Early Signs of the Effects of Forest Fragmentation on the Genetic Diversity and Structure of the Threatened Ecuadorian Tree Ocotea rotundata (Lauraceae)




Abstract
:1. Introduction
2. Materials and Methods
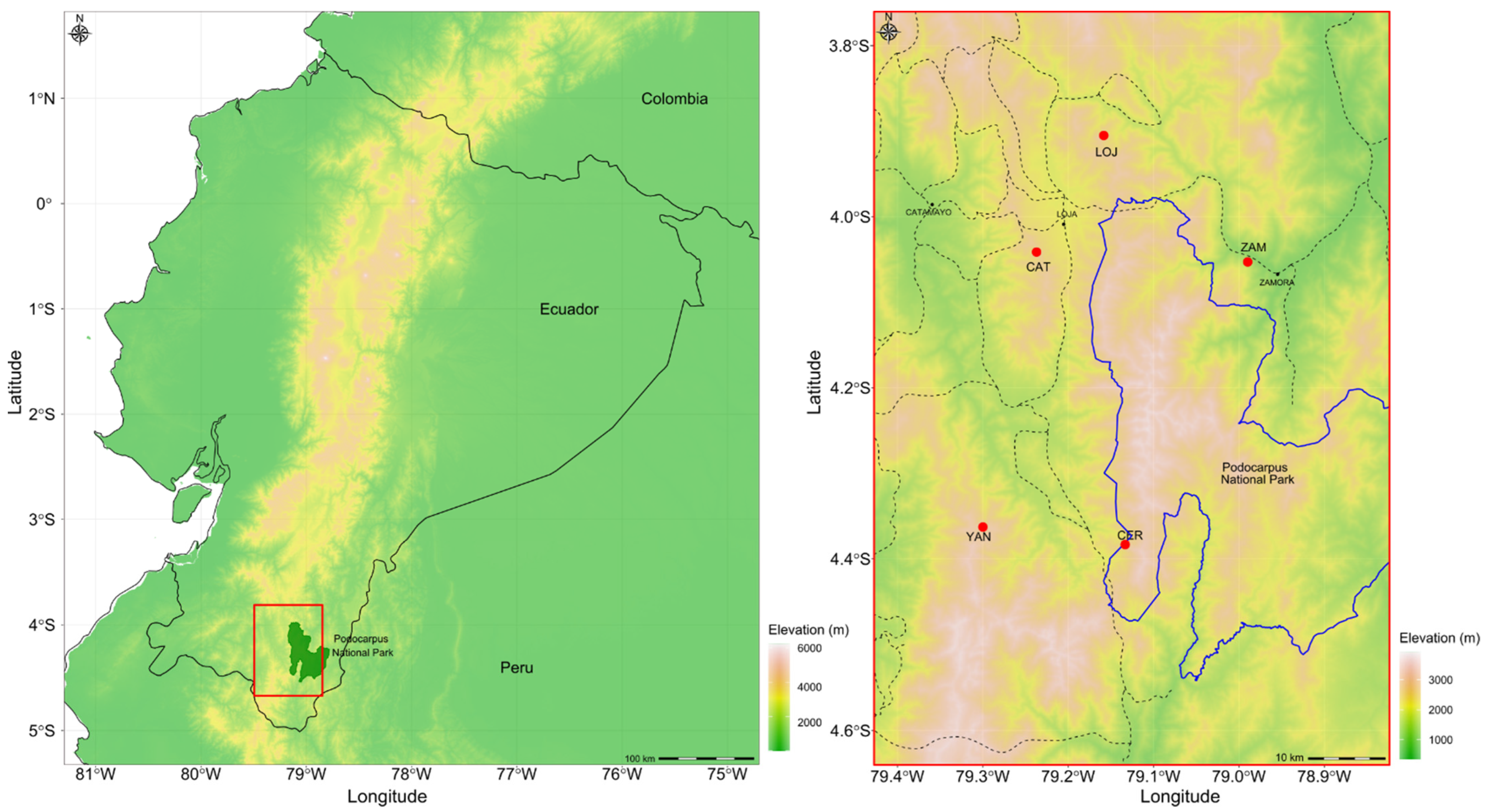
2.1. Population Sampling
2.2. DNA Extraction and Development of nSSRs
2.3. nSSR Amplification
2.4. Genetic Diversity and Differentiation
2.5. Genetic and Spatial Population Structure
3. Results
3.1. Diversity between Loci
3.2. Genetic Diversity and Differentiation in Ocotea rotundata
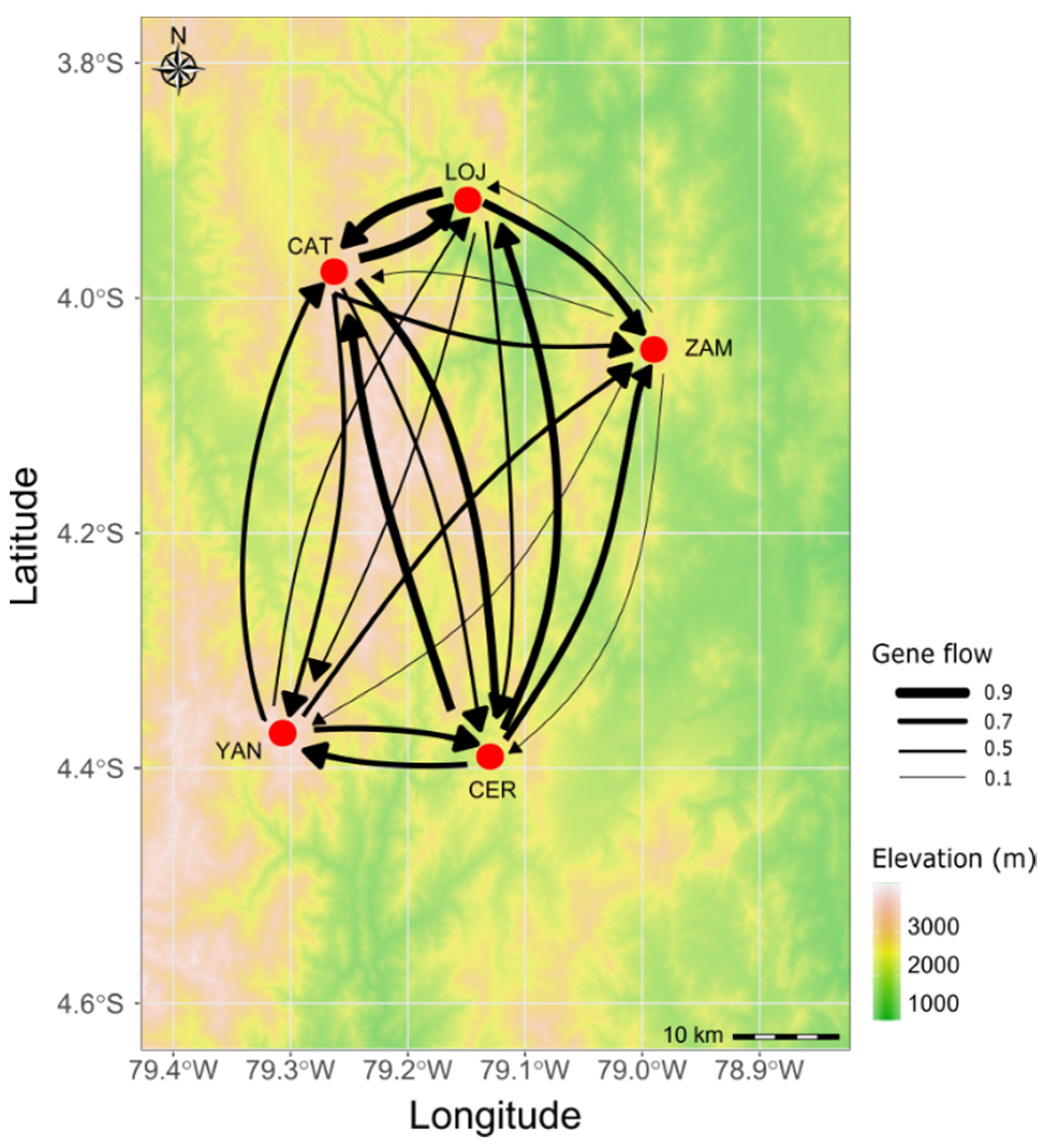
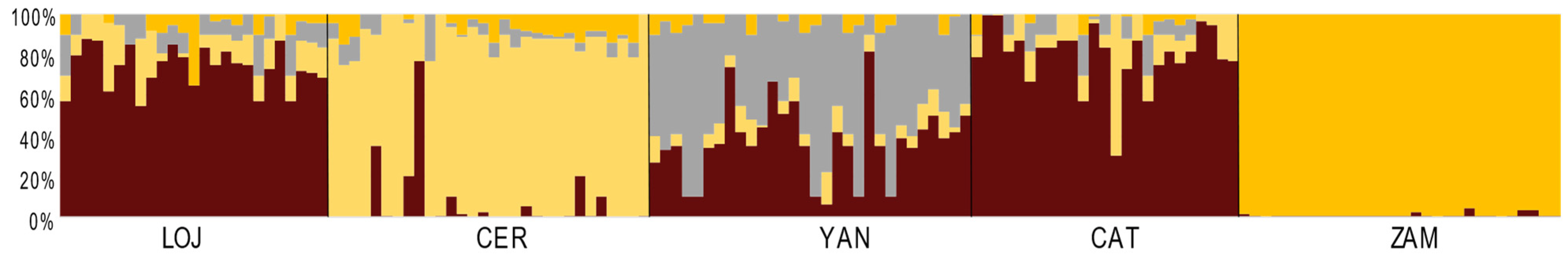
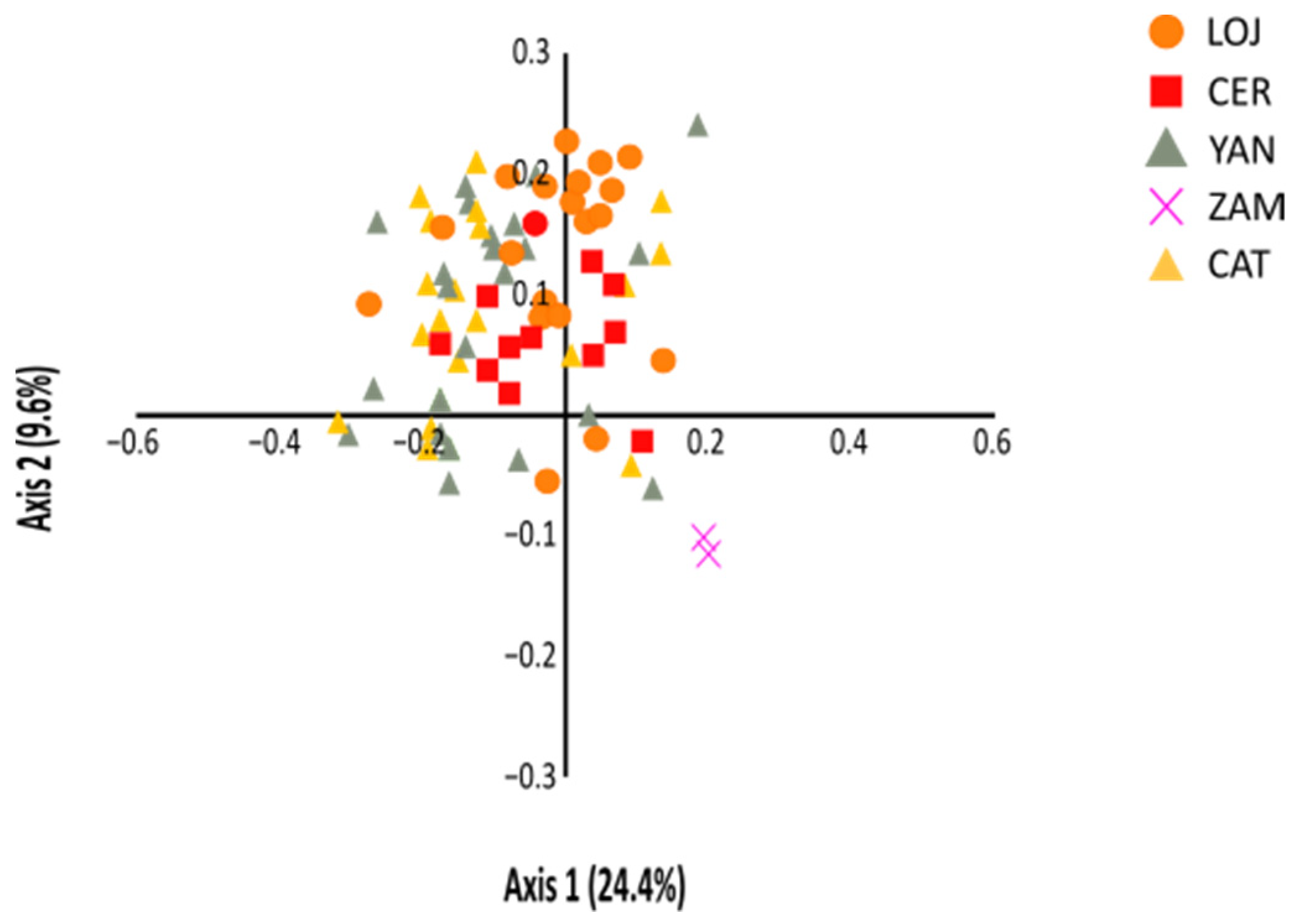
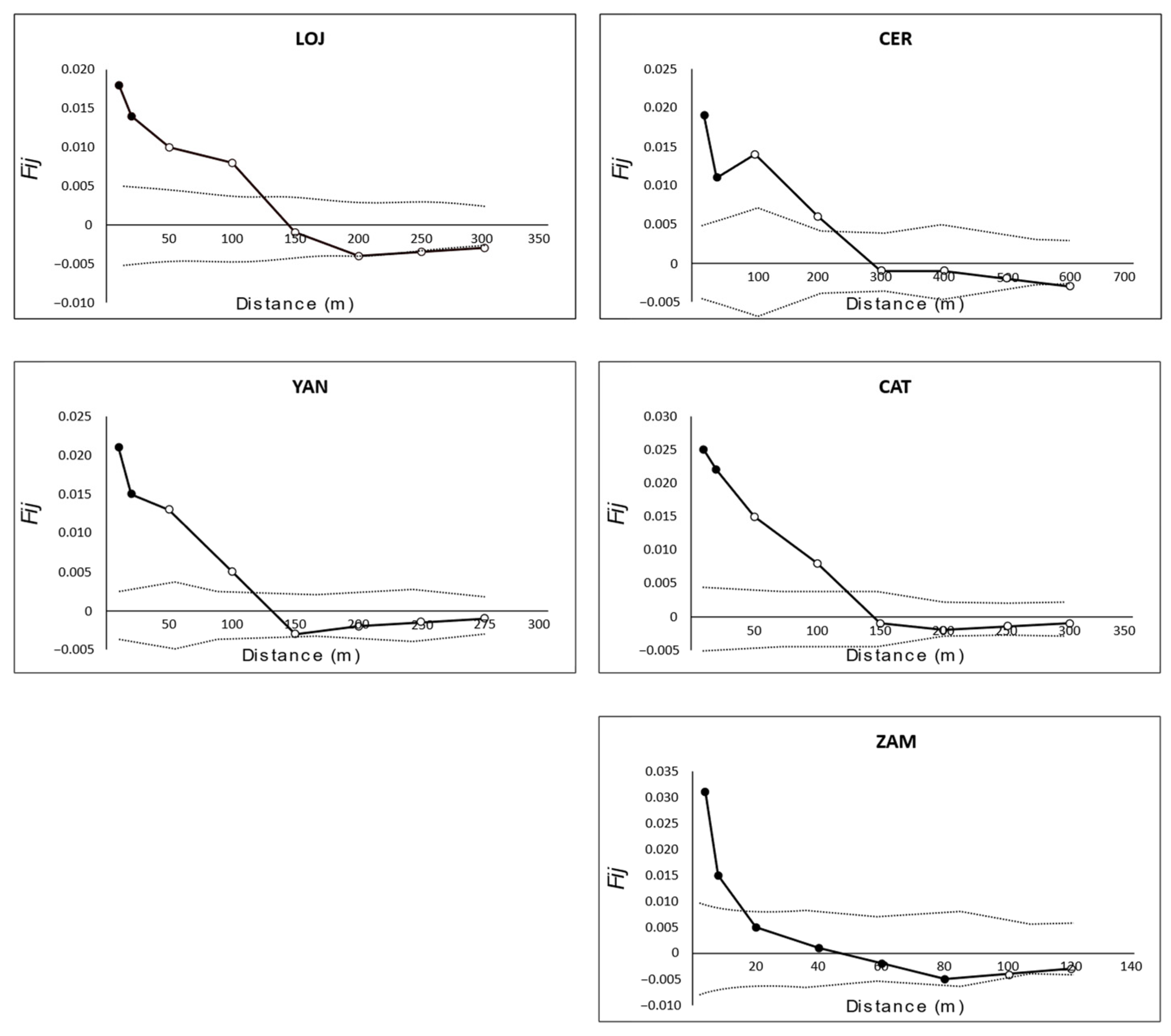
3.3. Genetic and Spatial Population Structure
4. Discussion
4.1. High Genetic Diversity in Ocotea rotundata
4.2. Low Differentiation and High Gene Flow between (Most) Populations
4.3. Genetic Insights for Conservation Actions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isabel, N.; Holliday, J.A.; Aitken, S.N. Forest genomics: Advancing climate adaptation, forest health, productivity, and conservation. Evol. Appl. 2020, 13, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allendorf, F.W.; Hohenlohe, P.A.; Luikart, G. Genomics and the future of conservation genetics. Nat. Rev. Genet. 2010, 11, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene Flow and Local Adaptation in Trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Vanhove, M.; Pina-Martins, F.; Coelho, A.C.; Branquinho, C.; Costa, A.; Batista, D.; Príncipe, A.; Sousa, P.; Henriques, A.; Marques, I.; et al. Using gradient Forest to predict climate response and adaptation in Cork oak. J. Evol. Biol. 2021, 34, 910–923. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, F.; Koskela, J.; Hubert, J.; Kraigher, H.; Longauer, R.; Olrik, D.C.; Schüler, S.; Bozzano, M.; Alizoti, P.; Bakys, R.; et al. Dynamic Conservation of Forest Genetic Resources in 33 European Countries. Conserv. Biol. 2013, 27, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Wu, B.; Wang, X.; Yao, Z.; Li, Y.; Liu, Y. Scale-dependent effects of habitat fragmentation on the genetic diversity of Actinidia chinensis populations in China. Hortic. Res. 2020, 7, 172. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Peñuelas, J. Genetic effects of chronic habitat fragmentation in a wind-pollinated tree. Proc. Natl. Acad. Sci. USA 2006, 103, 8096–8100. [Google Scholar] [CrossRef] [Green Version]
- Plomion, C.; Aury, J.-M.; Amselem, J.; Leroy, T.; Murat, F.; Duplessis, S.; Faye, S.; Francillonne, N.; Labadie, K.; Le Provost, G.; et al. Oak genome reveals facets of long lifespan. Nat. Plants 2018, 4, 440–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noreen, A.M.E.; Webb, E.L. High Genetic Diversity in a Potentially Vulnerable Tropical Tree Species Despite Extreme Habitat Loss. PLoS ONE 2013, 8, 82632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ony, M.A.; Nowicki, M.; Boggess, S.L.; Klingeman, W.E.; Zobel, J.M.; Trigiano, R.N.; Hadziabdic, D. Habitat fragmentation influences genetic diversity and differentiation: Fine-scale population structure of Cercis canadensis (eastern redbud). Ecol. Evol. 2020, 10, 3655–3670. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.L.; Wickneswari, R.; Mahani, M.C.; Zakri, A.H. Genetic Diversity of a Tropical Tree Species, Shorea leprosula Miq. (Dipterocarpaceae), In Malaysia: Implications for Conservation of Genetic Resources and Tree Improvement. Biotropica 2000, 32, 213–224. [Google Scholar] [CrossRef]
- Li, Z.; Wang, H.; Liao, P.-C.; Wang, C.-N.; Du, F.K.; Su, J.; Yan, Y.; Song, J.; Li, J.; Mao, J.; et al. Recent Fragmentation May Not Alter Genetic Patterns in Endangered Long-Lived Species: Evidence from Taxus cuspidata. Front. Plant Sci. 2018, 9, 1571. [Google Scholar] [CrossRef]
- Piotti, A.; Leonardi, S.; Buiteveld, J.; Geburek, T.; Gerber, S.; Kramer, K.; Vettori, C.; Vendramin, G.G. Comparison of pollen gene flow among four European beech (Fagus sylvatica L.) populations characterized by different management regimes. Heredity 2012, 108, 322–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provan, J.; Beatty, G.E.; Hunter, A.M.; McDonald, R.A.; McLaughlin, E.; Preston, S.J.; Wilson, S. Restricted gene flow in fragmented populations of a wind-pollinated tree. Conserv. Genet. 2008, 9, 1521–1532. [Google Scholar] [CrossRef]
- Hensen, I.; Cierjacks, A.; Hirsch, H.; Kessler, M.; Romoleroux, K.; Renison, D.; Wesche, K. Historic and recent fragmentation coupled with altitude affect the genetic population structure of one of the world’s highest tropical tree line species. Glob. Ecol. Biogeogr. 2012, 21, 455–464. [Google Scholar] [CrossRef]
- Leonardi, S.; Piovani, P.; Scalfi, M.; Piotti, A.; Giannini, R.; Menozzi, P. Effect of Habitat Fragmentation on the Genetic Diversity and Structure of Peripheral Populations of Beech in Central Italy. J. Hered. 2012, 103, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Finch, K.N.; Jones, F.A.; Cronn, R.C. Genomic resources for the Neotropical tree genus Cedrela (Meliaceae) and its relatives. BMC Genom. 2019, 20, 58. [Google Scholar] [CrossRef] [Green Version]
- Burley, J.T.; Kellner, J.R.; Hubbell, S.P.; Faircloth, B.C. Genome assemblies for two Neotropical trees: Jacaranda copaia and Handroanthus guayacan. G3 2021, 11, jkab010. [Google Scholar] [CrossRef]
- García-Barrios, L.; Galván-Miyoshi, Y.M.; Valdivieso-Pérez, I.A.; Masera, O.R.; Bocco, G.; Vandermeer, J. Neotropical Forest Conservation, Agricultural Intensification, and Rural Out-migration: The Mexican Experience. Bioscience 2009, 59, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Tapia-Armijos, M.F.; Homeier, J.; Espinosa, C.I.; Leuschner, C.; De La Cruz, M. Deforestation and forest fragmentation in south Ecuador since the 1970s—Losing a hotspot of biodiversity. PLoS ONE 2015, 10, e0133701. [Google Scholar] [CrossRef]
- Tapia-Armijos, M.F.; Homeier, J.; Draper Munt, D. Spatio-temporal analysis of the human footprint in South Ecuador: Influence of human pressure on ecosystems and effectiveness of protected areas. Appl. Geogr. 2017, 78, 22–32. [Google Scholar] [CrossRef]
- Rivas, C.A.; Guerrero-Casado, J.; Navarro-Cerillo, R.M. Deforestation and fragmentation trends of seasonal dry tropical forest in Ecuador: Impact on conservation. For. Ecosyst. 2021, 8, 46. [Google Scholar] [CrossRef]
- Kleemann, J.; Zamora, C.; Villacis-Chiluisa, A.B.; Cuenca, P.; Koo, H.; Noh, J.K.; Fürst, C.; Thiel, M. Deforestation in Continental Ecuador with a Focus on Protected Areas. Land 2022, 11, 268. [Google Scholar] [CrossRef]
- Ruilova, X.R.; Marques, I. Better common than rare? Effects of low reproductive success, scarce pollinator visits and interspecific gene flow in threatened and common species of Tibouchina (Melastomataceae). Plant Species Biol. 2016, 31, 288–295. [Google Scholar] [CrossRef]
- Ocotea rotundata. Available online: https://www.iucnredlist.org/species/38101/10096866 (accessed on 11 August 2022).
- van der Werff, H. A Synopsis of Ocotea (Lauraceae) in Central America and Southern Mexico. Ann. Missouri Bot. Gard. 2002, 89, 429–451. [Google Scholar] [CrossRef]
- Van Der Werff, H. Studies in andean ocotea (Lauraceae) II. Species with hermaphrodite flowers and densely pubescent lower leaf surfaces, occurring above 1000 meters in altitude. Novon 2013, 22, 336–370. [Google Scholar] [CrossRef]
- van der Werff, H. New Species of Lauraceae from Ecuador and Peru. Ann. Missouri Bot. Gard. 1991, 78, 409. [Google Scholar] [CrossRef]
- Palacios, W.A. Two new species of Lauraceae from Ecuador. Phytotaxa 2018, 346, 180–188. [Google Scholar] [CrossRef]
- Gilardoni, G.; Montalván, M.; Vélez, M.; Malagón, O. Chemical and Enantioselective Analysis of the Essential Oils from Different Morphological Structures of Ocotea quixos (Lam.) Kosterm. Plants 2021, 10, 2171. [Google Scholar] [CrossRef] [PubMed]
- Martins, E.M.; Lamont, R.W.; Martinelli, G.; Lira-Medeiros, C.F.; Quinet, A.; Shapcott, A. A Genetic diversity and population genetic structure in three threatened Ocotea species (Lauraceae) from Brazil’s Atlantic Rainforest and implications for their conservation. Conserv. Genet. 2015, 16, 1–14. [Google Scholar] [CrossRef]
- Danieli-Silva, A.; VARASSIN, I.G. Breeding system and thrips (Thysanoptera) pollination in the endangered tree Ocotea porosa (Lauraceae): Implications for conservation. Plant Species Biol. 2013, 28, 31–40. [Google Scholar] [CrossRef]
- Richard, J. The Effect of Logging and Invasion of Acacia mearnsii on Regeneration of Ocotea usambarensis in Chome Nature Reserve, Tanzania. Ph.D. Thesis, Sokoine University of Agriculture, Morogoro, Tanzania, 2016. [Google Scholar]
- Nsolomo, V.R.; Venn, K. Capacity of fungi to colonise wood of the East African camphor tree, Ocotea usambarensis. Mycol. Res. 2000, 104, 1468–1472. [Google Scholar] [CrossRef]
- Aguirre-Mendoza, Z.; Loja, Á.; Solano, C.; Aguirre, N. Especies Forestales mas Aprovechadas del sur del Ecuador; Biblioteca Hernán Malo González: Cuenca, Ecuador, 2015; ISBN 9788578110796. [Google Scholar]
- Radice, M.; Escobar, J.A.; Pérez Martínez, A. Ocotea quixos Essential oil: A Systematic Review about the Ethno-Medicinal Uses, Phytochemistry and Biological Activity. In MOL2NET 2017, International Conference on Multidisciplinary Sciences, 3rd ed.; Multidisciplinary Digital Publishing Institute: Basel, Switzerland, 2017; Volume 3. [Google Scholar] [CrossRef]
- Vallejos, P.Q.; Veit, P.G.; Tipula, P.; Reytar, K. Undermining Rights: Indigenous Lands and Mining in the Amazon. World Resour. Inst. 2020. [Google Scholar] [CrossRef]
- Roy, B.A.; Zorrilla, M.; Endara, L.; Thomas, D.C.; Vandegrift, R.; Rubenstein, J.M.; Policha, T.; Ríos-Touma, B.; Read, M. New Mining Concessions Could Severely Decrease Biodiversity and Ecosystem Services in Ecuador Introduction New Mining Concessions in Ecuador. Trop. Conserv. Sci. 2018, 11, 1940082918780427. [Google Scholar] [CrossRef]
- Curatola Fernández, G.F.; Obermeier, W.A.; Gerique, A.; López Sandoval, M.F.; Lehnert, L.W.; Thies, B.; Bendix, J. Land cover change in the Andes of southern Ecuador-Patterns and drivers. Remote Sens. 2015, 7, 2509–2542. [Google Scholar] [CrossRef] [Green Version]
- Ecuador|EU REDD Facility. Available online: https://euredd.efi.int/countries/ecuador/ (accessed on 18 October 2022).
- Kõressaar, T.; Lepamets, M.; Kaplinski, L.; Raime, K.; Andreson, R.; Remm, M. Primer3_masker: Integrating masking of template sequence with primer design software. Bioinformatics 2018, 34, 1937–1938. [Google Scholar] [CrossRef] [Green Version]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinform. Appl. 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Flamenco-Sandoval, A.; Martínez Ramos, M.; Masera, O.R. Assessing implications of land-use and land-cover change dynamics for conservation of a highly diverse tropical rain forest. Biol. Conserv. 2007, 138, 131–145. [Google Scholar] [CrossRef]
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Rice, W.R. Analyzing Tables of Statistical Tests. Evolution 1989, 43, 223. [Google Scholar] [CrossRef]
- Wright, S. The Interpretation of Population Structure By F-Statistics with Special Regard to Systems of Mating. Evolution 1965, 19, 395–420. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (Version 3.0): An Integrated Software Package for Population Genetics Data Analysis. Evol. Bioinform. Online 2005, 1, 117693430500100000. [Google Scholar] [CrossRef] [Green Version]
- Yeh, F.C.; Yang, R.-C. POPGENE Version 1.31 Microsoft Window-Based Freeware for Population Genetic Analysis. Quick User Guide, August 1999. Available online: https://sites.ualberta.ca/~fyeh/popgene.pdf (accessed on 18 October 2022).
- Alcala, N.; Goudet, J.; Vuilleumier, S. On the transition of genetic differentiation from isolation to panmixia: What we can learn from G S T and D. Theor. Popul. Biol. 2014, 93, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Sundqvist, L.; Keenan, K.; Zackrisson, M.; Prodöhl, P.; Kleinhans, D. Directional genetic differentiation and relative migration. Ecol. Evol. 2016, 6, 3461–3475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, K.; Mcginnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. diveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, O.J.; Vekemans, X. SPAGeDi: A Versatile Computer Program to Analyse Spatial Genetic Structure at the Individual or Population Levels. Molecular Ecology Notes 2002, 2, 618–620. [Google Scholar] [CrossRef] [Green Version]
- Loiselle, B.A.; Sork, V.L.; Nason, J.; Graham, C. Spatial genetic structure of a tropical understory shrub, Psychotria officinalis (Rubiaceae). Am. J. Bot. 1995, 82, 1420–1425. [Google Scholar] [CrossRef]
- Vekemans, X.; Hardy, O.J. New insights from fine-scale spatial genetic structure analyses in plant populations. Mol. Ecol. 2004, 13, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Geng, Q.; Sun, L.; Zhang, P.; Wang, Z.; Qiu, Y.; Liu, H.; Lian, C. Understanding population structure and historical demography of Litsea auriculata (Lauraceae), an endangered species in east China. OPEN. Sci. Rep. 2017, 7, 17343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, B.; Vu, D.D.; Vu, D.G.; Bui, T.T.X.; Rahman, S.U.; Pham, M.P.; Nguyen, M.T.; Nguyen, V.S.; Shah, S.N.M.; Tran, V.H. Genetic diversity and population structure of Cinnamomum balansae Lecomte inferred by microsatellites. Open Life Sci. 2022, 17, 323–332. [Google Scholar] [CrossRef]
- Senkoro, A.M.; Talhinhas, P.; Simões, F.; Batista-Santos, P.; Shackleton, C.M.; Voeks, R.A.; Marques, I.; Ribeiro-Barros, A.I. The genetic legacy of fragmentation and overexploitation in the threatened medicinal African pepper-bark tree, Warburgia salutaris. Sci. Rep. 2020, 10, 19725. [Google Scholar] [CrossRef]
- Cueva-Agila, A.; Vélez-Mora, D.; Arias, D.; Curto, M.; Meimberg, H.; Brinegar, C. Genetic characterization of fragmented populations of Cinchona officinalis L. (Rubiaceae), a threatened tree of the northern Andean cloud forests. Tree Genet. Genomes 2019, 15, 81. [Google Scholar] [CrossRef] [Green Version]
- Lundholm, J.; Johnson, A.L.; Rivkin, L.R.; Ksiazek-Mikenas, K.; Fant, J.B.; Skogen, K.A. Pollinator-Mediated Gene Flow Connects Green Roof Populations Across the Urban Matrix: A Paternity Analysis of the Self-Compatible Forb Penstemon hirsutus. Front. Ecol. Evol. 2019, 1, 299. [Google Scholar] [CrossRef] [Green Version]
- Solórzano, S.; Arias, S.; Dávila, P. Genetics and Conservation of Plant Species of Extremely Narrow Geographic Range. Diversity 2016, 8, 31. [Google Scholar] [CrossRef]
- Lowe, A.; Boshier, D.; Ward, M.; Bacles, C.; Navarro, C. Genetic resource impacts of habitat loss and degradation; reconciling empirical evidence and predicted theory for neotropical trees. Heredity 2005, 95, 255–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alleaume-Benharira, M.; Pen, I.R.; Ronce, O. Geographical patterns of adaptation within a species’ range: Interactions between drift and gene flow. J. Evol. Biol. 2006, 19, 203–215. [Google Scholar] [CrossRef]
- Fischer, M.; Matthies, D. RAPD variation in relation to population size and plant fitness in the rare Gentianella germanica (Gentianaceae). Am. J. Bot. 1998, 85, 811. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, D.R.; Braschler, B.; Rusterholz, H.-P.; Baur, B. Genetic effects of anthropogenic habitat fragmentation on remnant animal and plant populations: A meta-analysis. Ecosphere 2018, 9, e02488. [Google Scholar] [CrossRef]
- Shi, X.; Wen, Q.; Cao, M.; Guo, X.; Xu, L.A. Genetic diversity and structure of natural Quercus variabilis population in China as revealed by microsatellites markers. Forests 2017, 8, 495. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.-A.; Li, X.-J.; Pan, H.-X.; Zou, H.-Y.; Yin, T.-M.; Hunag, M.-R. Study on Population Genetic Structure in Castanopsis Fargesii with Microsatellite Markers. J. Integr. Plant Biol. 2001, 43, 409–412. [Google Scholar]
- Pometti, C.; Bessega, C.; Cialdella, A.; Ewens, M.; Saidman, B.; Vilardi, J. Spatial genetic structure within populations and management implications of the South American species Acacia aroma (Fabaceae). PLoS ONE 2018, 13, e0192107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclerc, T.; Vimal, R.; Troispoux, V.; Périgon, S.; Scotti, I. Life after disturbance (I): Changes in the spatial genetic structure of Jacaranda copaia (Aubl.) D. Don (Bignonianceae) after logging in an intensively studied plot in French Guiana. Ann. For. Sci. 2015, 72, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Zhang, X.; Guansong, Y.; Yang, L.; Wang, Y.; Shen, S. Biological characteristics and conservation genetics of the narrowly distributed rare plant Cinnamomum chago (Lauraceae). Plant Divers. 2016, 38, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, G.M.; Boshier, D.H.; Powell, W. Increased pollen flow counteracts fragmentation in a tropical dry forest: An example from Swietenia humilis Zuccarini. Proc. Natl. Acad. Sci. USA 2002, 99, 2038–2042. [Google Scholar] [CrossRef] [Green Version]
- Dick, C.W. Genetic rescue of remnant tropical trees by an alien pollinator. Proc. R. Soc. B Boil. Sci. 2001, 268, 2391–2396. [Google Scholar] [CrossRef]
- Liepelt, S.; Bialozyt, R.; Ziegenhagen, B. Wind-dispersed pollen mediates postglacial gene flow among refugia. Proc. Natl. Acad. Sci. USA 2002, 99, 14590–14594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Ma, Y.; Sun, W.; Wen, X.; Milne, R. High Genetic Diversity and Low Differentiation of Michelia coriacea (Magnoliaceae), a Critically Endangered Endemic in Southeast Yunnan, China. Int. J. Mol. Sci 2012, 13, 4396–4411. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.R.S.; Albuquerque, P.S.B.; Ervedosa, F.R.; Mota, J.W.S.; Figueira, A.; Sebbenn, A.M. Understanding the genetic diversity, spatial genetic structure and mating system at the hierarchical levels of fruits and individuals of a continuous Theobroma cacao population from the Brazilian Amazon. Heredity 2011, 106, 973–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, A.L.; García, M.V.; Heuertz, M.; González-Martínez, S.C. Demographic history and spatial genetic structure in a remnant population of the subtropical tree Anadenanthera colubrina var. cebil (Griseb.) Altschul (Fabaceae). Ann. For. Sci. 2019, 76, 18. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Primers (5′-3′) | Ta (°C) | Repeat Motif | Size Range (bp) | Accession Number | A | Ho | He |
|---|---|---|---|---|---|---|---|---|
| Orot5 | F: GGTACTCGCGTTTGGGTCTA | 58 | (AC)10 | 196–223 | OP428738 | 2 | 0.55 | 0.58 |
| R: AGAGTAGTAGTCCCGGGTAAAA | ||||||||
| Orot8 | F: GTCGGAAACTCTACCAAAGTGA | 58 | (TC)8 | 131–140 | OP428739 | 4 | 0.62 | 0.63 |
| R: CCATCCCCGTAGAGTCTCG | ||||||||
| Orot11 | F: TGACAAAGGGTACGTATGAGC | 57 | (CTT)6 | 188–193 | OP428740 | 6 | 0.41 | 0.49 |
| R: TAATCCCTACCACATGCCCG | ||||||||
| Orot15 | F: TTTCTATACCTACGCGCCGG | 58 | (TTCA)10 | 179–187 | OP428741 | 3 | 0.45 | 0.49 |
| R: TAAACCTCCCTCTCCCCTCT | ||||||||
| Orot21 | F: CGGGACTATCAGAAGGTACGT | 59 | (GT)22 | 180–185 | OP428742 | 6 | 0.13 * | 0.21 |
| R: TGGGTAAAAGTCTGCTGATCCT | ||||||||
| Orot22 | F: TCCTCCTACTCCTATCTACGGA | 50 | (CT)13 | 148–155 | OP428743 | 6 | 0.18 * | 0.32 |
| R: ATCGTCTCTGCTATCCCTGC | ||||||||
| Orot32 | F: CCTCTACTATTCTCTTTAGCGCA | 55 | (GAA)8 | 170–175 | OP428744 | 8 | 0.44 | 0.45 |
| R: TGCCGATCTGACTATGGAGG | ||||||||
| Orot34 | F: GGGATCGATCGAAAGCTACG | 60 | (TA)22 | 190–195 | OP428745 | 5 | 0.56 | 0.58 |
| R: TCCTCCTAGTCCCGTAGTCC |
| Populations | Location | Latitude | Longitude | Altitude | N | A | Ae | Apr | Ho | He | F |
|---|---|---|---|---|---|---|---|---|---|---|---|
| LOJ | Loja | −3.91 | −79.15 | 2600 | 25 | 11.3 b | 8.9 b | 8 b | 0.72 b | 0.86 b | 0.22 b* |
| CER | Cerro Toledo | −4.38 | −79.13 | 2900 | 30 | 9.1 b | 8.4 b | 11 b | 0.73 b | 0.81 b | 0.21 b* |
| YAN | Yangana | −4.36 | −79.30 | 3000 | 30 | 10.4 b | 10.8 b | 9 b | 0.69 b | 0.73 b | 0.19 a* |
| CAT | Catamayo | −4.04 | −79.23 | 2750 | 25 | 12.3 b | 10.1 b | 9 b | 0.67 b | 0.80 b | 0.11 a* |
| ZAM | Zamora | −4.05 | −78.99 | 2150 | 30 | 6.1 a | 4.2 a | 2 a | 0.43 a | 0.62 a | 0.27 c* |
| Overall | 140 | 9.84 ± 2.40 | 7.06 ± 2.86 | 7.81 ± 3.42 | 0.65 ± 0.12 | 0.76 ± 0.09 | 0.20 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, I.; Draper, D.; Riofrío, L.; Naranjo, C. Early Signs of the Effects of Forest Fragmentation on the Genetic Diversity and Structure of the Threatened Ecuadorian Tree Ocotea rotundata (Lauraceae). Forests 2022, 13, 1940. https://doi.org/10.3390/f13111940
Marques I, Draper D, Riofrío L, Naranjo C. Early Signs of the Effects of Forest Fragmentation on the Genetic Diversity and Structure of the Threatened Ecuadorian Tree Ocotea rotundata (Lauraceae). Forests. 2022; 13(11):1940. https://doi.org/10.3390/f13111940
Chicago/Turabian StyleMarques, Isabel, David Draper, Lorena Riofrío, and Carlos Naranjo. 2022. "Early Signs of the Effects of Forest Fragmentation on the Genetic Diversity and Structure of the Threatened Ecuadorian Tree Ocotea rotundata (Lauraceae)" Forests 13, no. 11: 1940. https://doi.org/10.3390/f13111940