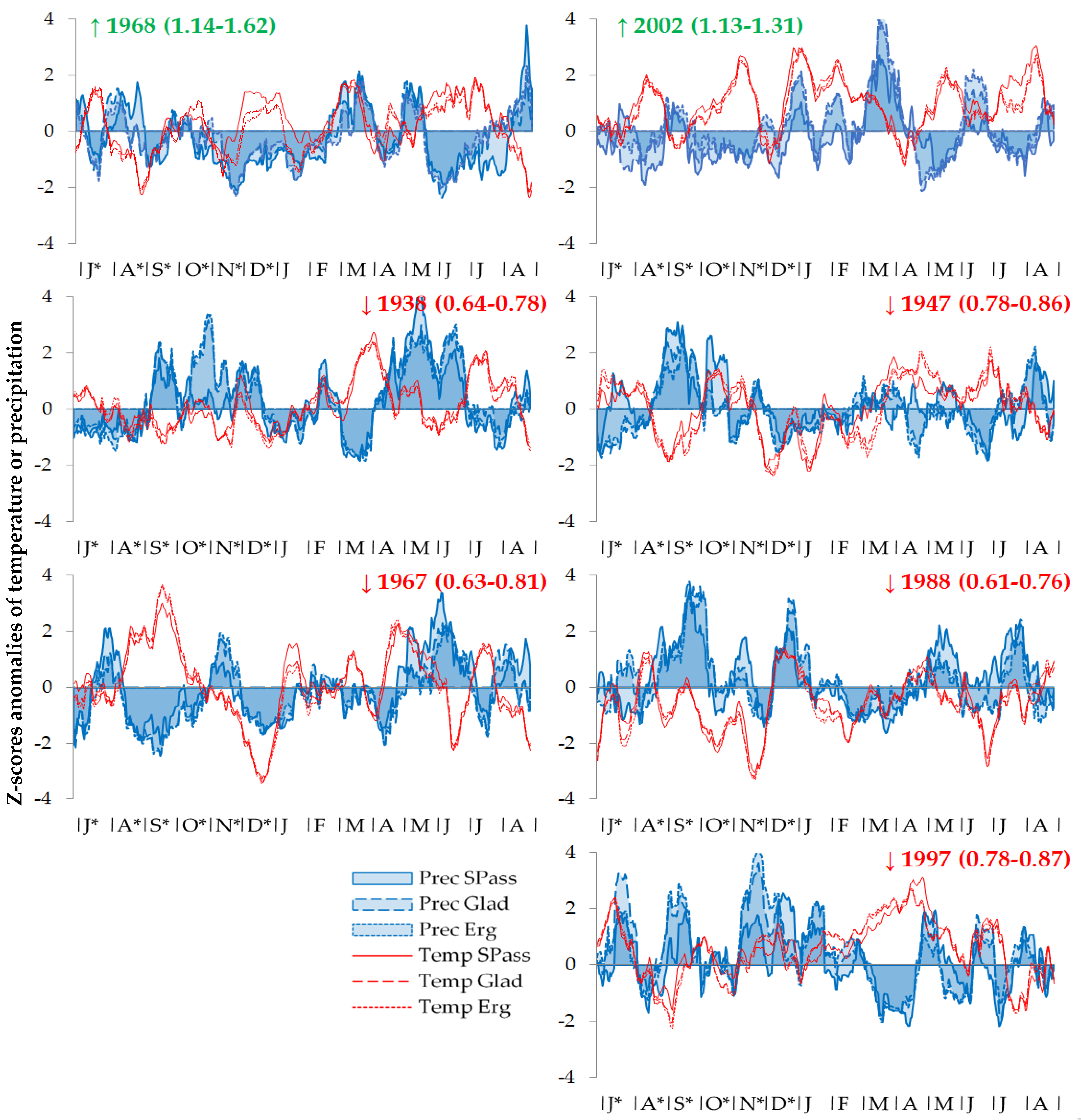
4.3. Spatiotemporal Stability of the Climate Response
4.4. Age-Related Patterns of Climate Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, A.; Cannell, M.G.; Friend, A.D. CO2 stabilization, climate change and the terrestrial carbon sink. Glob. Chang. Biol. 2000, 6, 817–833. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Parilova, T.A.; Kastrikin, V.A.; Bondar’, E.A. Longterm trends in phenological events under influence of climate change: A case study for the Khingansky State Nature Reserve in the Lower Amur Region. In Climate Change Impact on Ecosystems of the Amur River Basin; Darman, Y.A., Kokorin, A.O., Minin, A.A., Eds.; WWF˗Russia: Moscow, Russia, 2006; pp. 47–51. (In Russian) [Google Scholar]
- Kharuk, V.I.; Ranson, K.J.; Im, S.T.; Dvinskaya, M.L. Response of Pinus sibirica and Larix sibirica to climate change in southern Siberian alpine forest–tundra ecotone. Scand. J. For. Res. 2009, 24, 130–139. [Google Scholar] [CrossRef]
- Li, G.Q.; Bai, F.; Sang, W.G. Different responses of radial growth to climate warming in Pinus koraiensis and Picea jezoensis var. komarovii at their upper elevational limits in Changbai Mountain, China. Chin. J. Plant. Ecol. 2011, 35, 500–511. [Google Scholar] [CrossRef]
- Lett, S.; Dorrepaal, E. Global drivers of tree seedling establishment at alpine treelines in a changing climate. Funct. Ecol. 2018, 32, 1666–1680. [Google Scholar] [CrossRef] [Green Version]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Cornona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Parks, C.G.; Bernier, P. Adaptation of forests and forest management to changing climate with emphasis on forest health: A review of science, policies and practices. For. Ecol. Manag. 2010, 259, 657–659. [Google Scholar] [CrossRef]
- Schimel, D.S. Drylands in the earth system. Science 2010, 327, 418–419. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Comm. 2013, 4, 1340. [Google Scholar] [CrossRef] [Green Version]
- Morin, X.; Fahse, L.; Jactel, H.; Scherer-Lorenzen, M.; García-Valdés, R.; Bugmann, H. Long-term response of forest productivity to climate change is mostly driven by change in tree species composition. Sci. Rep. 2018, 8, 5627. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin, Germany, 2003. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Berlin, Germany, 2012. [Google Scholar]
- Bocharov, A.Y. Climatogenetic radial growth of conifers in the upper forest belt of the Seminsky range (the Central Altai Mountains). J. Sib. Fed. Univ. Biol. 2009, 2, 30–37. (In Russian) [Google Scholar]
- Holtmeier, F.K. Mountain Timberlines—Ecology, Patchiness, and Dynamics; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar] [CrossRef] [Green Version]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg (Ted), E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Fadrique, B.; Báez, S.; Duque, Á.; Malizia, A.; Blundo, C.; Carilla, J.; Osinaga-Acosta, O.; Malizia, L.; Silman, M.; Farfán-Ríos, W.; et al. Widespread but heterogeneous responses of Andean forests to climate change. Nature 2018, 564, 207–212. [Google Scholar] [CrossRef]
- McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.; Aukema, B.H.; Bond-Lamberty, B.; Chini, L.; Clark, J.S.; Dietze, M.; Grossiord, C.; Hanbury-Brown, A. Pervasive shifts in forest dynamics in a changing world. Science 2020, 368, eaaz9463. [Google Scholar] [CrossRef]
- Sedelnikov, V.P. High-Mountain Vegetation of the Altai-Sayan Mountains; Nauka Press: Novosibirsk, Russia, 1988. (In Russian) [Google Scholar]
- Chytrý, M.; Danihelka, J.; Kubešová, S.; Lustyk, P.; Ermakov, N.; Hájek, M.; Hájková, P.; Kočí, M.; Otýpková, Z.; Roleček, J.; et al. Diversity of forest vegetation across a strong gradient of climatic continentality: Western Sayan Mountains, southern Siberia. Plant Ecol. 2008, 196, 61–83. [Google Scholar] [CrossRef]
- Danchenko, A.M.; Beh, I.A. Cedar Forests of Western Siberia; Tomsk State University: Tomsk, Russia, 2010. (In Russian) [Google Scholar]
- Nikolaeva, S.A.; Velisevich, S.N.; Savchuk, D.A. Ontogeny of Siberian stone pine (Pinus sibirica Du Tour) in southeastern West Siberian Plain. J. Sib. Fed. Univ. Biol. 2011, 4, 3–22. (In Russian) [Google Scholar]
- Usoltsev, V.A.; Krudyshev, V.V. On the ecology and geography of the Siberian stone pine. Aktualnye Problemy Lesnogo Kompleksa [Actual Probl. For. Complex] 2011, 28, 147–153. (In Russian) [Google Scholar]
- Danchenko, A.M.; Danchenko, M.A.; Myasnikov, A.G.; Bekh, I.A. Kedry Rossii [Stone pines of Russia]; Tomsk State Univ. Publ. House: Tomsk, Russia, 2016. (In Russian) [Google Scholar]
- Tchebakova, N.M.; Parfenova, E.I.; Bazhina, E.V.; Soja, A.J.; Groisman, P.Y. Droughts are not the likely primary cause for Abies sibirica and Pinus sibirica forest dieback in the South Siberian mountains. Forests 2022, 13, 1378. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Tree Rings: Basics and Applications of Dendrochronology; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Frank, D.; Pederson, N.; Cook, E.; Buckley, B.; Nachin, B.; Mijiddorj, R.; Dugarjav, C. 1738 years of Mongolian temperature variability inferred from a tree-ring width chronology of Siberian pine. Geophys. Res. Lett. 2001, 28, 543–546. [Google Scholar] [CrossRef] [Green Version]
- Gerasimova, O.V.; Zharnikov, Z.Y.; Knorre, A.A.; Myglan, V.S. Climatically induced dynamic of radial growth of Siberian stone pine and Siberian fir in the mountain taiga belt in “Ergaki” National Park. J. Sib. Fed. Univ. Biol. 2010, 3, 18–29. (In Russian) [Google Scholar]
- Shah, S.; Yu, J.; Liu, Q.; Shi, J.; Ahmad, A.; Khan, D.; Mannan, A. Climate growth response of Pinus sibirica (Siberian pine) in the Altai mountains, northwestern China. Pak. J. Bot. 2020, 52, 593–600. [Google Scholar] [CrossRef]
- Zonneveld, B.J.M. Conifer genome sizes of 172 species, covering 64 of 67 genera, range from 8 to 72 picogram. Nordic J. Bot. 2012, 30, 490–502. [Google Scholar] [CrossRef]
- MacGillivray, C.W.; Grime, J.P. Genome size predicts frost resistance in British herbaceous plants: Implications for rates of vegetation response to global warming. Funct. Ecol. 1995, 9, 320–325. [Google Scholar] [CrossRef]
- Pellicer, J.; Leitch, I.J. The Plant DNA C-values database (release 7.1): An updated online repository of plant genome size data for comparative studies. New Phytol. 2020, 226, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B. The world-wide ‘fast–slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Ning, Q.R.; Gong, X.W.; Li, M.Y.; Hao, G.Y. Differences in growth pattern and response to climate warming between Larix olgensis and Pinus koraiensis in Northeast China are related to their distinctions in xylem hydraulics. Agric. For. Meteorol. 2022, 312, 108724. [Google Scholar] [CrossRef]
- Wang, H.; Shao, X.M.; Jiang, Y.; Fang, X.Q.; Wu, S.H. The impacts of climate change on the radial growth of Pinus koraiensis along elevations of Changbai Mountain in northeastern China. For. Ecol. Manag. 2013, 289, 333–340. [Google Scholar] [CrossRef]
- Yu, D.; Liu, J.; Lewis, B.J.; Li, Z.; Zhou, W.; Fang, X.; Wei, Y.; Jiang, S.; Dai, L. Spatial variation and temporal instability in the climate–growth relationship of Korean pine in the Changbai Mountain region of Northeast China. For. Ecol. Manag. 2013, 300, 96–105. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, H.; Zhang, Y.; Li, Z.; Wang, X. Climate–growth relationship for different directions of Pinus pumila radial growth at the treeline of northern Daxing’an Mountains, China. Trees 2018, 32, 311–322. [Google Scholar] [CrossRef]
- Cerrato, R.; Salvatore, M.C.; Gunnarson, B.E.; Linderholm, H.W.; Carturan, L.; Brunetti, M.; De Blasi, F.; Baroni, C. A Pinus cembra L. tree-ring record for late spring to late summer temperature in the Rhaetian Alps, Italy. Dendrochronologia 2019, 53, 22–31. [Google Scholar] [CrossRef]
- Brubaker, L.B. Responses of tree populations to climatic change. Vegetatio 1986, 67, 119–130. [Google Scholar] [CrossRef]
- Wieser, G.; Holtmeier, F.K.; Smith, W.K. Treelines in a changing global environment. In Trees in a Changing Environment; Tausz, M., Grulke, N., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 221–263. [Google Scholar] [CrossRef]
- Malanson, G.P. Mixed signals in trends of variance in high-elevation tree ring chronologies. J. Mt. Sci. 2017, 14, 1961–1968. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Herrmann, V.; Rollinson, C.R.; Gonzalez, B.; Gonzalez-Akre, E.B.; Pederson, N.; Alexander, M.R.; Allen, C.D.; Alfaro-Sánchez, R.; Awada, T.; et al. Joint effects of climate, tree size, and year on annual tree growth derived from tree-ring records of ten globally distributed forests. Glob. Chang. Biol. 2022, 28, 245–266. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Age-dependent tree-ring growth responses to climate in Larix decidua and Pinus cembra. Ecology 2004, 85, 730–740. [Google Scholar] [CrossRef]
- Esper, J.; Niederer, R.; Bebi, P.; Frank, D. Climate signal age effects—Evidence from young and old trees in the Swiss Engadin. For. Ecol. Manag. 2008, 255, 3783–3789. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; López, B.C.; Loepfe, L.; Lloret, F. Stand-and tree-level determinants of the drought response of Scots pine radial growth. Oecologia 2012, 168, 877–888. [Google Scholar] [CrossRef]
- Linares, J.C.; Taïqui, L.; Sangüesa-Barreda, G.; Seco, J.I.; Camarero, J.J. Age-related drought sensitivity of Atlas cedar (Cedrus atlantica) in the Moroccan Middle Atlas forests. Dendrochronologia 2013, 31, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Konter, O.; Büntgen, U.; Carrer, M.; Timonen, M.; Esper, J. Climate signal age effects in boreal tree-rings: Lessons to be learned for paleoclimatic reconstructions. Quat. Sci. Rev. 2016, 142, 164–172. [Google Scholar] [CrossRef]
- Pompa-García, M.; Hadad, M.A. Sensitivity of pines in Mexico to temperature varies with age. Atmósfera 2016, 29, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Jiao, L.; Jiang, Y.; Wang, M.; Zhang, W.; Zhang, Y. Age-effect radial growth responses of Picea schrenkiana to climate change in the eastern Tianshan Mountains, Northwest China. Forests 2017, 8, 294. [Google Scholar] [CrossRef] [Green Version]
- Van Wijk, M.T.; Clemmensen, K.E.; Shaver, G.R.; Williams, M.; Callaghan, T.V.; Chapin, F.S., III; Cornelissen, J.H.C.; Gough, L.; Hobbie, S.E.; Jonasson, S.; et al. Long-term ecosystem level experiments at Toolik Lake, Alaska, and at Abisko, Northern Sweden: Generalizations and differences in ecosystem and plant type responses to global change. Glob. Chang. Biol. 2003, 10, 105–123. [Google Scholar] [CrossRef]
- Sullivan, P.F.; Ellison, S.B.Z.; McNown, R.W.; Brownlee, A.H.; Sveinbjörnsson, B. Evidence of soil nutrient availability as the proximate constraint on growth of treeline trees in northwest Alaska. Ecology 2015, 96, 716–727. [Google Scholar] [CrossRef]
- Moyes, A.B.; Germino, M.J.; Kueppers, L.M. Moisture rivals temperature in limiting photosynthesis by trees establishing beyond their cold-edge range limit under ambient and warmed conditions. New Phytol. 2015, 207, 1005–1014. [Google Scholar] [CrossRef]
- Oberhuber, W. Influence of climate on radial growth of Pinus cembra within the alpine timberline ecotone. Tree Physiol. 2004, 24, 291–301. [Google Scholar] [CrossRef]
- Vorobev, V.N. (Ed.) Pine Forests: Utilization of Its Products; CRC Press: Boca Raton, CA, USA, 2006. [Google Scholar]
- Hallinger, M.; Manthey, M.; Wilmking, M. Establishing a missing link: Warm summers and winter snow cover promote shrub expansion into alpine tundra in Scandinavia. New Phytol. 2010, 186, 890–899. [Google Scholar] [CrossRef]
- Kirdyanov, A.V.; Hagedorn, F.; Knorre, A.A.; Fedotova, E.V.; Vaganov, E.A.; Naurzbaev, M.M.; Moiseev, P.A.; Rigling, A. 20th century tree-line advance and vegetation changes along an altitudinal transect in the Putorana Mountains, northern Siberia. Boreas 2012, 41, 56–67. [Google Scholar] [CrossRef]
- Hagedorn, F.; Shiyatov, S.G.; Mazepa, V.S.; Devi, N.M.; Grigor’ev, A.A.; Bartish, A.A.; Fomin, V.V.; Kapralov, D.S.; Moiseev, P.A. Treeline advances along the Urals mountain range—Driven by improved winter conditions? Glob. Chang. Biol. 2014, 20, 3530–3543. [Google Scholar] [CrossRef]
- Dan Moore, R.; Spittlehouse, D.; Story, A. Riparian microclimate and stream temperature response to forest harvesting: A review. Am. Water Resour. Assoc. 2005, 41, 813–834. [Google Scholar] [CrossRef]
- Monnier, Y.; Prévosto, B.; Ripert, C.; Corbani, A.C.; Fernandez, C. Forest microhabitats differentially influence seedling phenology of two co-existing Mediterranean oak species. J. Veg. Sci. 2012, 23, 260–270. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G. Treelines—Approaches at different scales. Sustainability 2017, 9, 808. [Google Scholar] [CrossRef] [Green Version]
- Albrich, K.; Rammer, W.; Seidl, R. Climate change causes critical transitions and irreversible alterations of mountain forests. Glob. Chang. Biol. 2020, 26, 4013–4027. [Google Scholar] [CrossRef]
- Harvey, J.E.; Smiljanić, M.; Scharnweber, T.; Buras, A.; Cedro, A.; Cruz-García, R.; Drobyshev, I.; Janecka, K.; Jansons, Ā.; Kaczka, R.; et al. Tree growth influenced by warming winter climate and summer moisture availability in northern temperate forests. Glob. Chang. Biol. 2020, 26, 2505–2518. [Google Scholar] [CrossRef]
- Christiansen, B.; Ljungqvist, F.C. Challenges and perspectives for large-scale temperature reconstructions of the past two millennia. Rev. Geophys. 2017, 55, 40–96. [Google Scholar] [CrossRef]
- Esper, J.; George, S.S.; Anchukaitis, K.; D’Arrigo, R.; Ljungqvist, F.C.; Luterbacher, J.; Schneider, L.; Stoffel, M.; Wilson, R.; Büntgen, U. Large-scale, millennial-length temperature reconstructions from tree-rings. Dendrochronologia 2018, 50, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Alisov, B.P. Climate of the USSR; Moscow State University: Moscow, Russia, 1956. (In Russian) [Google Scholar]
- Compo, G.P.; Whitaker, J.S.; Sardeshmukh, P.D.; Matsui, N.; Allan, R.J.; Yin, X.; Gleason, B.E.; Vose, R.S.; Rutledge, G.; Bessemoulin, P.; et al. The Twentieth Century Reanalysis Project. Q. J. R. Meteorol. Soc. 2011, 137, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Groisman, P.Y.; Rankova, E.Y. Precipitation trends over the Russian permafrost-free zone: Removing the artifacts of pre-processing. Int. J. Climatol. J. R. Meteorol. Soc. 2001, 21, 657–678. [Google Scholar] [CrossRef]
- Polikarpov, N.P.; Nazimova, D.I. The dark coniferous forests of the northern part of the west Siberian mountains. In Forestry Research in the Forests of Siberia; Institute for Forests and Wood: Krasnoyarsk, Russia, 1963; pp. 103–147. (In Russian) [Google Scholar]
- Monserud, R.A.; Tchebakova, N.M. A vegetation model for the Sayan Mountains, southern Siberia. Can. J. For. Res. 1996, 26, 1055–1068. [Google Scholar] [CrossRef]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 68–78. [Google Scholar]
- Rinn, F. TSAP-Win: Time Series Analysis and Presentation for Dendrochronology and Related Applications: User Reference; RINNTECH: Heidelberg, Germany, 2003. [Google Scholar]
- Cook, E.R.; Krusic, P.J. Program ARSTAN: A Tree-ring Standardization Program Based on Detrending and Autoregressive Time Series Modeling, with Interactive Graphics; Lamont-Doherty Earth Observatory, Columbia University: Palisades, LA, USA, 2005. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Appl. Meteorol. Climatol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Griçar, J.; Seo, J.W.; Rathgeber, C.B.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Wilks, D.S. Statistical Methods in the Atmospheric Sciences, 4th ed.; Elsevier: Cambridge, UK, 2019. [Google Scholar]
- Vittoz, P.; Rulence, B.; Largey, T.; Freléchoux, F. Effects of climate and land-use change on the establishment and growth of Cembran pine (Pinus cembra L.) over the altitudinal treeline ecotone in the Central Swiss Alps. Arct. Antarct. Alp. Res. 2008, 40, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Petrov, I.A.; Shushpanov, A.S.; Golyukov, A.S.; Kharuk, V.I. Pinus sibirica Du Tour response to climate change in the forests of the Kuznetsk Alatau Mountains. Sibirskij Lesnoj Zurnal [Sib. J. For. Sci.] 2019, 5, 43–53. [Google Scholar]
- Treml, V.; Ponocná, T.; Büntgen, U. Growth trends and temperature responses of treeline Norway spruce in the Czech-Polish Sudetes Mountains. Clim. Res. 2012, 55, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Chhetri, P.K.; Cairns, D.M. Dendroclimatic response of Abies spectabilis at treeline ecotone of Barun Valley, eastern Nepal Himalaya. J. For. Res. 2016, 27, 1163–1170. [Google Scholar] [CrossRef]
- Wang, T.; Ren, H.; Ma, K. Climatic signals in tree ring of Picea schrenkiana along an altitudinal gradient in the central Tianshan Mountains, northwestern China. Trees 2005, 19, 736–742. [Google Scholar] [CrossRef]
- De Grandpré, L.; Tardif, J.C.; Hessl, A.; Pederson, N.; Conciatori, F.; Green, T.R.; Oyunsanaa, B.; Baatarbileg, N. Seasonal shift in the climate responses of Pinus sibirica, Pinus sylvestris and Larix sibirica trees from semi-arid, north-central Mongolia. Can. J. For. Res. 2011, 41, 1242–1255. [Google Scholar] [CrossRef]
- Timoshok, E.E.; Timoshok, E.N.; Skorokhodov, S.N. Ecology of Siberian stone pine (Pinus sibirica Du Tour) and Siberian larch (Larix sibirica Ledeb.) in the Altai mountain glacial basins. Russ. J. Ecol. 2014, 45, 194–200. [Google Scholar] [CrossRef]
- Nazarov, A.N.; Myglan, V.S. The possibility of construction of the 6000-year chronology for Siberian pine in the Central Altai. J. Sib. Fed. Univ. Biol. 2012, 5, 70–88. (In Russian) [Google Scholar]
- Rolland, C. Decreasing teleconnections with inter-site distance in monthly climatic data and tree-ring width networks in a mountainous Alpine area. Theor. Appl. Climatol. 2002, 71, 63–75. [Google Scholar] [CrossRef]
- Frank, D.; Esper, J. Characterization and climate response patterns of a high-elevation, multi-species tree-ring network in the European Alps. Dendrochronologia 2005, 22, 107–121. [Google Scholar] [CrossRef]
- Li, M.H.; Yang, J.; Kräuchi, N. Growth responses of Picea abies and Larix decidua to elevation in subalpine areas of Tyrol, Austria. Can. J. For. Res. 2003, 33, 653–662. [Google Scholar] [CrossRef]
- Danek, M.; Chuchro, M.; Walanus, A. Tree-ring growth of Larch (Larix decidua Mill.) in the Polish Sudetes—The influence of altitude and site-related factors on the climate-growth relationship. Forests 2018, 9, 663. [Google Scholar] [CrossRef] [Green Version]
- Carnicer, J.; Vives-Ingla, M.; Blanquer, L.; Méndez-Camps, X.; Rosell, C.; Sabaté, S.; Gutiérrez, E.; Sauras, T.; Peñuelas, J.; Barbeta, A. Forest resilience to global warming is strongly modulated by local-scale topographic, microclimatic and biotic conditions. J. Ecol. 2021, 109, 3322–3339. [Google Scholar] [CrossRef]
- Jacoby, G.C.; D’Arrigo, R.D.; Davaajamts, T. Mongolian tree rings and 20th-century warming. Science 1996, 273, 771–773. [Google Scholar] [CrossRef]
- Shi, P.; Körner, C.; Hoch, G. A test of the growth-limitation theory for alpine tree line formation in evergreen and deciduous taxa of the eastern Himalayas. Funct. Ecol. 2008, 22, 213–220. [Google Scholar] [CrossRef]
- Lenz, A.; Hoch, G.; Körner, C. Early season temperature controls cambial activity and total tree ring width at the alpine treeline. Plant Ecol. Divers. 2013, 6, 365–375. [Google Scholar] [CrossRef]
- Lange, O.L.; Schulze, E.D. Untersuchungen über die Dickenentwicklung der kutikularen Zellwandschichten bei der Fichtennadel. Forstwissenschaftliches Centralblatt 1966, 85, 27–38. (In Germany) [Google Scholar] [CrossRef]
- Tranquillini, W. Physiological Ecology of the Alpine Timberline. Tree Existence in High Altitudes with Special Reference to the European Alps; Springer: Berlin, Germany, 1979. [Google Scholar]
- Mayr, S. Limits in water relations. In Trees at their Upper Limit; Wieser, G., Tausz, M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 145–162. [Google Scholar]
- Mayr, S.; Hacke, U.; Schmid, P.; Schwienbacher, F.; Gruber, A. Frost drought in conifers at the alpine timberline: Xylem dysfunction and adaptations. Ecology 2006, 87, 3175–3185. [Google Scholar] [CrossRef] [Green Version]
- Man, R.; Kayahara, G.J.; Dang, Q.L.; Rice, J.A. A case of severe frost damage prior to budbreak in young conifers in Northeastern Ontario: Consequence of climate change? For. Chron. 2009, 85, 453–462. [Google Scholar] [CrossRef]
- Yin, D.; Xu, D.; Tian, K.; Xiao, D.; Zhang, W.; Sun, D.; Sun, H.; Zhang, Y. Radial growth response of Abies georgei to climate at the upper timberlines in central Hengduan Mountains, Southwestern China. Forests 2018, 9, 606. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Jiang, S.; Tardif, J.C.; Liang, H.; Zhang, S.; Li, J.; Yu, B.; Bergeron, Y.; Rossi, S.; Wang, Z. Radial growth responses of two dominant conifers to climate in the Altai Mountains, Central Asia. Agric. For. Meteorol. 2021, 298–299, 108297. [Google Scholar] [CrossRef]
- Wason, J.W.; Beier, C.M.; Battles, J.J.; Dovciak, M. Acidic deposition and climate warming as drivers of tree growth in high-elevation spruce-fir forests of the Northeastern US. Front. For. Glob. Chang. 2019, 2, 63. [Google Scholar] [CrossRef] [Green Version]
- Roitto, M.; Sutinen, S.; Wang, A.F.; Domisch, T.; Lehto, T.; Repo, T. Waterlogging and soil freezing during dormancy affected root and shoot phenology and growth of Scots pine saplings. Tree Physiol. 2019, 39, 805–818. [Google Scholar] [CrossRef]
- Sutinen, M.L.; Holappa, T.; Ritari, A.; Kujala, K. Seasonal changes in soil temperature and snow-cover under different simulated winter conditions: Comparison with frost hardiness of Scots pine (Pinus sylvestris) roots. Chemos.-Glob. Chang. Sci. 1999, 1, 485–492. [Google Scholar] [CrossRef]
- Kirdyanov, A.; Hughes, M.; Vaganov, E.; Schweingruber, F.; Silkin, P. The importance of early summer temperature and date of snow melt for tree growth in the Siberian Subarctic. Trees 2003, 17, 61–69. [Google Scholar] [CrossRef]
- Repo, T.; Lehto, T.; Finér, L. Delayed soil thawing affects root and shoot functioning and growth in Scots pine. Tree Physiol. 2008, 28, 1583–1591. [Google Scholar] [CrossRef] [Green Version]
- Sanmiguel-Vallelado, A.; Camarero, J.J.; Morán-Tejeda, E.; Gazol, A.; Colangelo, M.; Alonso-González, E.; Lopez-Moreno, J.I. Snow dynamics influence tree growth by controlling soil temperature in mountain pine forests. Agric. For. Meteorol. 2021, 296, 108205. [Google Scholar] [CrossRef]
- Pomeroy, J.W.; Marsh, P.; Gray, D.M. Application of a distributed blowing snow model to the Arctic. Hydrol. Process. 1997, 11, 1451–1464. [Google Scholar] [CrossRef]
- Cosma, S.; Richard, E.; Miniscloux, F. The role of small-scale orographic features in the spatial distribution of precipitation. Q. J. R. Meteorol. Soc. 2002, 128, 75–92. [Google Scholar] [CrossRef]
- Konrad, C.E. Maximum precipitation rates in the southern Blue Ridge Mountains of the southeastern United States. Clim. Res. 1995, 5, 159–166. [Google Scholar] [CrossRef]
- Kurita, N. Modern isotope climatology of Russia: A first assessment. J. Geophys. Res. 2004, 109. [Google Scholar] [CrossRef]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Ren, P.; Rossi, S.; Camarero, J.J.; Ellison, A.M.; Liang, E.; Peñuelas, J. Critical temperature and precipitation thresholds for the onset of xylogenesis of Juniperus przewalskii in a semi-arid area of the north-eastern Tibetan Plateau. Ann. Bot. 2018, 121, 617–624. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Manzanedo, R.D.; Zhang, F.; Pederson, N. Cambial phenology and xylogenesis of Juniperus przewalskii over a climatic gradient is influenced by both temperature and drought. Agric. For. Meteorol. 2018, 260, 165–175. [Google Scholar] [CrossRef]
- Rogers, J.C.; Mosley-Thompson, E. Atlantic Arctic cyclones and the mild Siberian winters of the 1980s. Geophys. Res. Lett. 1995, 22, 799–802. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Wang, J.; Jin, L.; Zhang, Q.; Li, J.; Chen, J. Rapid warming in mid-latitude central Asia for the past 100 years. Front. Earth Sci. China 2009, 3, 42–50. [Google Scholar] [CrossRef]
- Tchebakova, N.M.; Rehfeldt, G.E.; Parfenova, E.I. From vegetation zones to climatypes: Effects of climate warming on Siberian ecosystems. In Permafrost Ecosystems; Osawa, A., Zyryanova, O., Matsuura, Y., Kajimoto, T., Wein, R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 427–446. [Google Scholar] [CrossRef]
- Asante, D.K.; Yakovlev, I.A.; Fossdal, C.G.; Holefors, A.; Opseth, L.; Olsen, J.E.; Junttila, O.; Johnsen, Ø. Gene expression changes during short day induced terminal bud formation in Norway spruce. Plant. Cell. Environ. 2011, 34, 332–346. [Google Scholar] [CrossRef]
- Jyske, T.; Mäkinen, H.; Kalliokoski, T.; Nöjd, P. Intra-annual tracheid production of Norway spruce and Scots pine across a latitudinal gradient in Finland. Agric. For. Meteorol. 2014, 194, 241–254. [Google Scholar] [CrossRef]
- Rathgeber, C.B.; Pérez-de-Lis, G.; Fernández-de-Uña, L.; Fonti, P.; Rossi, S.; Treydte, K.; Gessler, A.; Deslauriers, A.; Fonti, M.V.; Ponton, S. Anatomical, developmental and physiological bases of tree-ring formation in relation to environmental factors. In Stable Isotopes in Tree Rings; Siegwolf, R.T.W., Brooks, J.R., Roden, J., Saurer, M., Eds.; Springer: Cham, Switzerland, 2022; pp. 61–99. [Google Scholar] [CrossRef]
- Szeicz, J.M.; MacDonald, G.M. Age-dependent tree-ring growth responses of subarctic white spruce to climate. Can. J. For. Res. 1994, 24, 120–132. [Google Scholar] [CrossRef]
- Schuster, R.; Oberhuber, W. Age-dependent climate–growth relationships and regeneration of Picea abies in a drought-prone mixed-coniferous forest in the Alps. Can. J. For. Res. 2013, 43, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Primicia, I.; Camarero, J.J.; Janda, P.; Čada, V.; Morrissey, R.C.; Trotsiuk, V.; Bače, R.; Teodosiu, M.; Svoboda, M. Age, competition, disturbance and elevation effects on tree and stand growth response of primary Picea abies forest to climate. For. Ecol. Manag. 2015, 354, 77–86. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; McRae, D.J. Spatial and age-dependent tree-ring growth responses of Larix gmelinii to climate in northeastern China. Trees 2009, 23, 875–885. [Google Scholar] [CrossRef]
- Gurskaya, M.A.; Shiyatov, S.G. Distribution of frost injuries in the wood of conifers. Russ. J. Ecol. 2006, 37, 7–12. [Google Scholar] [CrossRef]
- Tuovinen, M.; Jalkanen, R.; MccArroll, D. The effect of severe ground frost on Scots pine (Pinus sylvestris) trees in northern Finland and implications for palaeoclimate reconstruction. Fennia 2005, 183, 109–117. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Site Code | °N | °E | Range of Elevation, m a.s.l. | Slope | Sample | Chronology | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Trees/Cores | Cover Period, Years | Length, Years | Mean TRW, mm | SD | r-bar | sens | ||||||
| Sayansky Pass 1 | SPass | 51.71 | 89.86 | 1970–2020 | SSW | 52/80 | 1524–2019 | 496 | 0.58 | 0.14 | 0.30 | 0.16 |
| SPassY | 19/38 | 1912–2019 | 108 | 1.08 | 0.12 | 0.35 | 0.15 | |||||
| SPassM | 17/21 | 1778–2019 | 242 | 0.62 | 0.17 | 0.35 | 0.18 | |||||
| SPassO | 16/21 | 1524–2019 | 496 | 0.50 | 0.15 | 0.41 | 0.16 | |||||
| Gladenkaya Mt. | GladSW | 52.91 | 91.36 | 1600–1640 | SW | 27/30 | 1633–2017 | 385 | 0.63 | 0.15 | 0.35 | 0.16 |
| GladE | 52.92 | 91.37 | 1530–1570 | E | 27/27 | 1786–2017 | 232 | 0.90 | 0.14 | 0.28 | 0.15 | |
| Ergaki Ridge, Lake Oyskoe | ErgO | 52.86 | 93.25 | 1520–1600 | S | 22/22 | 1688–2018 | 331 | 0.94 | 0.15 | 0.40 | 0.17 |
| Ergaki Ridge, Vidovka Mt. | ErgV | 52.80 | 93.43 | 1600–1680 | SSW | 22/23 | 1642–2020 | 379 | 0.85 | 0.16 | 0.51 | 0.18 |
| Ergaki Ridge, Lake Zolotarnoe | ErgZ | 52.82 | 93.44 | 1700–1780 | WSW | 16/16 | 1790–2020 | 231 | 0.56 | 0.21 | 0.37 | 0.22 |
| Chronologies per se | High-Frequency (below Diagonal) and Low-Frequency (above Diagonal) Components Separated with a 5-Year Smoothing Filter | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | SPass | GladSW | GladE | ErgO | ErgV | SPass | GladSW | GladE | ErgO | ErgV | ErgZ |
| SPass | 0.66 | 0.55 | 0.72 | 0.68 | 0.48 | ||||||
| GladSW | 0.70 | 0.72 | 0.73 † | 0.68 | 0.67 | 0.59 | |||||
| GladE | 0.66 | 0.84 † | 0.68 | 0.87 † | 0.69 | 0.45 | 0.41 | ||||
| ErgO | 0.67 | 0.75 | 0.73 | 0.69 | 0.78 | 0.74 | 0.66 † | 0.60 † | |||
| ErgV | 0.69 | 0.72 | 0.58 | 0.76 † | 0.71 | 0.70 | 0.56 | 0.80 † | 0.61 † | ||
| ErgZ | 0.70 | 0.65 | 0.57 | 0.66 † | 0.74 † | 0.74 | 0.67 | 0.58 | 0.73 † | 0.80 † | |
| Climatic Variable | Chronology | |||||
|---|---|---|---|---|---|---|
| SPass | GladSW | GladE | ErgO | ErgV | ErgZ | |
| Seasonality (beginning and ending dates) | ||||||
| Temperature | ||||||
| prev | 15 Aug. *–23 Sep. * | 25 Jul. *–11 Sep. * | 18 Jul. *–23 Sep. * | 15 Aug. *–22 Sep. * | 25 Jul. *–20 Sep. * | 14 Aug. *–4 Oct. * |
| cold | 10 Nov. *–26 Mar. | 12 Dec. *–26 Mar. | 24 Nov. *–26 Mar. | 10 Dec. *–26 Mar. | 19 Nov. *–27 Mar. | 16 Nov. *–18 Mar. |
| prev + cold | 15 Aug. *–26 Mar. | 25 Jul. *–26 Mar. | 18 Jul. *–26 Mar. | 15 Aug. *–26 Mar. | 25 Jul. *–27 Mar. | 22 Jul. *–18 Mar. |
| spring | 3 Apr.–10 May | 5 Apr.–3 May | 3 Apr.–3 May | 5 Apr.–3 May | 8 Apr.–11 May | 28 Mar.–11 May |
| curr | 12 May–25 Aug. | 18 May–31 Aug. | 13 May–1 Sep. | 13 May–3 Sep. | 23 May–2 Sep. | 24 May–13 Aug. |
| Precipitation | ||||||
| prev | 8 Sep. *–25 Dec. * | 13 Jul. *–1 Dec. * | 14 Jul. *–25 Dec. * | 29 Aug. *–30 Nov. * | 24 Jul. *–1 Dec. * | 5 Sep. *–25 Dec. * |
| cold | 26 Dec. *–30 Mar. | 2 Dec. *–4 Apr. | 26 Dec. *–4 Apr. | 1 Dec. *–20 Apr. | 2 Dec. *–20 Apr. | 26 Dec. *–4 Apr. |
| curr | 5 Apr.–8 Aug. | 12 Apr.–10 Aug. | 5 Apr.–7 Aug. | 28 Apr.–21 Jul. | 21 Apr.–10 Aug. | 20 Apr.–6 Aug. |
| Correlations between time series per se | ||||||
| Temperature | ||||||
| prev | 0.26 | 0.29 | 0.22 | 0.26 | 0.20 | 0.11 |
| cold | 0.30 | 0.35 | 0.33 | 0.44 | 0.38 | 0.25 |
| prev + cold | 0.31 | 0.33 | 0.35 | 0.40 | 0.38 | 0.27 |
| spring | −0.17 | −0.21 | −0.19 | −0.22 | −0.19 | −0.25 |
| curr | 0.37 | 0.20 | 0.20 | 0.31 | 0.38 | 0.22 |
| Precipitation | ||||||
| prev | −0.37 | −0.37 | −0.30 | −0.29 | −0.24 | −0.36 |
| cold | 0.13 | 0.07 | 0.04 | 0.25 | 0.25 | 0.25 |
| curr | −0.28 | −0.32 | −0.27 | −0.21 | −0.25 | −0.19 |
| Correlations between time series separated into high-frequency/low-frequency domains with 5-year smoothing filter | ||||||
| Temperature | ||||||
| prev | 0.22/0.50 | 0.23/0.36 | 0.20/0.31 | 0.07/0.64 | −0.02/0.59 | 0.12/0.40 |
| cold | 0.33/0.42 | 0.37/0.28 | 0.32/0.45 | 0.42/0.51 | 0.43/0.33 | 0.35/0.29 |
| prev + cold | 0.35/0.48 | 0.39/0.26 | 0.38/0.42 | 0.34/0.58 | 0.39/0.44 | 0.38/0.43 |
| spring | −0.21/0.18 | −0.27/−0.18 | −0.24/−0.07 | −0.30/−0.21 | −0.29/0.04 | −0.17/0.04 |
| curr | 0.44/0.36 | 0.35/0.07 | 0.27/0.20 | 0.36/0.35 | 0.45/0.46 | 0.39/0.47 |
| Precipitation | ||||||
| prev | −0.49/−0.22 | −0.45/−0.31 | −0.39/−0.01 | −0.29/−0.11 | −0.28/−0.16 | −0.43/0.21 |
| cold | 0.02/0.13 | −0.04/0.25 | 0.00/−0.18 | 0.13/0.36 | 0.13/0.42 | 0.12/0.35 |
| curr | −0.47/0.00 | −0.41/−0.27 | −0.37/−0.24 | −0.31/0.02 | −0.37/−0.20 | −0.32/−0.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhirnova, D.F.; Belokopytova, L.V.; Krutovsky, K.V.; Kholdaenko, Y.A.; Babushkina, E.A.; Vaganov, E.A. Spatial-Coherent Dynamics and Climatic Signals in the Radial Growth of Siberian Stone Pine (Pinus sibirica Du Tour) in Subalpine Stands along the Western Sayan Mountains. Forests 2022, 13, 1994. https://doi.org/10.3390/f13121994
Zhirnova DF, Belokopytova LV, Krutovsky KV, Kholdaenko YA, Babushkina EA, Vaganov EA. Spatial-Coherent Dynamics and Climatic Signals in the Radial Growth of Siberian Stone Pine (Pinus sibirica Du Tour) in Subalpine Stands along the Western Sayan Mountains. Forests. 2022; 13(12):1994. https://doi.org/10.3390/f13121994
Chicago/Turabian StyleZhirnova, Dina F., Liliana V. Belokopytova, Konstantin V. Krutovsky, Yulia A. Kholdaenko, Elena A. Babushkina, and Eugene A. Vaganov. 2022. "Spatial-Coherent Dynamics and Climatic Signals in the Radial Growth of Siberian Stone Pine (Pinus sibirica Du Tour) in Subalpine Stands along the Western Sayan Mountains" Forests 13, no. 12: 1994. https://doi.org/10.3390/f13121994