
Characterization of the Chloroplast Genome Structure of Gueldenstaedtia verna (Papilionoideae) and Comparative Analyses among IRLC Species
Abstract
:1. Introduction
2. Results
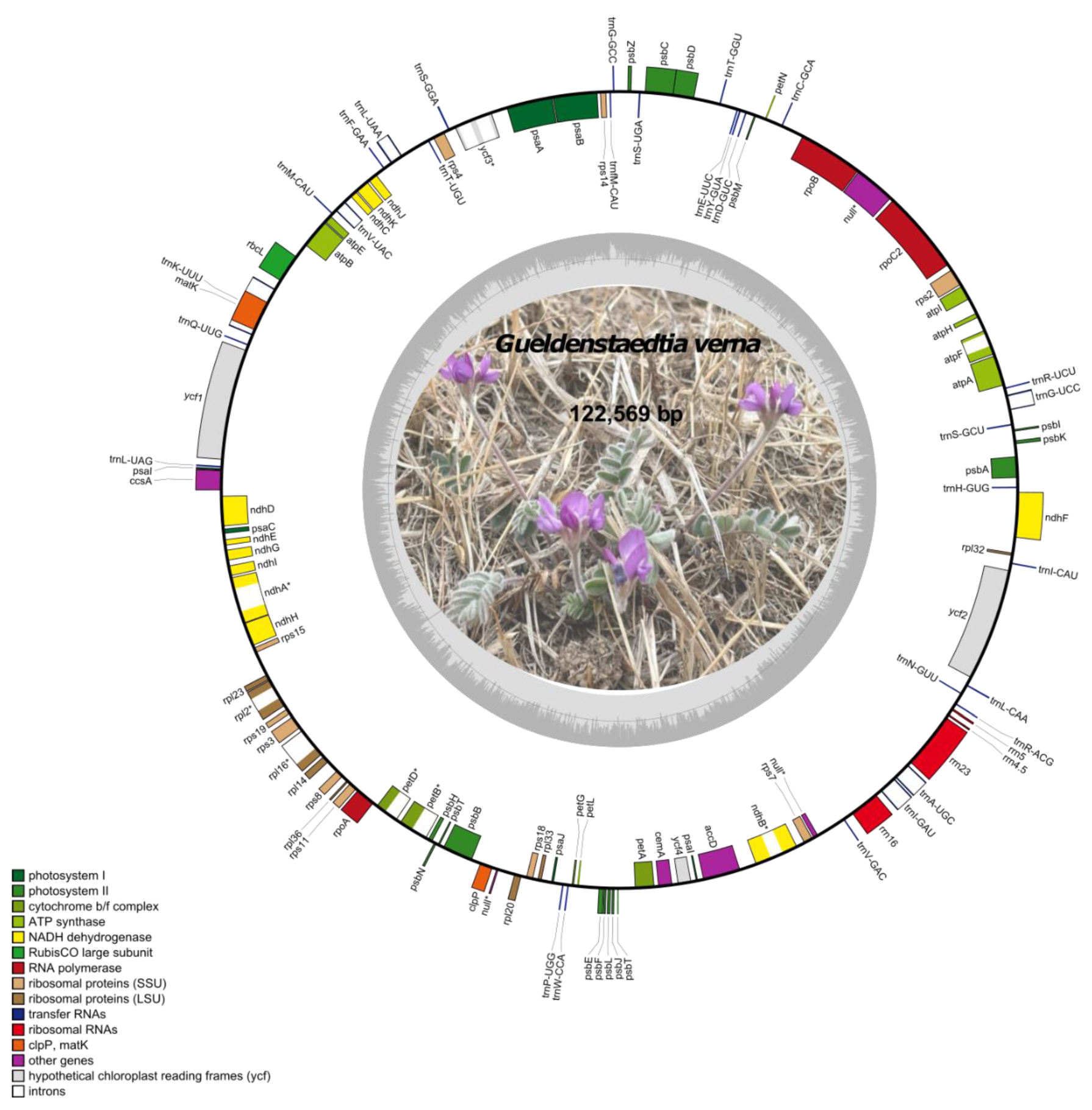
2.1. Characterization of the Chloroplast Genome of Gieldenstaedtia verna
2.2. Comparison of cp Genomes within Fabaceae
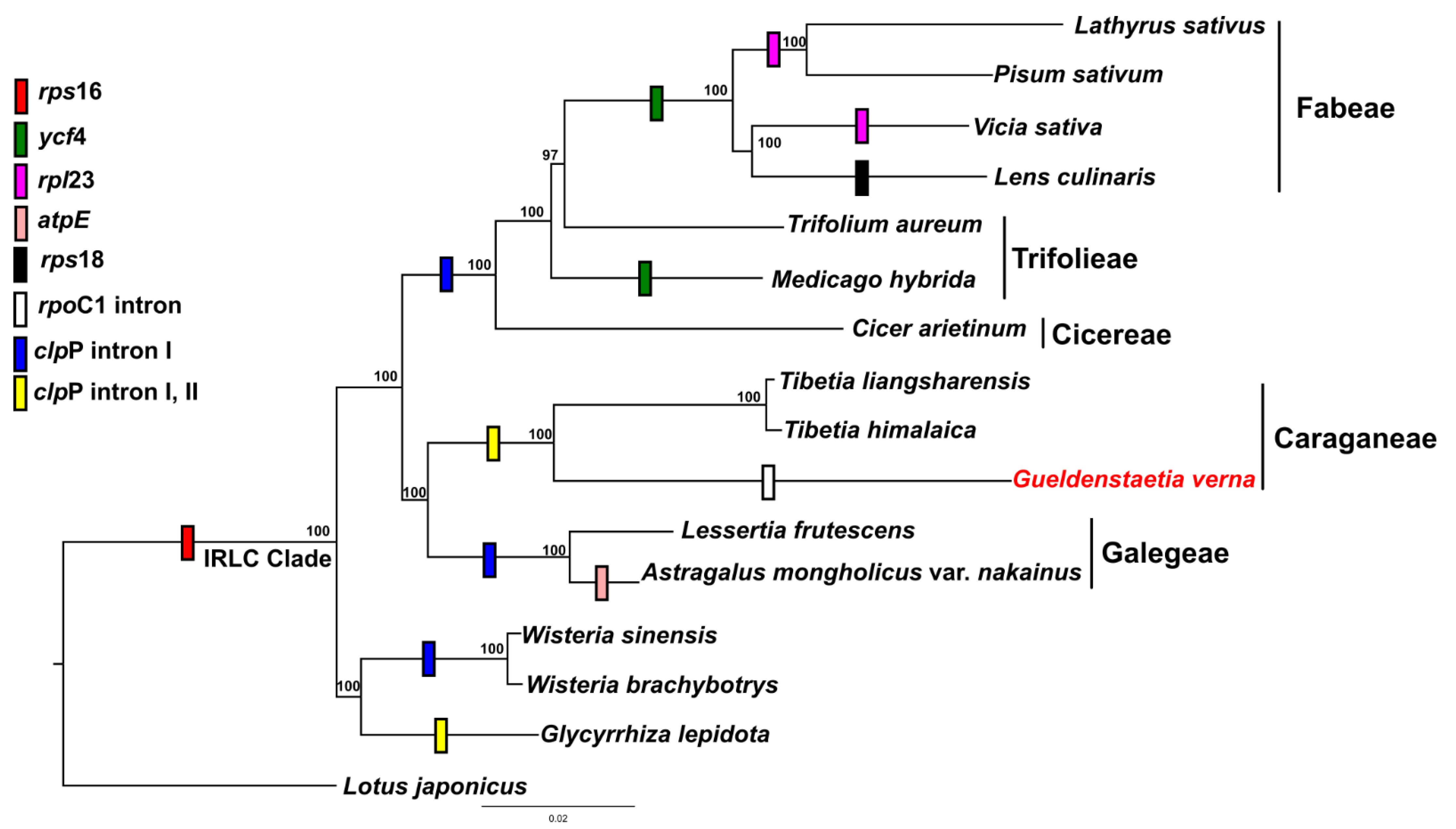
2.3. Phylogenetic Analysis
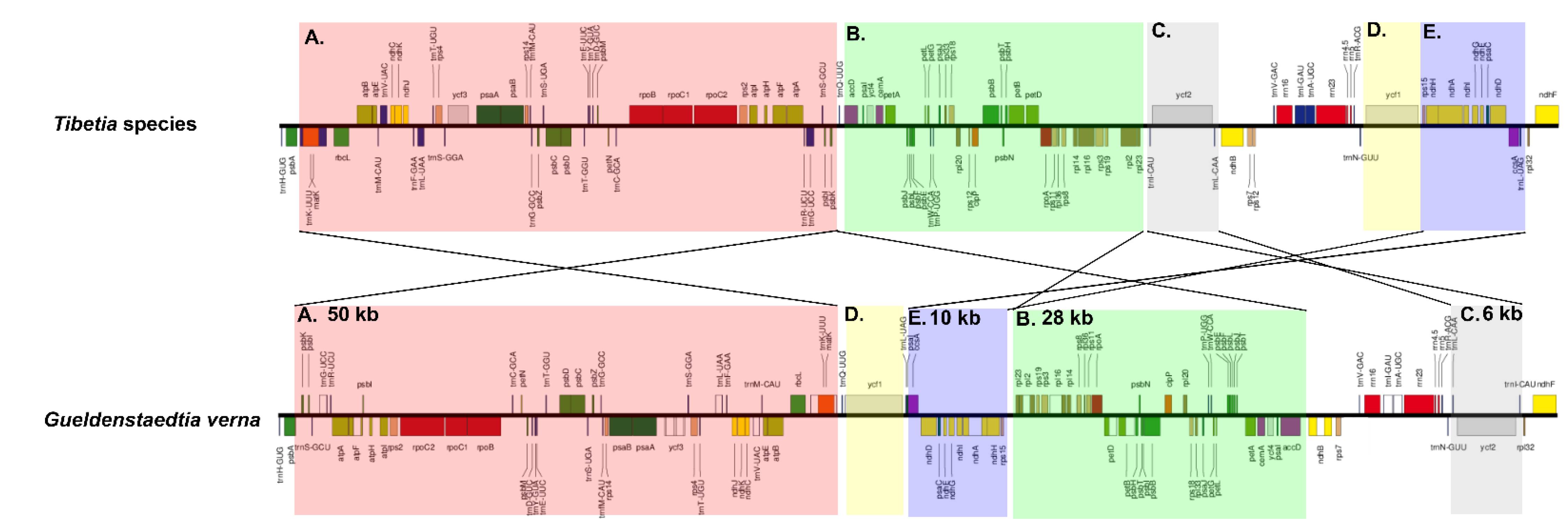
2.4. Inversion in cp of G. verna
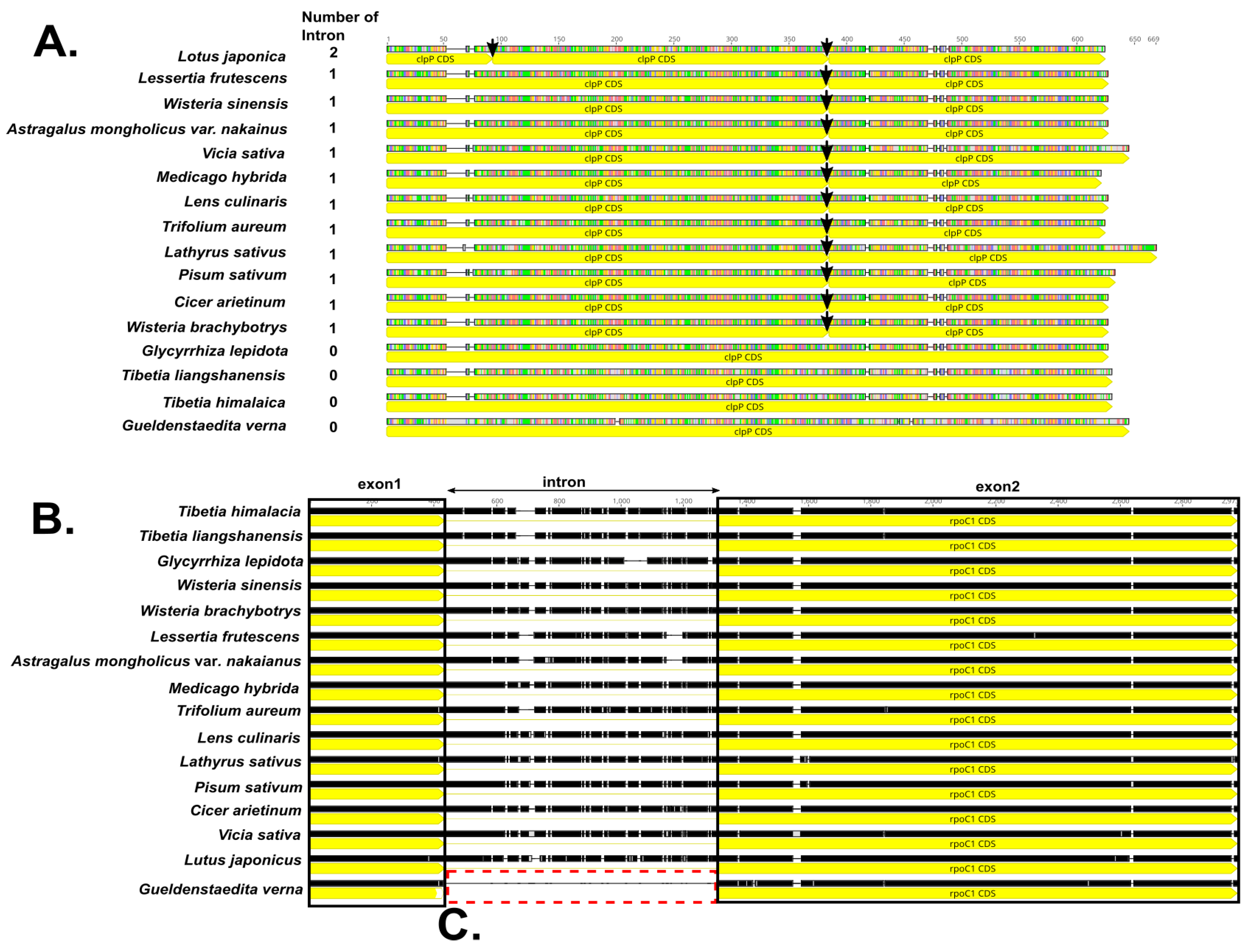
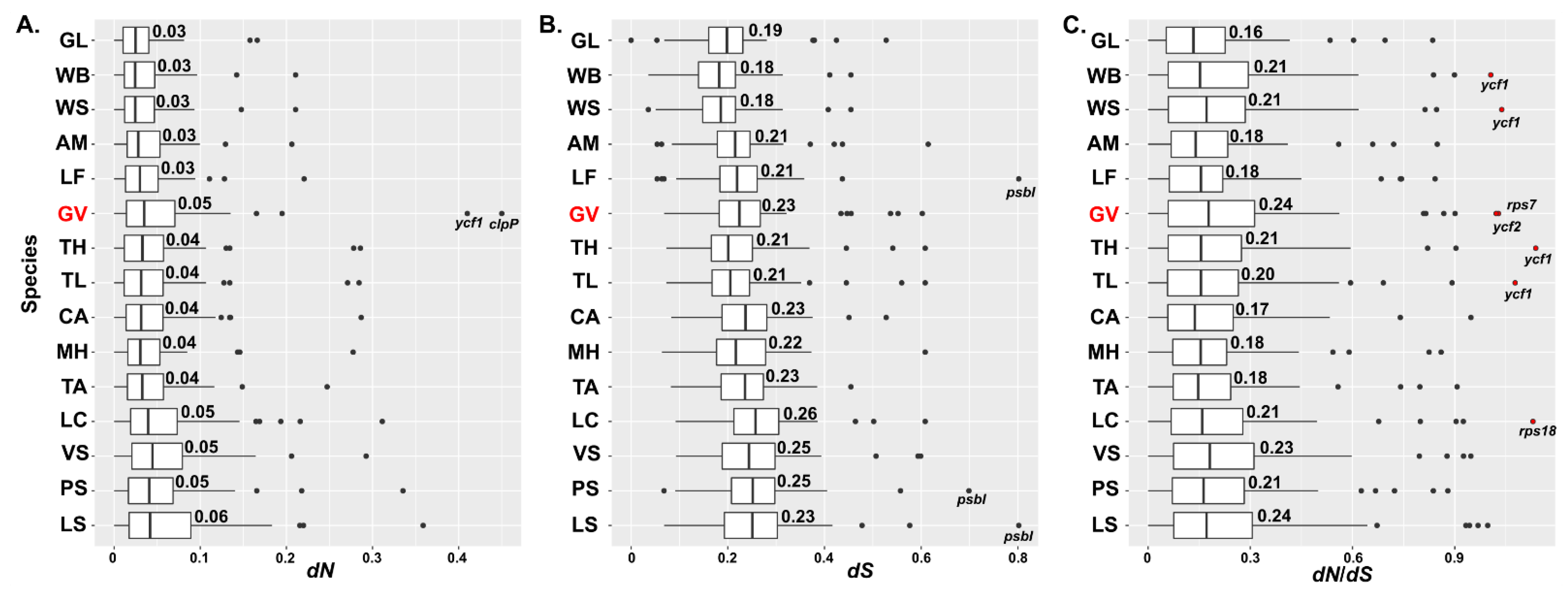
2.5. Substitution Analysis
3. Discussion
4. Materials and Methods
4.1. Sampling, DNA Extraction, and Sequencing
4.2. Genome Assembly, Genome Annotation, and Comparison of Genome Structure
4.3. Repeat Analysis
4.4. Phylogenetic Analysis
4.5. Substitution Rate Estimation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jansen, R.K.; Kaittanis, C.; Saski, C.; Lee, S.-B.; Tomkins, J.; Alverson, A.J.; Daniell, H. Phylogenetic Analyses of Vitis (Vitaceae) Based on Complete Chloroplast Genome Sequences: Effects of Taxon Sampling and Phylogenetic Methods on Resolving Relationships among Rosids. BMC Evol. Biol. 2006, 6, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhardt, N.; Brassac, J.; Kilian, B.; Blattner, F.R. Dated Tribe-Wide Whole Chloroplast Genome Phylogeny Indicates Recurrent Hybridizations within Triticeae. BMC Evol. Biol. 2017, 17, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, M.-L.; Blazier, J.C.; Govindu, M.; Jansen, R.K. Reconstruction of the Ancestral Plastid Genome in Geraniaceae Reveals a Correlation between Genome Rearrangements, Repeats, and Nucleotide Substitution Rates. Mol. Biol. Evol. 2014, 31, 645–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.S.; Ha, Y.-H.; Gil, H.-Y.; Choi, K.; Kim, D.-K.; Oh, S.-H. Two Korean Endemic Clematis Chloroplast Genomes: Inversion, Reposition, Expansion of the Inverted Repeat Region, Phylogenetic Analysis, and Nucleotide Substitution Rates. Plants 2021, 10, 397. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Cheng, T.; Zhou, S. Complete Chloroplast Genome of Sedum sarmentosum and Chloroplast Genome Evolution in Saxifragales. PLoS ONE 2013, 8, e77965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, C.; Ma, L.; Wu, Z.; Chen, K.; Wang, Y. Comparative Analyses of Chloroplast Genomes from 22 Lythraceae Species: Inferences for Phylogenetic Relationships and Genome Evolution within Myrtales. BMC Plant Biol. 2019, 19, 281. [Google Scholar] [CrossRef]
- Wu, F.-H.; Chan, M.-T.; Liao, D.-C.; Hsu, C.-T.; Lee, Y.-W.; Daniell, H.; Duvall, M.R.; Lin, C.-S. Complete Chloroplast Genome of Oncidium gower Ramsey and Evaluation of Molecular Markers for Identification and Breeding in Oncidiinae. BMC Plant Biol. 2010, 10, 68. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Yang, M.; Mo, C.; Xie, W.; Liu, C.; Wu, B.; Ma, X. Complete Chloroplast Genomes of Two Siraitia Merrill Species: Comparative Analysis, Positive Selection and Novel Molecular Marker Development. PLoS ONE 2019, 14, e0226865. [Google Scholar] [CrossRef]
- Jansen, R.K.; Cai, Z.; Raubeson, L.A.; Daniell, H.; Depamphilis, C.W.; Leebens-Mack, J.; Müller, K.F.; Guisinger-Bellian, M.; Haberle, R.C.; Hansen, A.K.; et al. Analysis of 81 Genes from 64 Plastid Genomes Resolves Relationships in Angiosperms and Identifies Genome-Scale Evolutionary Patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 19369–19374. [Google Scholar] [CrossRef] [Green Version]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The Evolution of the Plastid Chromosome in Land Plants: Gene Content, Gene Order, Gene Function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- Cai, Z.; Guisinger, M.; Kim, H.-G.; Ruck, E.; Blazier, J.C.; McMurtry, V.; Kuehl, J.V.; Boore, J.; Jansen, R.K. Extensive Reorganization of the Plastid Genome of Trifolium Subterraneum (Fabaceae) Is Associated with Numerous Repeated Sequences and Novel DNA Insertions. J. Mol. Evol. 2008, 67, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Chris Blazier, J.; Guisinger, M.M.; Jansen, R.K. Recent Loss of Plastid-Encoded Ndh Genes within Erodium (Geraniaceae). Plant Mol. Biol. 2011, 76, 263–272. [Google Scholar] [CrossRef]
- Cosner, M.E.; Jansen, R.K.; Palmer, J.D.; Downie, S.R. The Highly Rearranged Chloroplast Genome of Trachelium caeruleum (Campanulaceae): Multiple Inversions, Inverted Repeat Expansion and Contraction, Transposition, Insertions/Deletions, and Several Repeat Families. Curr. Genet. 1997, 31, 419–429. [Google Scholar] [CrossRef]
- Cosner, M.E.; Raubeson, L.A.; Jansen, R.K. Chloroplast DNA Rearrangements in Campanulaceae: Phylogenetic Utility of Highly Rearranged Genomes. BMC Evol. Biol. 2004, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-A.; Cheon, K.-S. Complete Chloroplast Genome Sequence of Adenophora racemosa (Campanulaceae): Comparative Analysis with Congeneric Species. PLoS ONE 2021, 16, e0248788. [Google Scholar] [CrossRef] [PubMed]
- Wicke, S. Genomic Evolution in Orobanchaceae. In Parasitic Orobanchaceae; Joel, D.M., Gressel, J., Musselman, L.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 267–286. ISBN 978-3-642-38145-4. [Google Scholar]
- Li, X.; Zhang, T.-C.; Qiao, Q.; Ren, Z.; Zhao, J.; Yonezawa, T.; Hasegawa, M.; Crabbe, M.J.C.; Li, J.; Zhong, Y. Complete Chloroplast Genome Sequence of Holoparasite Cistanche deserticola (Orobanchaceae) Reveals Gene Loss and Horizontal Gene Transfer from Its Host Haloxylon ammodendron (Chenopodiaceae). PLoS ONE 2013, 8, e58747. [Google Scholar] [CrossRef] [Green Version]
- Samigullin, T.H.; Logacheva, M.D.; Penin, A.A.; Vallejo-Roman, C.M. Complete Plastid Genome of the Recent Holoparasite Lathraea squamaria Reveals Earliest Stages of Plastome Reduction in Orobanchaceae. PLoS ONE 2016, 11, e0150718. [Google Scholar] [CrossRef] [Green Version]
- Azani, N.; Babineau, M.; Bailey, C.D.; Banks, H.; Barbosa, A.R.; Pinto, R.B.; Boatwright, J.S.; Borges, L.M.; Brown, G.K.; Bruneau, A.; et al. A New Subfamily Classification of the Leguminosae Based on a Taxonomically Comprehensive Phylogeny: The Legume Phylogeny Working Group (LPWG). Taxon 2017, 66, 44–77. [Google Scholar] [CrossRef] [Green Version]
- Lavin, M.; Doyle, J.J.; Palmer, J.D. Evolutionary Significance of the Loss of the Chloroplast-DNA Inverted Repeat in the Leguminosae Subfamily Papilionoideae. Evolution 1990, 44, 390–402. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L.; Palmer, J.D. Multiple Independent Losses of Two Genes and One Intron from Legume Chloroplast Genomes. Syst. Bot. 1995, 20, 272–294. [Google Scholar] [CrossRef]
- Magee, A.M.; Aspinall, S.; Rice, D.W.; Cusack, B.P.; Sémon, M.; Perry, A.S.; Stefanović, S.; Milbourne, D.; Barth, S.; Palmer, J.D.; et al. Localized Hypermutation and Associated Gene Losses in Legume Chloroplast Genomes. Genome Res. 2010, 20, 1700–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Chen, J.; Li, Y.; Liu, A.; Li, A.; Yin, M.; Shrestha, N.; Liu, J.; Ren, G. Extensive Genomic Rearrangements Mediated by Repetitive Sequences in Plastomes of Medicago and Its Relatives. BMC Plant Biol. 2021, 21, 421. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; He, X.; Song, R.; Wang, X.; Zhang, H.; Aili, R.; Chao, Y.; Shen, Y.; Yu, L.; Zhang, T.; et al. Recent Structural Variations in the Medicago Chloroplast Genomes and Their Horizontal Transfer into Nuclear Chromosomes. J. Syst. Evol. 2022, jse.12900. [Google Scholar] [CrossRef]
- Choi, I.-S.; Jansen, R.; Ruhlman, T. Lost and Found: Return of the Inverted Repeat in the Legume Clade Defined by Its Absence. Genome Biol. Evol. 2019, 11, 1321–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, F.E. Gueldenstaedtia. Mém. Soc. Nat. Moscon 1823, 6, 170. [Google Scholar]
- Sanderson, M.J.; Wojciechowski, M.F. Diversification rates in a Temperate Legume Clade: Are therer “So Many Species” of Astragalus (Fabaceae)? Am. J. Bot. 1996, 83, 1488–1502. [Google Scholar] [CrossRef]
- Zhu, X. Pollen and Seed Morphology of Gueldenstaedtia and Tibetia (Leguminosae)—with a Special Reference to the Taxonomic Significance. Nord. J. Bot. 2003, 23, 373–384. [Google Scholar] [CrossRef]
- Zhu, X. A Revision of the Genus Gueldenstaedtia (Fabaceae). Ann. Bot. Fenn. 2004, 41, 283–291. [Google Scholar]
- Duan, L.; Yang, X.; Liu, P.; Johnson, G.; Wen, J.; Chang, Z. A Molecular Phylogeny of Caraganeae (Leguminosae, Papilionoideae) Reveals Insights into New Generic and Infrageneric Delimitations. PhytoKeys 2016, 70, 111–137. [Google Scholar] [CrossRef] [Green Version]
- Moghaddam, M.; Kazempour-Osaloo, S. Extensive Survey of the Ycf4 Plastid Gene throughout the IRLC Legumes: Robust Evidence of Its Locus and Lineage Specific Accelerated Rate of Evolution, Pseudogenization and Gene Loss in the Tribe Fabeae. PLoS ONE 2020, 15, e0229846. [Google Scholar] [CrossRef]
- Wojciechowski, M.F.; Sanderson, M.J.; Hu, J.-M. Evidence on the Monophyly of Astragalus (Fabaceae) and Its Major Subgroups Based on Nuclear Ribosomal DNA ITS and Chloroplast DNA trnL Intron Data. Syst. Bot. 1999, 24, 409–437. [Google Scholar] [CrossRef]
- Lei, W.; Ni, D.; Wang, Y.; Shao, J.; Wang, X.; Yang, D.; Wang, J.; Chen, H.; Liu, C. Intraspecific and Heteroplasmic Variations, Gene Losses and Inversions in the Chloroplast Genome of Astragalus membranaceus. Sci. Rep. 2016, 6, 21669. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Xiong, Y.; He, J.; Yu, Q.; Zhao, J.; Lei, X.; Dong, Z.; Yang, J.; Peng, Y.; Zhang, X.; et al. The Complete Chloroplast Genome of Two Important Annual Clover Species, Trifolium alexandrinum and T. resupinatum: Genome Structure, Comparative Analyses and Phylogenetic Relationships with Relatives in Leguminosae. Plants 2020, 9, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Castillo-Ramírez, S.; González, V.; Bustos, P.; Luís Fernández-Vázquez, J.; Santamaría, R.I.; Arellano, J.; Cevallos, M.A.; Dávila, G. Rapid Evolutionary Change of Common Bean (Phaseolus vulgaris L.) Plastome, and the Genomic Diversification of Legume Chloroplasts. BMC Genom. 2007, 8, 228. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.K.; Wojciechowski, M.F.; Sanniyasi, E.; Lee, S.-B.; Daniell, H. Complete Plastid Genome Sequence of the Chickpea (Cicer arietinum) and the Phylogenetic Distribution of rps12 and clpP Intron Losses among Legumes (Leguminosae). Mol. Phylogenet. Evol. 2008, 48, 1204–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabir, J.; Schwarz, E.; Ellison, N.; Zhang, J.; Baeshen, N.A.; Mutwakil, M.; Jansen, R.; Ruhlman, T. Evolutionary and Biotechnology Implications of Plastid Genome Variation in the Inverted-Repeat-Lacking Clade of Legumes. Plant Biotechnol. J. 2014, 12, 743–754. [Google Scholar] [CrossRef]
- Moghaddam, M.; Ohta, A.; Shimizu, M.; Terauchi, R.; Kazempour-Osaloo, S. The Complete Chloroplast Genome of Onobrychis gaubae (Fabaceae-Papilionoideae): Comparative Analysis with Related IR-Lacking Clade Species. BMC Plant Biol. 2022, 22, 75. [Google Scholar] [CrossRef]
- Erixon, P.; Oxelman, B. Whole-Gene Positive Selection, Elevated Synonymous Substitution Rates, Duplication, and Indel Evolution of the Chloroplast clpP1 Gene. PLoS ONE 2008, 3, e1386. [Google Scholar] [CrossRef] [Green Version]
- Wallace, R.S.; Cota, J.H. An Intron Loss in the Chloroplast Gene rpoC1 Supports a Monophyletic Origin for the Subfamily Cactoideae of the Cactaceae. Curr. Genet. 1996, 29, 275–281. [Google Scholar] [CrossRef]
- Downie, S.R.; Llanas, E.; Katz-Downie, D.S. Multiple Independent Losses of the rpoC1 Intron in Angiosperm Chloroplast DNA’s. Syst. Bot. 1996, 21, 135. [Google Scholar] [CrossRef]
- Hiratsuka, J.; Shimada, H.; Whittier, R.; Ishibashi, T.; Sakamoto, M.; Mori, M.; Kondo, C.; Honji, Y.; Sun, C.-R.; Meng, B.-Y.; et al. The Complete Sequence of the Rice (Oryza sativa) Chloroplast Genome: Intermolecular Recombination between DistincttRNAGenes Accounts for a Major Plastid DNA Inversion during the Evolution of the Cereals. Molec. Gen. Genet. 1989, 217, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.F.; Zanis, M.J.; Emery, N.C. Comparative Analysis of Complete Chloroplast Genome Sequence and Inversion Variation in Lasthenia burkei (Madieae, Asteraceae). Am. J. Bot. 2014, 101, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, S.M.; Bernardo Carvalho, A.; Clark, A.G. Local Rates of Recombination Are Positively Correlated with GC Content in the Human Genome. Mol. Biol. Evol. 2001, 18, 1139–1142. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.-J.; Cheng, C.-L.; Chang, C.-C.; Wu, C.-L.; Su, T.-M.; Chaw, S.-M. Dynamics and Evolution of the Inverted Repeat-Large Single Copy Junctions in the Chloroplast Genomes of Monocots. BMC Evol. Biol. 2008, 8, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinn, B.T.; Sedmak, D.D.; Kelly, L.M.; Freudenstein, J.V. Total Duplication of the Small Single Copy Region in the Angiosperm Plastome: Rearrangement and Inverted Repeat Instability in Asarum. Am. J. Bot. 2018, 105, 71–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, L.; Wen, J.; Yang, X.; Liu, P.-L.; Arslan, E.; Ertuğrul, K.; Chang, Z.-Y. Phylogeny of Hedysarum and Tribe Hedysareae (Leguminosae: Papilionoideae) Inferred from Sequence Data of ITS, MatK, TrnL-F and PsbA-TrnH. Taxon 2015, 64, 49–64. [Google Scholar] [CrossRef]
- Wolfe, K.H.; Li, W.H.; Sharp, P.M. Rates of Nucleotide Substitution Vary Greatly among Plant Mitochondrial, Chloroplast, and Nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [Green Version]
- Perry, A.; Wolfe, K. Nucleotide Substitution Rates in Legume Chloroplast DNA Depend on the Presence of the Inverted Repeat. J. Mol. Evol. 2002, 55, 501–508. [Google Scholar] [CrossRef]
- Schwarz, E.N.; Ruhlman, T.A.; Weng, M.-L.; Khiyami, M.A.; Sabir, J.S.M.; Hajarah, N.H.; Alharbi, N.S.; Rabah, S.O.; Jansen, R.K. Plastome-Wide Nucleotide Substitution Rates Reveal Accelerated Rates in Papilionoideae and Correlations with Genome Features Across Legume Subfamilies. J. Mol. Evol. 2017, 84, 187–203. [Google Scholar] [CrossRef]
- Shrestha, B.; Weng, M.-L.; Theriot, E.C.; Gilbert, L.E.; Ruhlman, T.A.; Krosnick, S.E.; Jansen, R.K. Highly Accelerated Rates of Genomic Rearrangements and Nucleotide Substitutions in Plastid Genomes of Passiflora Subgenus Decaloba. Mol. Phylogenet. Evol. 2019, 138, 53–64. [Google Scholar] [CrossRef]
- Claude, S.-J.; Park, S.; Park, S. Gene Loss, Genome Rearrangement, and Accelerated Substitution Rates in Plastid Genome of Hypericum Ascyron (Hypericaceae). BMC Plant Biol. 2022, 22, 135. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; dePamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A Fast and Versatile Toolkit for Accurate de Novo Assembly of Organelle Genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq–Versatile and Accurate Annotation of Organelle Genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. TRNAscan-SE 2.0: Improved Detection and Functional Classification of Transfer RNA Genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) Version 1.3.1: Expanded Toolkit for the Graphical Visualization of Organellar Genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The Manifold Applications of Repeat Analysis on a Genomic Scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [Green Version]
- Xia, M.-Q.; Liao, R.-Y.; Zhou, J.-T.; Lin, H.-Y.; Li, J.-H.; Li, P.; Fu, C.-X.; Qiu, Y.-X. Phylogenomics and Biogeography of Wisteria: Implications on Plastome Evolution among Inverted Repeat-Lacking Clade (IRLC) Legumes. J. Syst. Evol. 2022, 60, 253–265. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Genome Size (bp) | GC Contents | Gene | Number of Repeat a (F/R/C/P) b | Length of Total Repeats (bp) | Repeats Percentage (%) | ||
|---|---|---|---|---|---|---|---|---|
| Coding Genes | tRNA | rRNA | ||||||
| Lotus japonicus | 150,519 | 36.0% | 78 | 30 | 4 | 61 (26/3/2/33) | 2834 | 1.8% |
| Glycyrrhiza lepidota | 127,939 | 34.2% | 77 | 30 | 4 | 92 (59/4/4/25) | 4315 | 3.3% |
| Wisteria sinensis | 130,561 | 34.4% | 77 | 30 | 4 | 109 (70/14/0/25) | 4622 | 3.5% |
| Wisteria brachybotrys | 131,179 | 34.4% | 77 | 30 | 4 | 89 (50/11/2/26) | 3615 | 2.7% |
| Astragalus mongholicus var. nakaianus | 123,633 | 34.1% | 77 | 30 | 4 | 67 (37/7/1/22) | 2597 | 2.1% |
| Lessertia frutescens | 122,700 | 34.2% | 77 | 30 | 4 | 35 (16/3/0/16) | 1394 | 1.1% |
| Gueldenstaedtia verna | 122,569 | 36.0% | 77 | 30 | 4 | 74 (51/0/0/23) | 3727 | 3.0% |
| Tibetia himalaica | 124,201 | 34.5% | 77 | 30 | 4 | 236 (227/1/0/8) | 11,917 | 9.5% |
| Tibetia liangshanensis | 122,372 | 34.7% | 77 | 30 | 4 | 93 (82/1/0/10) | 4260 | 3.4% |
| Cicer arietinum | 125,319 | 33.9% | 76 | 30 | 4 | 75 (45/4/1/25) | 3548 | 2.8% |
| Medicago hybrida | 125,208 | 33.8% | 76 | 30 | 4 | 105 (80/5/0/20) | 4062 | 3.2% |
| Trifolium aureum | 126,970 | 34.6% | 77 | 30 | 4 | 51 (34/3/0/14) | 2834 | 2.2% |
| Lens culinaris | 122,967 | 34.4% | 75 | 30 | 4 | 105 (89/0/0/16) | 4561 | 3.7% |
| Vicia sativa | 122,467 | 35.2% | 76 | 30 | 4 | 78 (65/0/1/12) | 6004 | 4.9% |
| Pisum sativum | 122,169 | 34.8% | 75 | 30 | 4 | 61 (54/1/0/6) | 2564 | 2.0% |
| Lathyrus sativus | 121,020 | 35.1% | 76 | 30 | 4 | 78 (50/0/0/28) | 3343 | 2.7% |
| LJ | GL | WB | WS | AM | LF | GV | TH | TL | CA | MH | TA | LC | VS | PS | LS | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene loss | ||||||||||||||||
| rps16 | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x |
| rps18 | o | o | o | o | o | o | o | o | o | o | o | o | x | o | o | o |
| rpl23 | o | o | o | o | o | o | o | o | o | o | o | o | o | x | x | x |
| atpE | o | o | o | o | x | o | o | o | o | o | o | o | o | o | o | o |
| ycf4 | o | o | o | o | o | o | o | o | o | o | x | o | x | x | x | x |
| Number of introns | ||||||||||||||||
| atpF | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| clpP | 2 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| ndhA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| ndhB | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| petB | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| petD | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| rpl2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| rpl16 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 |
| rps12 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| rpoC1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| ycf3 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| trnG-UCC | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 |
| trnL-UAA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| trnV-UAC | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| trnK-UUU | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| trnI-GAU | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| trnA-UGC | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, O.; Choi, K.S. Characterization of the Chloroplast Genome Structure of Gueldenstaedtia verna (Papilionoideae) and Comparative Analyses among IRLC Species. Forests 2022, 13, 1942. https://doi.org/10.3390/f13111942
Son O, Choi KS. Characterization of the Chloroplast Genome Structure of Gueldenstaedtia verna (Papilionoideae) and Comparative Analyses among IRLC Species. Forests. 2022; 13(11):1942. https://doi.org/10.3390/f13111942
Chicago/Turabian StyleSon, Ogyeong, and Kyoung Su Choi. 2022. "Characterization of the Chloroplast Genome Structure of Gueldenstaedtia verna (Papilionoideae) and Comparative Analyses among IRLC Species" Forests 13, no. 11: 1942. https://doi.org/10.3390/f13111942