
Type 2 Nondeep Physiological Dormancy in Seeds of Fraxinus chinensis subsp. rhynchophylla (Hance) A.E.Murray
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
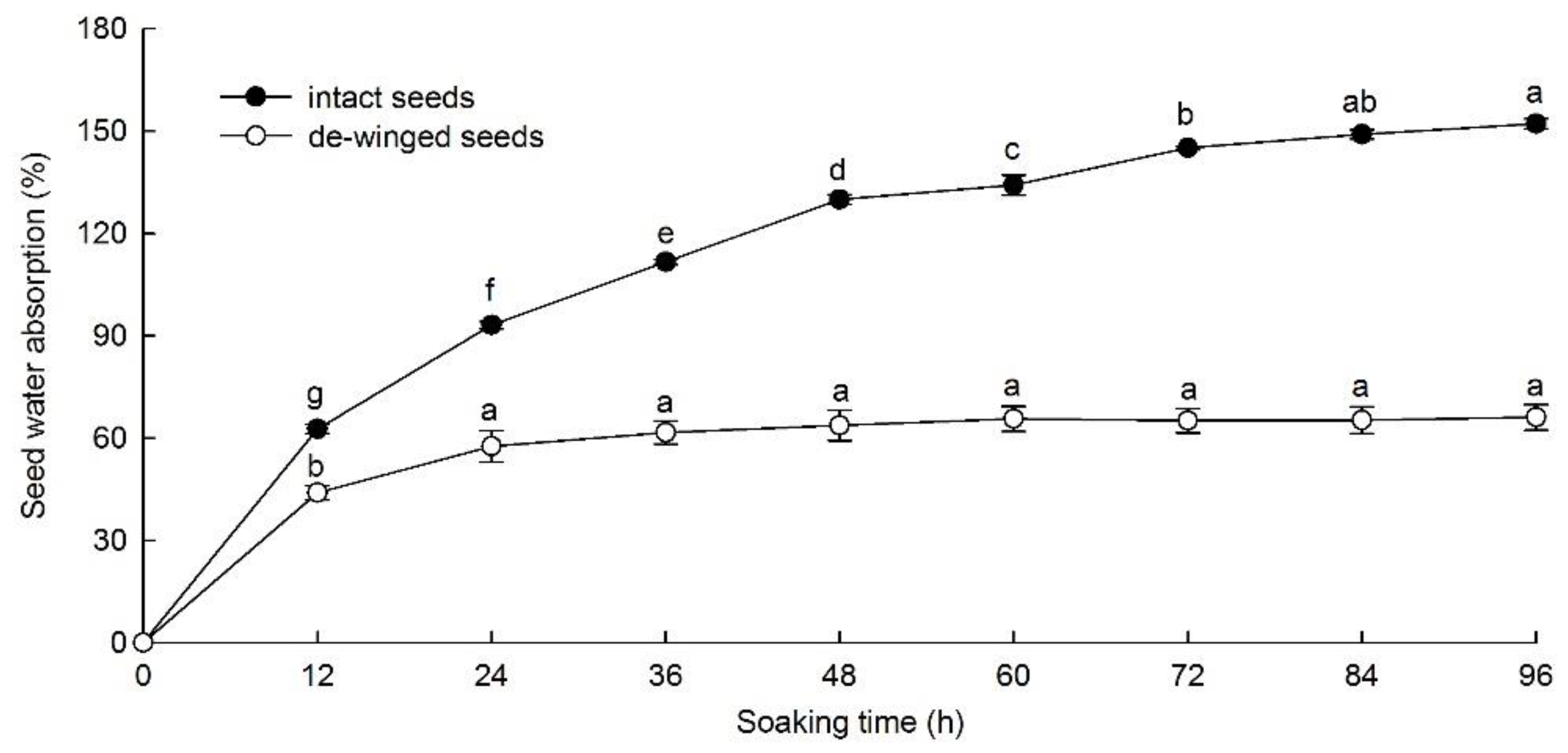
2.2. Water Uptake Test
2.3. Cold Stratification
2.4. Developmental Status of Seed Embryo
2.5. Germination Test
2.6. Statistical Analyses
3. Results
3.1. Seeds Water Permeability
3.2. Embryo Developmental Status
3.3. Germination of Cold Stratified Seeds at Different Temperatures
3.3.1. Germination of Seeds after Different Cold Stratification Time
3.3.2. Germination of Non-Stratified Seeds at Different Temperatures
3.3.3. Germination after 4 Weeks Cold Stratification at Different Temperatures
3.3.4. Germination after 8 Weeks Cold Stratification at Different Temperatures
3.3.5. Germination of Seeds at Constant Temperatures after Cold Stratification
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baskin, J.M.; Baskin, C.C. The great diversity in kinds of seed dormancy: A revision of the Nikolaeva–Baskin classification system for primary seed dormancy. Seed Sci. Res. 2021, 31, 249–277. [Google Scholar] [CrossRef]
- Finchsavage, W.; Leubnermetzger, G. Tansley review: Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Nikolaeva, M.G. Physiology of deep dormancy in seeds. In Seeds; NSF: Washington, DC, USA, 1969. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: London, UK, 2014. [Google Scholar]
- Kortessis, N.; Chesson, P. Germination variation facilitates the evolution of seed dormancy when coupled with seedling competition. Theor. Popul. Biol. 2019, 130, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Tonguc, T. Dormancy level and dormancy-breaking pretreatments in seeds of Fraxinus ornus subsp. cilicica. Propag. Ornam. Plants 2013, 13, 40–45. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Chapter 3 Types of Seeds and Kinds of Seed Dormancy. In Seeds, 2nd ed.; Academic Press: New York, NY, USA, 2014; pp. 37–77. [Google Scholar]
- Finch-Savage, W.E.; Footitt, S. Seed dormancy cycling and the regulation of dormancy mechanisms to time germination in variable field environments. J. Exp. Bot. 2017, 68, 843–856. [Google Scholar] [CrossRef] [Green Version]
- Baskin, C.C.; Baskin, J.M. Chapter 4 Germination Ecology of Seeds with Nondeep Physiological Dormancy. In Seeds, 2nd ed.; Academic Press: New York, NY, USA, 2014; pp. 79–117. [Google Scholar]
- Hema, S.N.D.; Steven, J.S. Variation of Seed Dormancy and Germination Ecology of Cowcockle (Vaccaria hispanica). Weed Sci. 2014, 62, 483–492. [Google Scholar]
- Vegis, A. Chapter 15 Climatic Control of Germination, Bud Break, and Dormancy. In Environmental Control of Plant Growth; Academic Press: New York, NY, USA, 1963; pp. 265–287. [Google Scholar]
- Cheng, P.; Wang, P.; Sun, J.K.; Fei, M.L.; Yang, H. Research progresses on regulation mechanisms of plant seeds dormancy and germination. J. Cent. South Univ. 2013, 33, 52–58. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Chapter 6 Germination Ecology of Seeds with Physical Dormancy. In Seeds, 2nd ed.; Academic Press: New York, NY, USA, 2014; pp. 145–185. [Google Scholar]
- Zhang, P. Seed characteristic variation and seedling growth variation among progenies from plus trees of Fraxinus rhynohophylla. China For. Sci. Technol. 2013, 2, 28–31. [Google Scholar]
- Zhang, F.; Hu, W.L.; Kong, W.X. Comparison of five broad-leaved seedlings’ physiological characteristics in natural secondary forest of eastern Liaoning. Chin. J. Ecol. 2004, 23, 106–110. [Google Scholar]
- Xiang, F.W.; Huang, Y.F.; Wang, Z.L. On the Provenance Test during Seedling Stage of Fraxinus chinensis Hance in the Southeast Mountain Area of Jilin Province. J. Beihua Univ. (Nat. Sci.) 2013, 3, 255–258. [Google Scholar]
- Sun, J.J.; Yao, C.J.; Liu, Y.A. Investigation on the Suitable Locality of Fraxinus chinensis. J. Jilin For. Univ. 1999, 15, 239–241. [Google Scholar]
- Sun, J.J.; Yao, C.J.; Jia, H.Y. The Speed-up and High Yield Breeding Technology for Fraxinus chinensis. J. Jilin For. Univ. 1999, 15, 242–244. [Google Scholar]
- Xiang, F.W.; Yao, C.J.; Liu, Y.H. On the Distribution Growth and Natural Regeneration of Fraxinus chinensis. J. Jilin For. Univ. 1997, 13, 212–215. [Google Scholar]
- Wang, X.X. Seeding and seedling raising techniques of economic tree Fraxinus rhynchophylla. For. By-Prod. Spec. China 2020, 167, 66–67. [Google Scholar]
- Ashley, J.A.; Preece, J.E. Seed cutting treatments stimulate germination and elucidate a dormancy gradient in dormant Fraxinus Americana L. and Fraxinus Pennsylvanica Marsh. Propag. Ornam. Plants 2009, 9, 121–128. [Google Scholar]
- Piotto, B. Storage of non-dormant seeds of Fraxinus angustifolia Vahl. New For. 1997, 14, 157–166. [Google Scholar] [CrossRef]
- Emery, D.E. Seed propagation of native California plants. ICSID Rev. 1988, 29, 451–473. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Chapter 9 A Geographical Perspective on Germination Ecology. In Seeds, 2nd ed.; Academic Press: New York, NY, USA, 2014; pp. 689–807. [Google Scholar]
- Ye, Y.; Wang, C.; Shi, Y. Preliminary study on cause of seed dormancy of Fraxinus hupehensis. Hubei Agric. Sci. 1999, 20, 45–47. [Google Scholar]
- Cram, W.H.; Lindquist, C.H. Germination of green ash is related to seed moisture content at harvest (Fraxinus pennsylvanica). For. Sci. 1982, 28, 809–812. [Google Scholar]
- Ye, J.F. Study on dormancy and seedling of Fraxinus americana. Heilongjiang Agric. Sci. 2015, 03, 61–63. [Google Scholar]
- Liu, Y.; Zhang, J.; Li, G.; Wang, Q.; Xu, S. The research for the germination characteristics of Fraxinus hupehensis Seeds. Seed. 2016, 35, 27–31. [Google Scholar]
- Tinus, R.W. Effects of de-winging, soaking, stratification, and growth regulators on germination of green ash seed. Can. J. For. Res. 1982, 12, 931–935. [Google Scholar] [CrossRef]
- Asakawa, S. Some observations on Fraxinus seeds. J. Jpn. For. Soc. 2008, 37, 1–5. [Google Scholar]
- Ferenczy, L. The dormancy and germination of seeds of the Fraxinus excelsior L. Acta Biol. 1955, 12, 17–24. [Google Scholar]
- Wagner, J. Changes in dormancy levels of Fraxinus excelsior L. embryos at different stages of morphological and physiological maturity. Trees 1996, 10, 177–182. [Google Scholar]
- Zhang, P.; Shen, H. Morphological and Physiological Changes of Manchurian Ash (Fraxinus mandshurica Rupr.) Seeds Collected at Different Developmental Periods and Its Germination Responses after Stratification. Plant Physiol. 2010, 46, 125–130. [Google Scholar]
- Ji, Y. Effects of dry storage at different stage of seed stratification on seed germination of Fraxinus mandshurica. Terr. Nat. Resour. Stu. 2019, 06, 75–77. [Google Scholar]
- Asakawa, S. Studies on the delayed germination of Fraxinus mandshurica var. japonica seeds. Bull. Gov. For. Exp. Stn. Tokyo 1956, 83, 19–28. [Google Scholar]
- Hinsinger, D.D.; Gaudeul, M.; Couloux, A.; Bousquet, J.; Frascaria-Lacoste, N. The phylogeography of Eurasian Fraxinus species reveals ancient transcontinental reticulation. Mol. Phylogenet. Evol. 2014, 77, 223–237. [Google Scholar] [CrossRef]
- Steinbauer, G.P. Dormancy and germination of Fraxinus seeds. Plant Physiol. 1935, 12, 813–824. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Chapter 5 Germination Ecology of Seeds with Morphophysiological Dormancy. In Seeds, 2nd ed.; Academic Press: New York, NY, USA, 2014; pp. 119–143. [Google Scholar]
- Dou, Q.L.; Zhang, R.B. Seed germination characteristics of Celastrus rosthornianus loes. J. Plant Physiol. 2013, 49, 75–80. [Google Scholar]
- Pan, R.; Sun, W.B. Seed Dormancy and Germination of the Critically Endangered Manglietia grandis Hu et Cheng. Plant Physiol. Commun. 2009, 45, 1089–1092. [Google Scholar]
- Zhang, L.W.; Liu, H.L.; Zhang, D.Y.; Bian, W.G. Seed dormancy release and germination characteristics of Corispermum lehmannianum Bunge, an endemic species in the Gurbantunggut desert of China. Phyton 2015, 84, 58–63. [Google Scholar]
- Sui, X.L.; Li, A.R.; Guan, K.Y. Impacts of climatic changes as well as seed germination characteristics on the population expansion of Pedicularis verticillata. Ecol. Environ. Sci. 2013, 22, 1099–1104. [Google Scholar]
- Chien, C.T.; Yang, J.C.; Lin, T.P. Seed storage behavior of Lindera communis, Lindera megaphylla, Phoebe formosana, Helicia cochinchinensis, and Helicia formosana in Taiwan. Taiwan J. For. Sci. 2004, 19, 119–131. [Google Scholar]
- Zhang, H.B.; Wang, Z.; Yang, X.; Liu, T.Y.; Zhang, P. Effect of Seed Soaking with Plant Growth Regulators and Stratification on Seed Germination of Fraxinus rhynchophylla. Heilongjiang Sci. 2022, 9, 1248–1261. [Google Scholar]
- Soltani1, E.; Baskin, C.C.; Baskin, J.M. A graphical method for identifying the six types of nondeep physiological dormancy in seeds. Plant Biol. 2017, 19, 673–682. [Google Scholar] [CrossRef]


{kind=link}
| Cold Stratification (Week) | E (1) (mm) | E:S (2) Ratio (%) | Embryo:Seed Dry Weight Ratio (%) |
|---|---|---|---|
| 0 | 13.9 ± 0.2 a | 93.4 ± 0.7 a | 33.2 ± 0.5 a |
| 2 | 13.5 ± 0.3 a | 94.5 ± 0.5 a | 33.2 ± 0.5 a |
| 4 | 13.2 ± 0.3 a | 93.4 ± 0.4 a | 33.0 ± 0.6 a |
| 6 | 13.8 ± 0.3 a | 93.9 ± 0.6 a | 32.0 ± 0.3 a |
| 8 | 13.6 ± 0.3 a | 94.8 ± 0.6 a | 33.6 ± 0.9 a |
| Cold Stratification Time | 4 Weeks/8 Weeks | 8 Weeks/12 Weeks | ||||
|---|---|---|---|---|---|---|
| Source of Variation | p Value | |||||
| GP (1) | GI (2) | MGT (3) | GP | GI | MGT | |
| Cold stratification time | 0.007 | <0.001 | 0.128 | 0.251 | 0.363 | 0.017 |
| Temperature | <0.001 | <0.001 | 0.006 | <0.001 | <0.001 | <0.001 |
| Cold stratification time × temperature | 0.012 | 0.195 | 0.031 | <0.001 | 0.189 | 0.149 |
| Temperature (°C) | GP (1) | GI (2) | MGT (3) (d) |
|---|---|---|---|
| 35 | 26.0 ± 4.2 f | 0.4 ± 0.1 ghi | 17.5 ± 0.6 abc |
| 30 | 52.0 ± 2.8 abcd | 1.0 ± 0.1 cdefg | 15.0 ± 1.3 bcde |
| 25 | 63.0 ± 6.0 ab | 1.1± 0.1 bcde | 15.3 ± 0.4 bcde |
| 20 | 28.0 ± 6.3 ef | 0.5 ± 0.1 fghi | 15.9 ± 1.3 bcde |
| 15 | 6.0 ± 1.2 g | 0.1 ± 0.0 i | 17.6 ± 3.0 ab |
| 10 | 5.0 ± 2.5 g | 0.1 ± 0.1 g | - |
| 5 | 1.0 ± 1.0 g | 0.0 ± 0.0 g | - |
| 35/5 | 48.0 ± 5.4 abcde | 1.0 ± 0.2 cdefg | 15.5 ± 1.4 bcde |
| 35/10 | 35.0 ± 10.4 abcd | 1.3 ± 0.2 abcde | 13.4 ± 2.0 cde |
| 35/15 | 56.0± 4.3 abc | 1.7 ± 0.4 ab | 13.1 ± 1.9 de |
| 35/20 | 46.0 ± 6.2 abcdef | 1.4 ± 0.3 abcde | 12.1 ± 2.0 e |
| 35/25 | 66.0 ± 4.2 ab | 1.8 ± 0.3 a | 12.8 ± 1.1 de |
| 35/30 | 39.0 ± 4.1 cdef | 0.8 ± 0.1 efgh | 16.7 ± 1.1 abcd |
| 30/5 | 56.0 ± 7.1 abc | 1.1± 0.2 bcdef | 15.4 ± 0.5 bcde |
| 30/10 | 44.0 ± 6.3 bcdef | 0.8 ± 0.1 defgh | 16.1 ± 1.3 bcde |
| 30/15 | 67.0 ± 9.7 a | 1.8 ± 0.4 a | 13.1 ± 1.7 de |
| 30/20 | 52.0 ± 4.3 abcd | 1.1 ± 0.1 bcdef | 13.3 ± 0.5 cde |
| 30/25 | 52.0 ± 4.9 abcd | 1.3 ± 0.1 abcde | 13.0 ± 0.7 de |
| 25/5 | 48.0 ± 5.9 abcde | 1.0 ± 0.2 cdefgh | 15.1 ± 1.3 bcde |
| 25/10 | 65.0 ± 8.1 ab | 1.3 ± 0.2 abcde | 14.9 ± 0.2 bcde |
| 25/15 | 61.0 ± 8.9 abc | 1.5 ± 0.3 abcd | 12.2 ± 0.6 e |
| 25/20 | 63.0 ± 1.9 ab | 1.5 ± 0.1 abc | 12.8 ± 1.0 de |
| 20/5 | 53.0 ± 9.2 abcd | 0.9 ± 0.1 cdefgh | 16.7 ± 0.3 abcd |
| 20/10 | 66.0 ± 7.4 ab | 1.3 ± 0.2 abcde | 14.6 ± 0.3 bcde |
| 20/15 | 49.0 ± 13.4 abcd | 0.9 ± 0.3 cdefgh | 16.0 ± 0.5 bcde |
| 15/5 | 33.0 ± 3.8 def | 0.5 ± 0.1 ghi | 18.6 ± 0.4 ab |
| 15/10 | 27.0 ± 5.3 f | 0.4 ± 0.1 hi | 20.4 ± 0.7 a |
| 10/5 | 8.0 ± 2.8 g | 0.1 ± 0.0 g | - |
| Temperature (°C) | GP (1) | GI (2) | MGT (3) (d) |
|---|---|---|---|
| 35 | 49.0 ± 6.8 fg | 0.8 ± 0.1 k | 18.2 ± 0.8 a |
| 30 | 59.0 ± 3.0 defg | 1.2 ± 0.1 hijk | 13.2 ± 0.2 bcdef |
| 25 | 56.0 ± 4.3 defg | 1.1 ± 0.1 ijk | 13.9 ± 0.5 bc |
| 20 | 52.0 ± 3.7 efg | 1.1 ± 0.1 ijk | 13.2 ± 0.6 bcdef |
| 15 | 47.0 ± 3.4 g | 0.9 ± 0.1 jk | 13.7 ± 0.4 bc |
| 10 | 57.0 ± 3.8 defg | 0.8 ± 0.1 k | 18.9 ± 1.4 a |
| 5 | 3.0 ± 1.0 i | 0.1 ± 0.0 l | - |
| 35/5 | 52.0 ± 8.2 efg | 1.1 ± 0.2 ijk | 13.2 ± 0.7 bcdef |
| 35/10 | 48.0 ± 3.3 g | 1.1 ± 0.1 ijk | 12.6 ± 0.8 bcdefgh |
| 35/15 | 70.0 ± 4.2 abcde | 2.0 ± 0.2 bcd | 10.8 ± 0.6 ghij |
| 35/20 | 67.0 ± 5.5 abcdef | 1.9 ± 0.2 bcdef | 11.3 ± 0.6 efghij |
| 35/25 | 58.0 ± 5.3 defg | 1.6 ± 0.2 cdefghi | 11.9 ± 0.9 cdefghi |
| 35/30 | 59.0 ± 1.0 defg | 1.4 ± 0.0 defghijk | 12.8 ± 0.7 bcdefg |
| 30/5 | 61.0 ± 8.9 cdefg | 1.3 ± 0.2 fghijk | 13.4 ± 0.3 bcde |
| 30/10 | 65.0 ± 1.9 bcdefg | 1.8 ± 0.1 bcdefgh | 10.0 ± 0.9 ij |
| 30/15 | 77.0 ± 6.0 abc | 2.3 ± 0.3 ab | 11.0 ± 0.9 ghij |
| 30/20 | 77.0 ± 7.7 abc | 2.6 ± 0.3 a | 9.5 ± 0.2 j |
| 30/25 | 63.0 ± 7.0 bcdefg | 1.9 ± 0.5 bcdef | 10.7 ± 0.9 ghij |
| 25/5 | 72.0 ± 5.9 abcd | 1.5 ± 0.2 cdefghij | 13.5 ± 0.7 bcd |
| 25/10 | 67.0 ± 5.7 abcdef | 1.7 ± 0.2 bcdefghi | 11.2 ± 0.5 efghij |
| 25/15 | 79.0 ± 5.5 ab | 2.0 ± 0.1 bc | 11.4 ± 0.8 defghij |
| 25/20 | 68.0 ± 5.9 abcde | 1.8 ± 0.2 bcdefg | 10.4 ± 0.4 hij |
| 20/5 | 82.0 ± 3.5 a | 1.7 ± 0.1 bcdefghi | 12.8 ± 0.1 bcdefg |
| 20/10 | 77.0 ± 5.0 abc | 2.0 ± 0.2 bcde | 10.9 ± 0.5 ghij |
| 20/15 | 64.0 ± 2.8 bcdefg | 1.6 ± 0.1 bcdefghi | 11.1 ± 0.2 fghij |
| 15/5 | 69.0 ± 1.9 abcde | 1.2 ± 0.0 ghijk | 14.8 ± 0.1 b |
| 15/10 | 73.0 ± 6.2 abcd | 1.4 ± 0.1 efghijk | 14.7 ± 0.4 b |
| 10/5 | 37.0 ± 2.5 h | 0.5 ± 0.1 kl | - |
| Temperature (°C) | GP (1) | GI (2) | MGT (3) (d) |
|---|---|---|---|
| 35 | 40.0 ± 4.3 e | 1.5 ± 0.2 defg | 14.2 ± 0.6 bc |
| 30 | 62.0 ± 6.0 abcd | 2.5 ± 0.5 abcde | 10.5 ± 0.7 efghi |
| 25 | 68.0 ± 4.0 abcd | 2.4 ± 0.4 abcdef | 12.6 ± 1.0 cde |
| 20 | 51.0 ± 3.4 de | 2.4 ± 0.3 abcde | 11.6 ± 0.7 defg |
| 15 | 57.0 ± 3.4 abcde | 2.1 ± 0.2 bcdefg | 11.5 ± 0.6 defg |
| 10 | 62.0 ± 6.8 abcd | 1.6 ± 0.4 cdefg | 13.6 ± 0.9 bcd |
| 5 | 17.0 ± 1.0 f | 0.3 ± 0.1 h | 21.3 ± 1.0 a |
| 35/5 | 53.0 ± 10.0 cde | 1.4 ± 0.4 fg | 12.3 ± 0.8 cdef |
| 35/10 | 53.0 ± 5.7 cde | 2.3 ± 0.4 abcdefg | 8.6 ± 1.4 hij |
| 35/15 | 65.0 ± 4.1 abcd | 2.6 ± 0.2 abcd | 10.7 ± 0.3 efghi |
| 35/20 | 64.0 ± 4.3 abcd | 3.3 ± 0.3 a | 7.5 ± 0.6 j |
| 35/25 | 70.0 ± 6.8 abc | 2.9 ± 0.4 ab | 10.4 ± 0.3 efghi |
| 35/30 | 57.0 ± 5.7 abcde | 1.6 ± 0.3 cdefg | 11.6 ± 1.0 defg |
| 30/5 | 55.0 ± 8.5 bcde | 1.5 ± 0.2 efg | 11.8 ± 0.8 cdefg |
| 30/10 | 67.0 ± 5.5 abcd | 2.3 ± 0.2 abcdefg | 9.7 ± 0.8 fghij |
| 30/15 | 69.0 ± 4.1 abcd | 2.5 ± 0.2 abcde | 9.4 ± 0.5 ghij |
| 30/20 | 74.0 ± 3.8 a | 2.6 ± 0.2 abc | 9.3 ± 0.5 ghij |
| 30/25 | 73.0 ± 2.5 ab | 2.3 ± 0.3 abcdefg | 10.7 ± 1.0 efghi |
| 25/5 | 62.0 ± 5.3 abcd | 2.2 ± 0.3 abcdefg | 10.8 ± 0.6 efghi |
| 25/10 | 74.0 ± 3.5 a | 2.7 ± 0.1 abc | 8.2 ± 0.5 ij |
| 25/15 | 74.0 ± 6.0 a | 2.6 ± 0.5 abc | 9.3 ± 0.9 ghij |
| 25/20 | 73.0 ± 1.9 abcd | 2.3 ± 0.2 abcdefg | 9.9 ± 0.6 fghij |
| 20/5 | 61.0 ± 4.1 abcd | 1.8 ± 0.4 bcdefg | 11.4 ± 0.8 defg |
| 20/10 | 70.0 ± 4.8 abc | 2.0 ± 0.2 bcdefg | 11.1 ± 0.7 defgh |
| 20/15 | 67.0 ± 5.5 abcd | 2.9 ± 0.5 ab | 8.7 ± 0.7 hij |
| 15/5 | 69.0 ± 3.4 abcd | 1.7 ± 0.2 cdefg | 11.6 ± 0.6 defg |
| 15/10 | 68.0 ± 4.0 abcd | 2.4 ± 0.5 abcdefg | 10.4 ± 0.9 efghi |
| 10/5 | 60.0 ± 5.9 abcd | 1.3 ± 0.2 g | 15.5 ± 0.5 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, K.; Yang, M.; Baskin, C.C.; Li, M.; Zhu, M.; Jiao, C.; Wu, H.; Zhang, P. Type 2 Nondeep Physiological Dormancy in Seeds of Fraxinus chinensis subsp. rhynchophylla (Hance) A.E.Murray. Forests 2022, 13, 1951. https://doi.org/10.3390/f13111951
An K, Yang M, Baskin CC, Li M, Zhu M, Jiao C, Wu H, Zhang P. Type 2 Nondeep Physiological Dormancy in Seeds of Fraxinus chinensis subsp. rhynchophylla (Hance) A.E.Murray. Forests. 2022; 13(11):1951. https://doi.org/10.3390/f13111951
Chicago/Turabian StyleAn, Kangming, Minghan Yang, Carol Caudle Baskin, Mingyue Li, Meiru Zhu, Chunjing Jiao, Haibo Wu, and Peng Zhang. 2022. "Type 2 Nondeep Physiological Dormancy in Seeds of Fraxinus chinensis subsp. rhynchophylla (Hance) A.E.Murray" Forests 13, no. 11: 1951. https://doi.org/10.3390/f13111951