

Soil Organic Nitrogen Components and N−Cycling Enzyme Activities Following Vegetation Restoration of Cropland in Danxia Degraded Region
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Soil Sampling
2.3. Soil Analysis
2.4. Data Analysis
3. Results
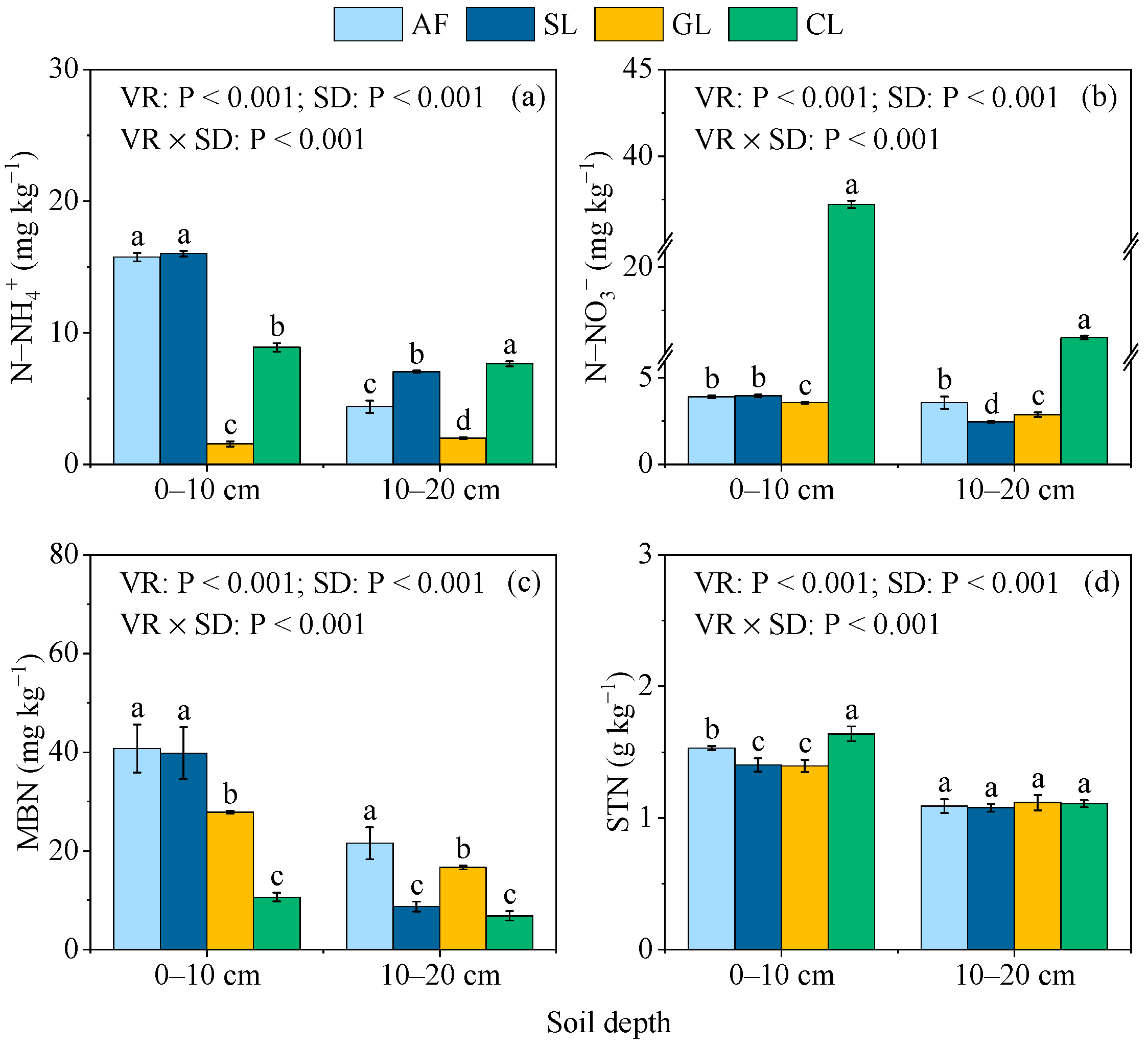
3.1. Soil Mineral N, MBN, and STN Contents
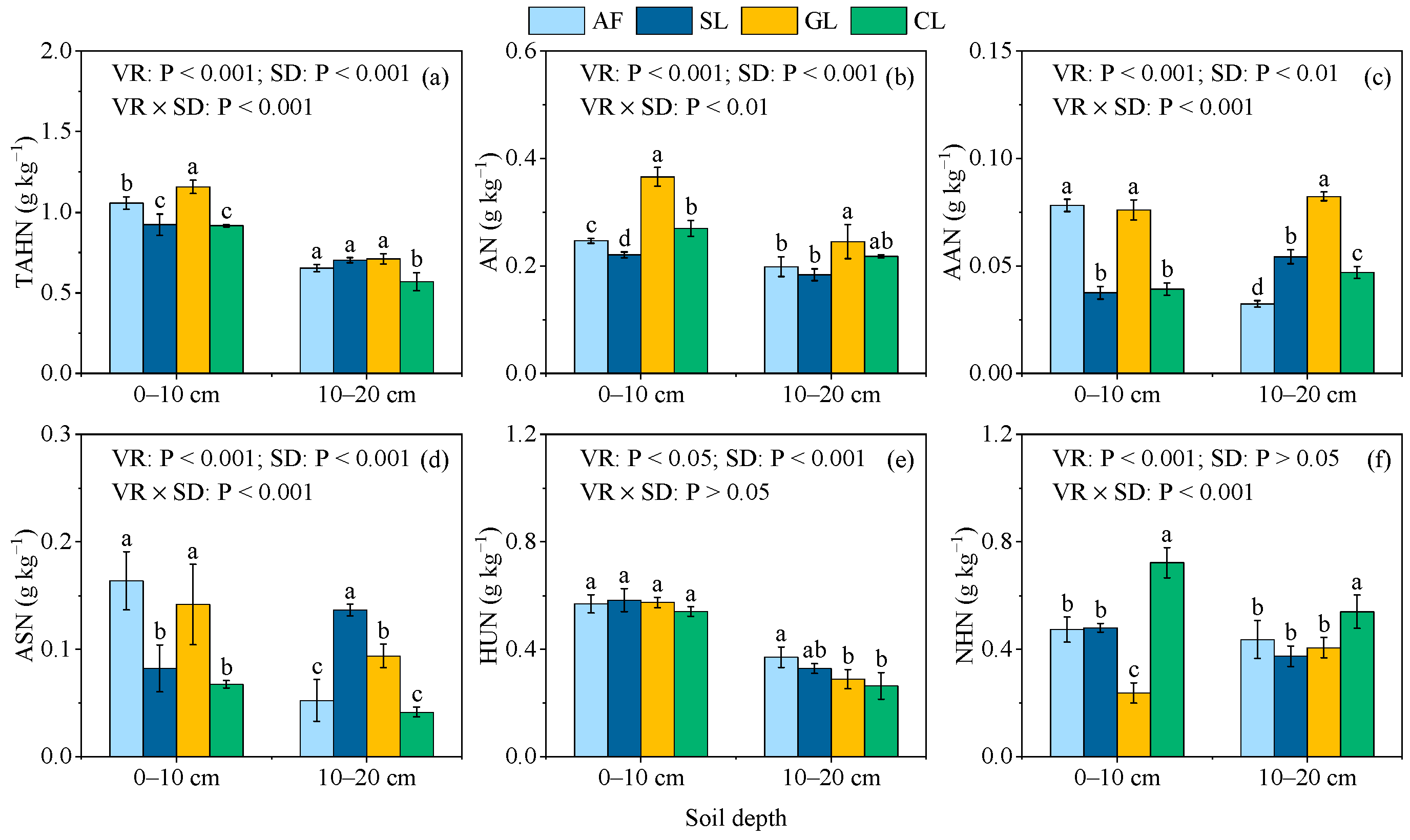
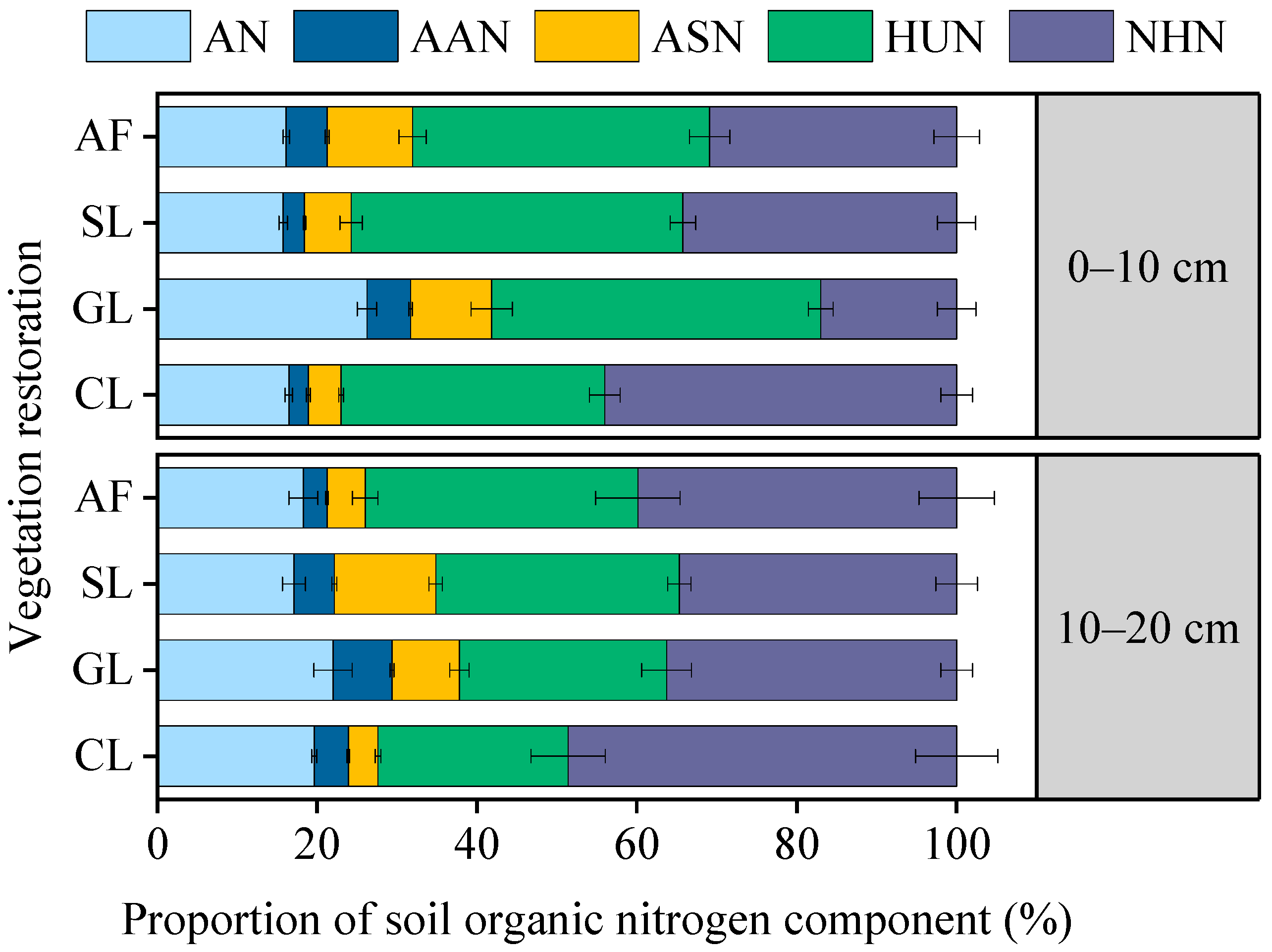
3.2. Contents and Proportion of SON Components
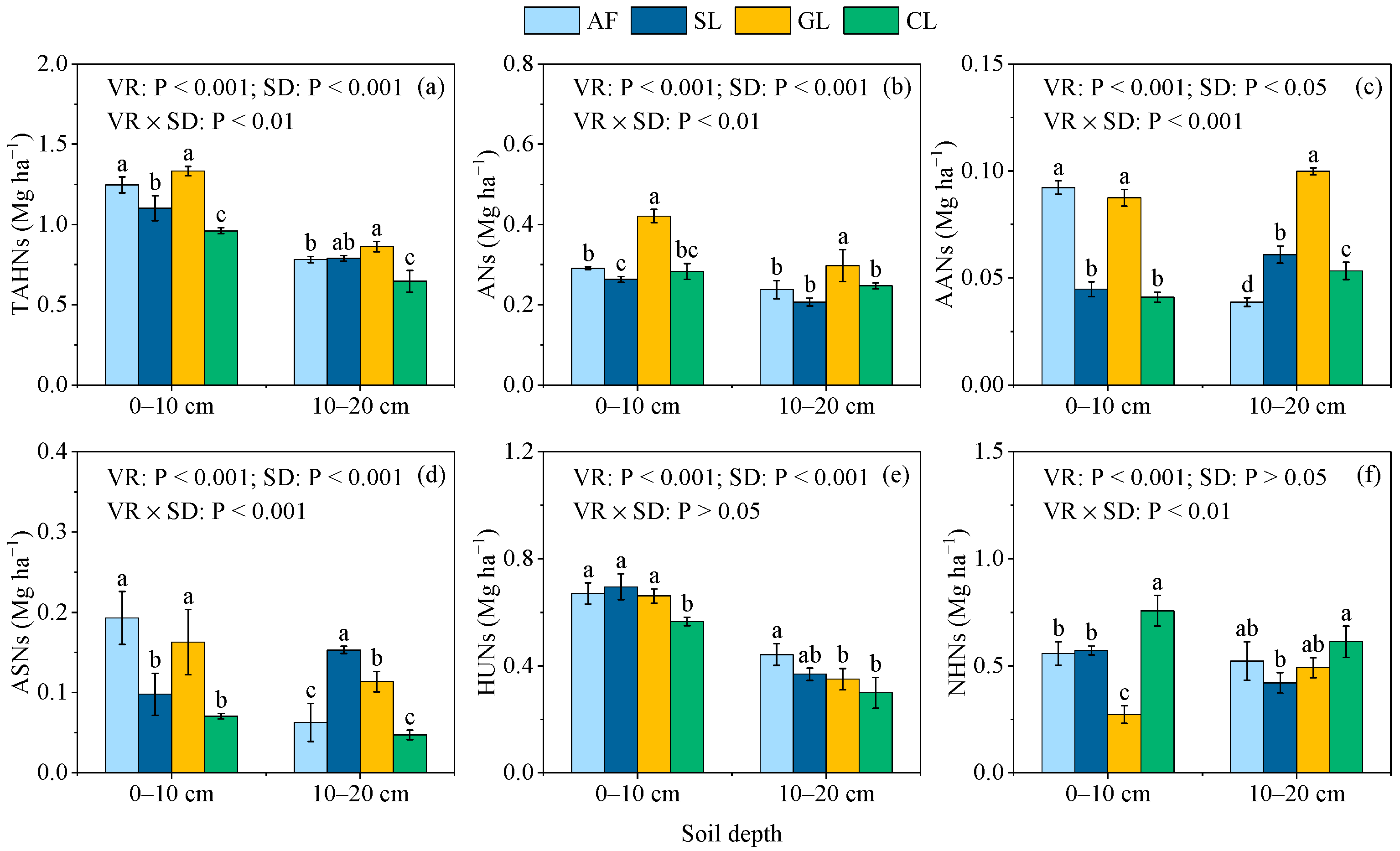
3.3. Stocks of SON Components
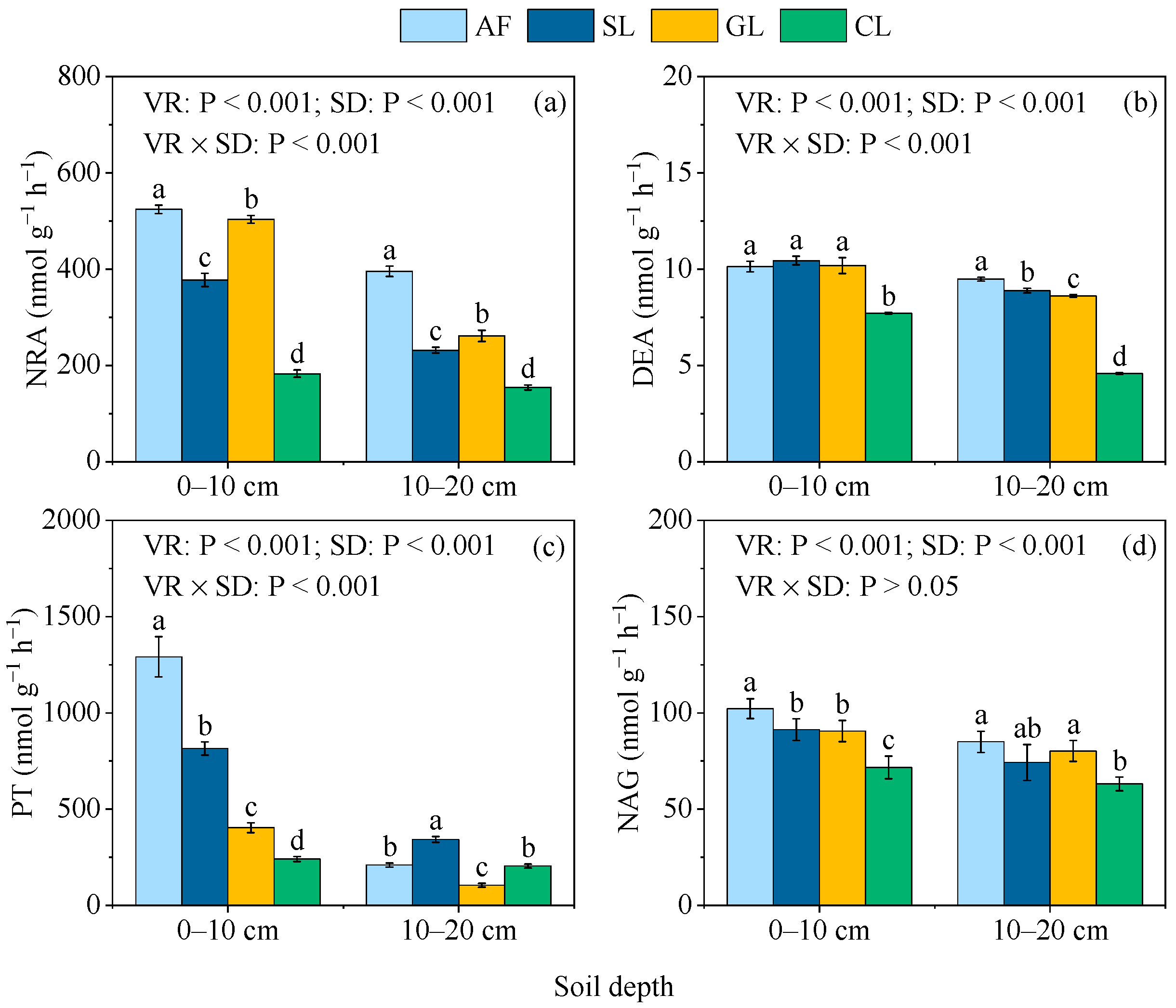
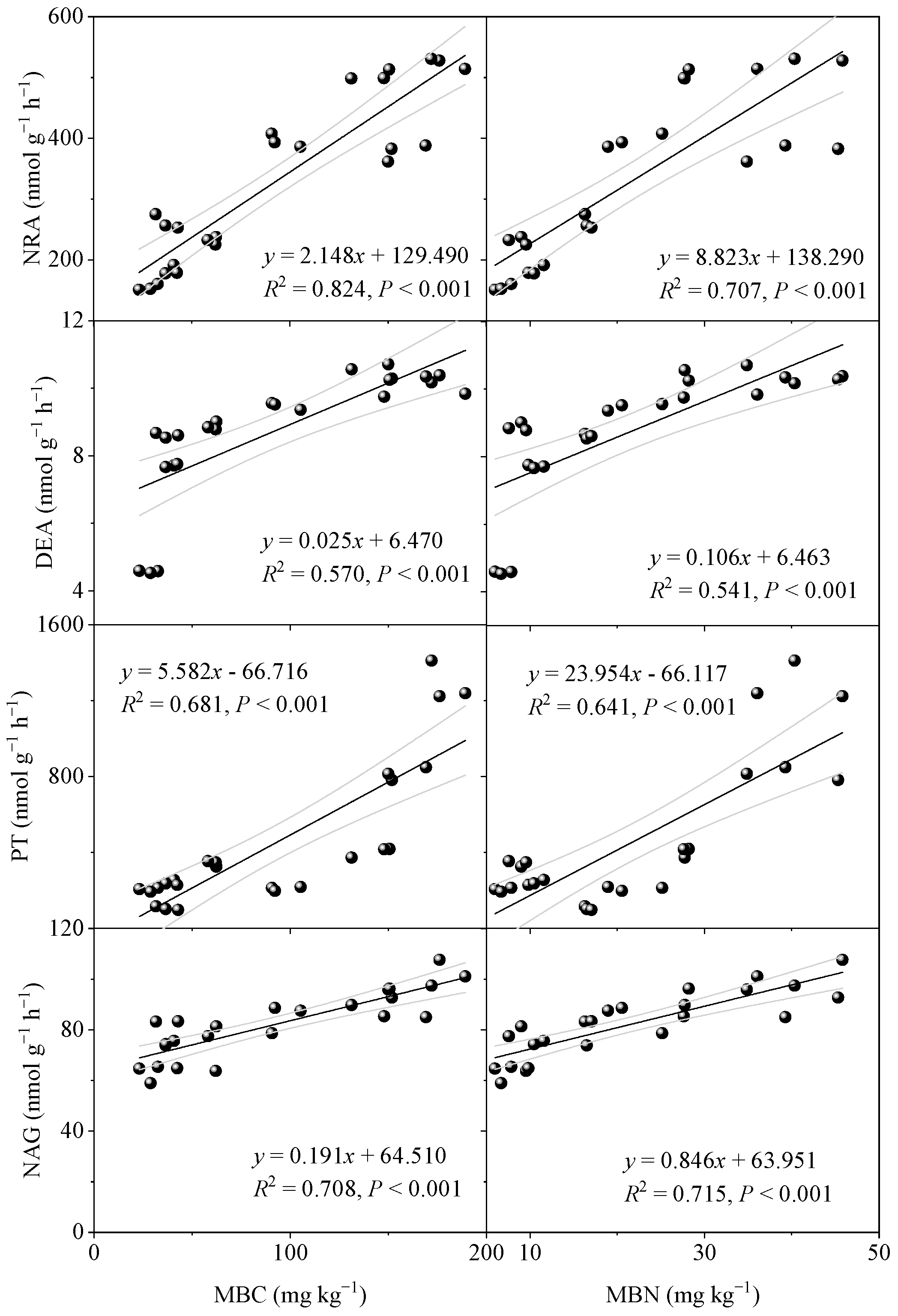
3.4. Soil-N-Cycling Enzyme Activities
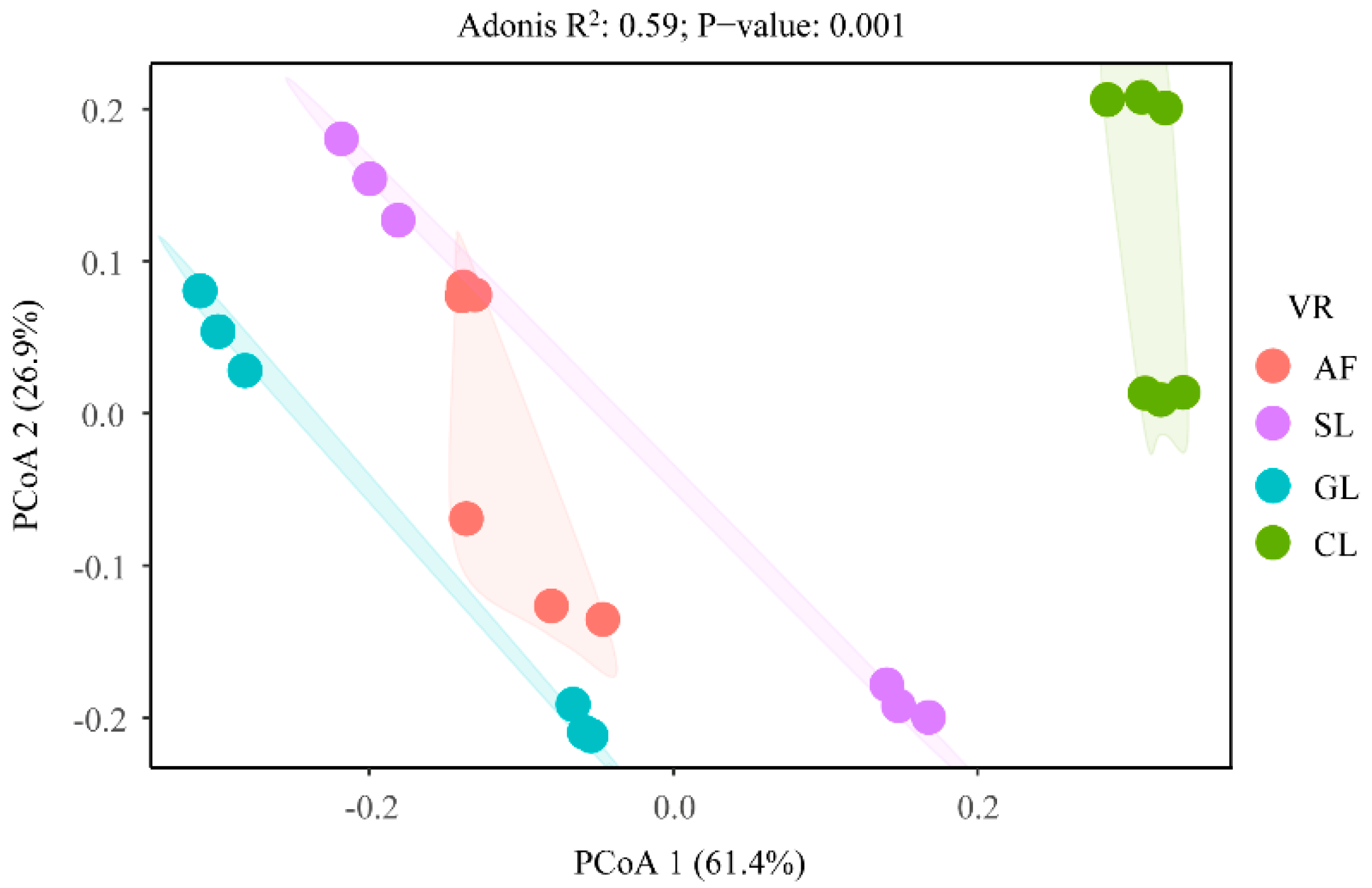
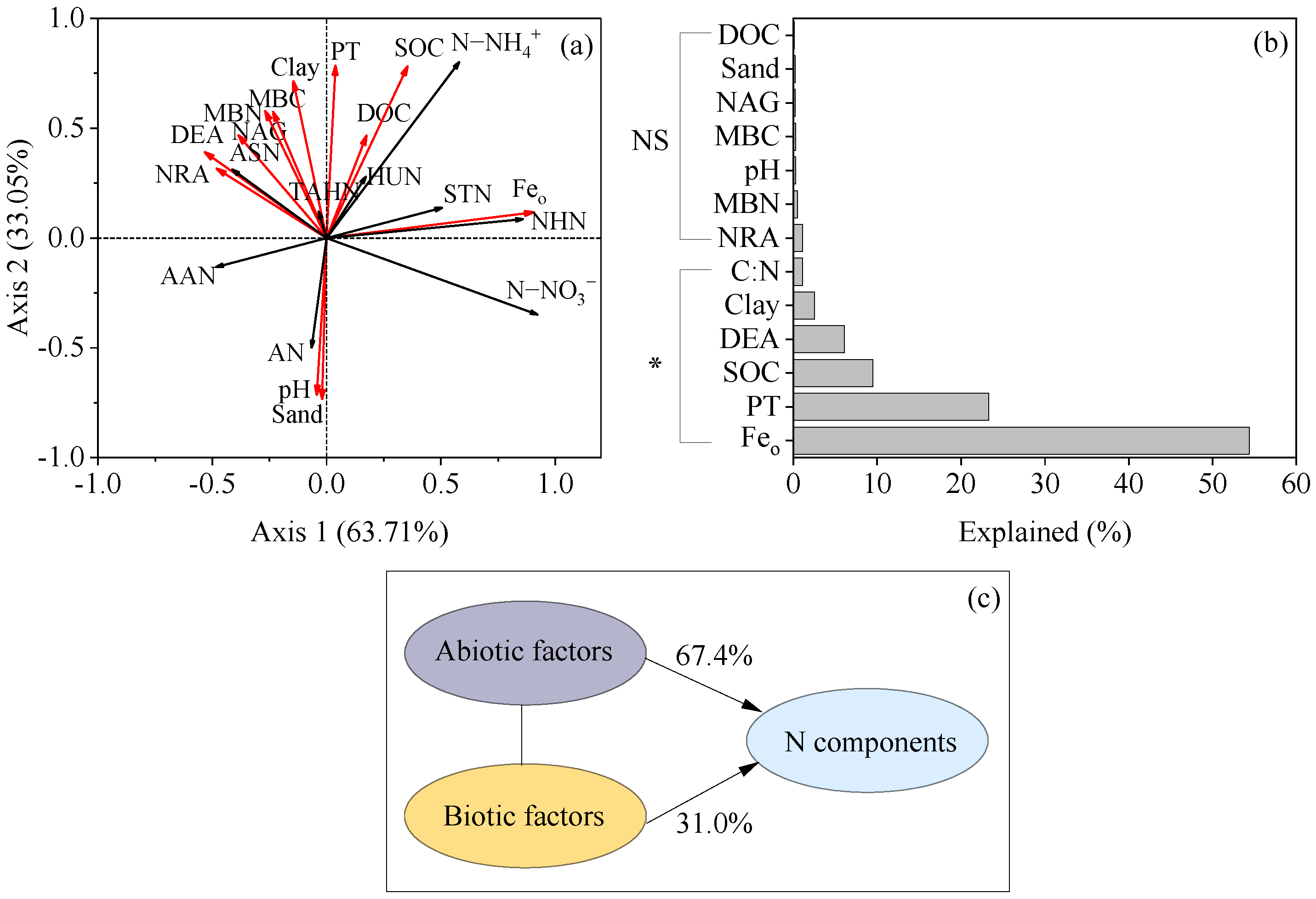
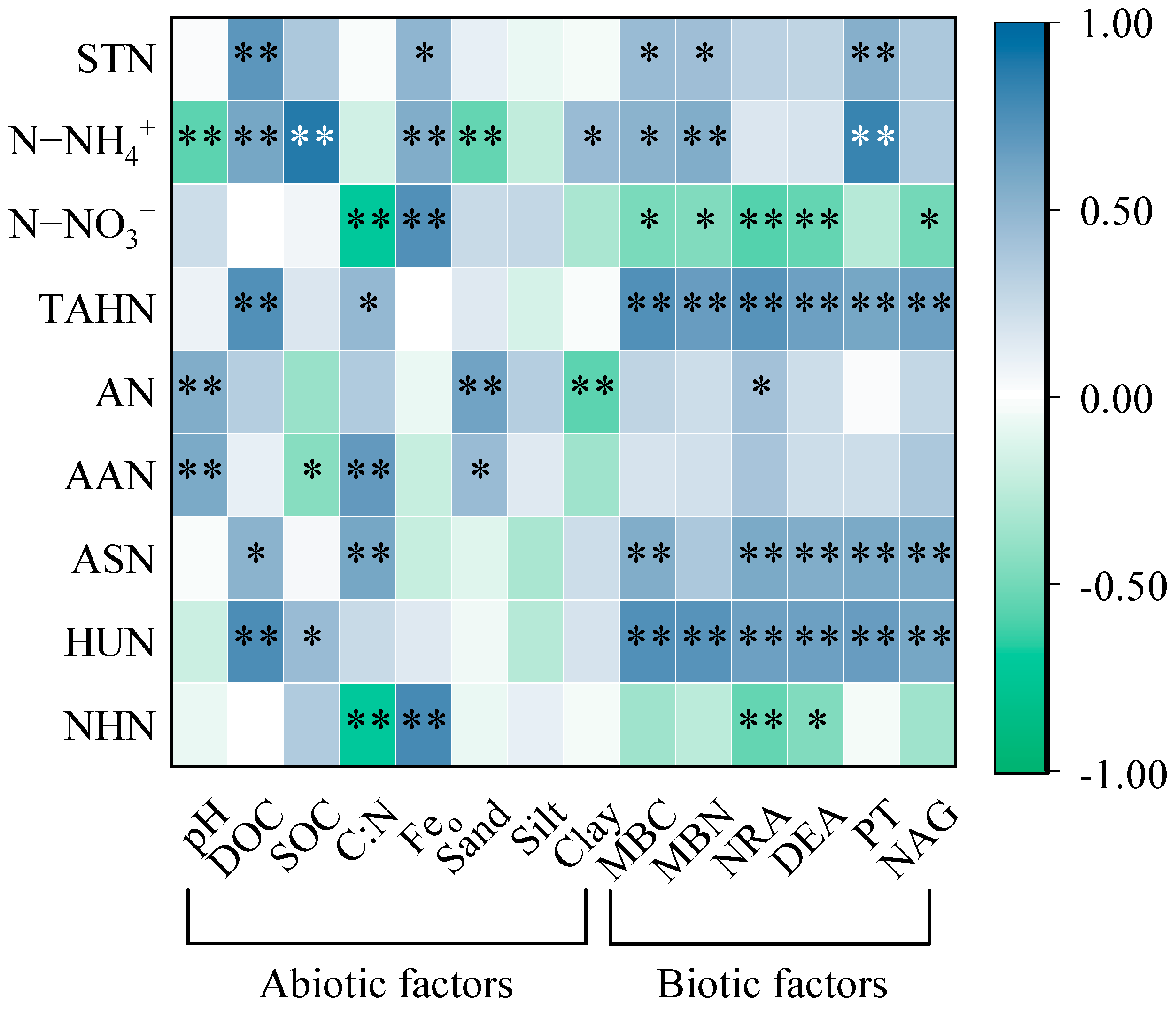
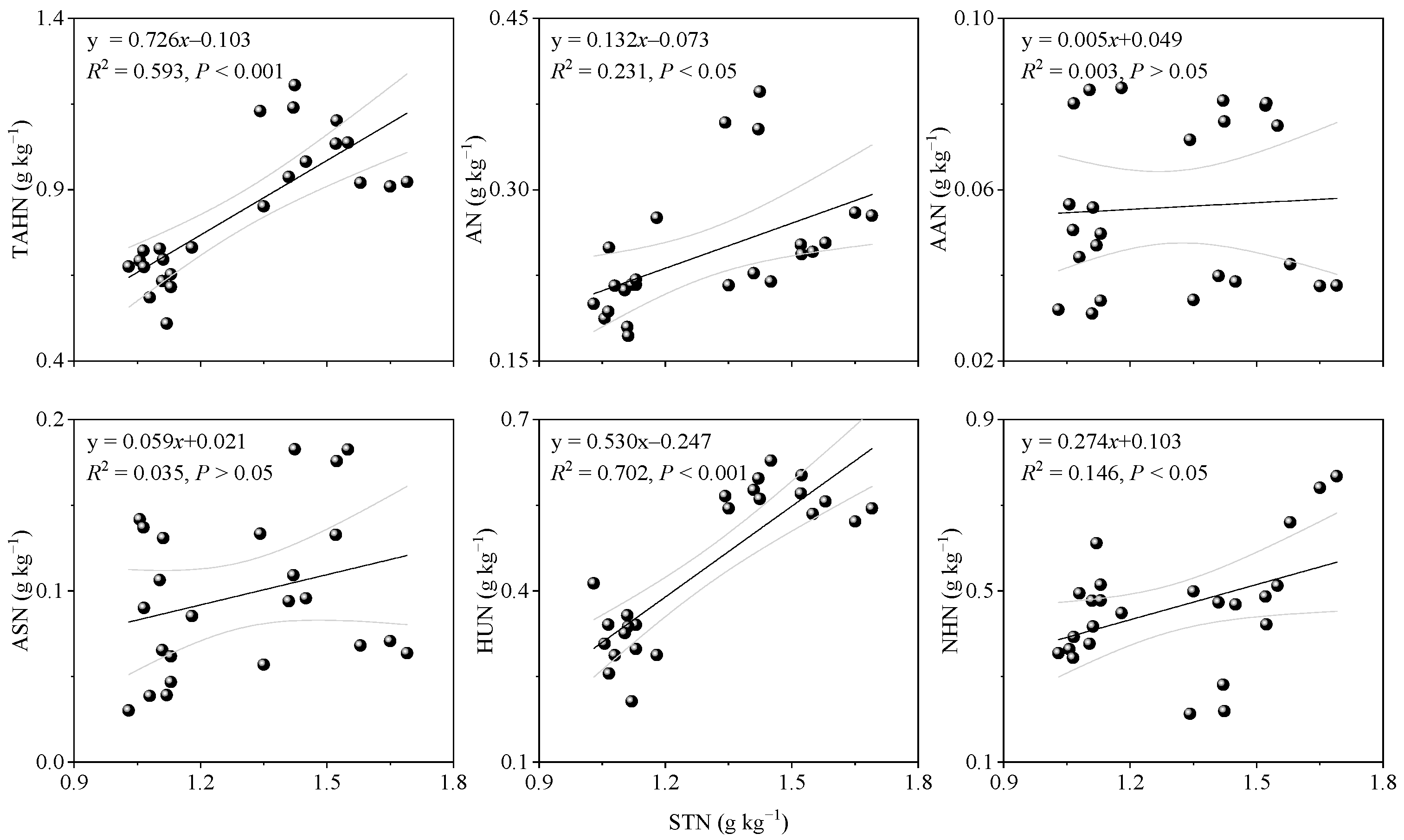
3.5. Factors That Affected the Transformation of Soil N Components
4. Discussion
4.1. Effect of Vegetation Restoration on SON Components
4.2. Effect of Vegetation Restoration on Soil Cycling Enzyme Activities
4.3. Factors That Influenced the Soil N Transformation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Feng, J.; Turner, B.L.; Wei, K.; Tian, J.H.; Chen, Z.H.; Lü, X.T.; Wang, C.; Chen, L.J. Divergent composition and turnover of soil organic nitrogen along a climate gradient in arid and semiarid grasslands. Geoderma 2018, 327, 36–44. [Google Scholar] [CrossRef]
- Xu, L.; He, N.P.; Yu, G.R. Nitrogen storage in China's terrestrial ecosystems. Sci. Total Environ. 2020, 709, 136201. [Google Scholar] [CrossRef] [PubMed]
- Li, D.J.; Yang, Y.; Chen, H.; Xiao, K.C.; Song, T.Q.; Wang, K.L. Soil gross nitrogen transformations in typical karst and non-karst forests, Southwest China. J. Geophys. Res. Biogeosci. 2017, 122, 2831–2840. [Google Scholar] [CrossRef]
- Wu, G.H.; Chen, Z.H.; Jiang, N.; Jiang, H.; Chen, L.J. Effects of long-term no-tillage with different residue application rates on soil nitrogen cycling. Soil Till. Res. 2021, 212, 105044. [Google Scholar] [CrossRef]
- Lin, T.C.; Shaner, P.J.L.; Wang, L.J.; Shih, Y.T.; Wang, C.P.; Huang, G.H.; Huang, J.C. Effects of mountain tea plantations on nutrient cycling at upstream watersheds. Hydrol. Earth Syst. Sci. 2015, 19, 4493–4504. [Google Scholar] [CrossRef] [Green Version]
- Lozano, Y.M.; Hortal, S.; Armas, C.; Pugnaire, F.I. Interactions among soil, plants, and microorganisms drive secondary succession in a dry environment. Soil Biol. Biochem. 2014, 78, 298–306. [Google Scholar] [CrossRef]
- Ma, R.T.; Hu, F.N.; Xu, C.Y.; Liu, J.F.; Zhao, S.W. Response of soil aggregate stability and splash erosion to different breakdown mechanisms along natural vegetation restoration. Catena 2022, 208, 105775. [Google Scholar] [CrossRef]
- Lan, J.C.; Hu, N.; Fu, W.L. Soil carbon-nitrogen coupled accumulation following the natural vegetation restoration of abandoned farmlands in a karst rocky desertification region. Ecol. Eng. 2020, 158, 106033. [Google Scholar] [CrossRef]
- Hu, P.L.; Zhao, Y.; Xiao, D.; Xu, Z.H.; Zhang, W.; Xiao, J.; Wang, K.L. Dynamics of soil nitrogen availability following vege-tation restoration along a climatic gradient of a subtropical karst region in China. J. Soils Sediments 2021, 21, 2167–2178. [Google Scholar] [CrossRef]
- Zhang, D.; Cai, X.W.; Diao, L.F.; Wang, Y.Q.; Wang, J.S.; An, S.Q.; Cheng, X.L.; Yang, W. Changes in soil organic carbon and nitrogen pool sizes, dynamics, and biochemical stability during ~160 years natural vegetation restoration on the Loess Plateau, China. Catena 2022, 211, 106014. [Google Scholar] [CrossRef]
- Stevenson, F.J. Organic forms of soil nitrogen. In Nitrogen in Agricultural Soils; American Society of Agronomy: Madison, WI, USA, 1982; Volume 22, pp. 1–42. [Google Scholar]
- Amelung, W.; Zhang, X. Determination of amino acid enantiomers in soils. Soil Biol. Biochem. 2001, 33, 553–562. [Google Scholar] [CrossRef]
- Lü, H.J.; He, H.B.; Zhao, J.S.; Wang, W.; Xie, H.T.; Hu, G.Q.; Liu, X.; Wu, Y.Y.; Zhang, X.D. Dynamics of fertilizer derived organic nitrogen fractions in an arable soil during a growing season. Plant Soil 2013, 373, 595–607. [Google Scholar] [CrossRef]
- Zaman, T.; Iqbal, A.; Shaukat, A.; Nazir, R.; Pervez, A.; Bilal, M.; Faridullah, M.R.; Ali, S.; Alkahtani, S.; Abdel-Daim, M.M.; et al. Assessing the N cycling ecosystem function-processes and the involved functional guilds upon plant litter amendment in lower Himalaya. Pol. J. Environ. Stud. 2021, 30, 917–926. [Google Scholar] [CrossRef]
- Luxhøi, J.; Elsgarrd, L.; Thomsen, I.K.; Jensen, L.S. Effects of long-term annual inputs of straw and organic manure on plant N uptake and soil N fluxes. Soil Use Manag. 2007, 23, 368–373. [Google Scholar] [CrossRef]
- Zhang, M.; Dong, L.G.; Wang, Y.; Bai, X.X.; Ma, Z.T.; Yu, X.; Zhao, Z. The response of soil microbial communities to soil erodibility depends on the plant and soil properties in semiarid regions. Land Degrad. Dev. 2021, 32, 3180–3193. [Google Scholar] [CrossRef]
- Fujii, K.; Yamada, T.; Hayakawa, C.; Nakanishi, A.; Funakawa, S. Decoupling of protein depolymerization and ammonification in nitrogen mineralization of acidic forest soils. Appl. Soil Ecol. 2020, 153, 103572. [Google Scholar] [CrossRef]
- Tian, J.H.; Wei, K.; Sun, T.; Jiang, N.; Chen, Z.H.; Feng, J.; Cai, K.Z.; Chen, L.J. Different forms of nitrogen deposition show variable effects on soil organic nitrogen turnover in a temperate forest. Appl. Soil Ecol. 2022, 169, 104212. [Google Scholar] [CrossRef]
- Guan, H.L.; Fan, J.W.; Lu, X.K. Soil specific enzyme stoichiometry reflects nitrogen limitation of microorganisms under different types of vegetation restoration in the karst areas. Appl. Soil Ecol. 2021, 169, 104253. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, C.; Wang, Y.Q.; Cheng, H.; An, S.S.; Chang, S.X. Soil extracellular enzyme stoichiometry reflects the shift from P- to N-limitation of microorganisms with grassland restoration. Soil Biol. Biochem. 2020, 149, 107928. [Google Scholar] [CrossRef]
- Li, X.J.; Yang, H.T.; Shi, W.L.; Li, Y.F.; Guo, Q. Afforestation with xerophytic shrubs accelerates soil net nitrogen nitrification and mineralization in the Tengger Desert, Northern China. Catena 2018, 169, 11–20. [Google Scholar] [CrossRef]
- Pastore, G.; Tobin, B.; Nieuwenhuis, M. Quantifying carbon and nitrogen losses by respiration and leaching from decomposing woody debris in reforested coniferous stands in Ireland. Agric. For. Meteorol. 2019, 265, 195–207. [Google Scholar] [CrossRef]
- Ding, X.Q.; Chang, Y.; Hou, H.B.; Peng, P.Q.; Xiang, W.H. Quantification of the sources of soluble organic N (SON) from new litter or indigenous soil in a typical subtropical forest. Land Degrad. Dev. 2021, 32, 2528–2539. [Google Scholar] [CrossRef]
- Yan, L.B.; Peng, H.; Zhang, S.; Zhang, R.X.; Kasanin-Grubin, M.; Lin, K.R.; Tu, X.J. The spatial patterns of red beds and Danxia landforms: Implication for the formation factors-China. Sci. Rep. 2019, 9, 1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.L.; Liao, W.B.; Li, Z.; Jia, F.L.; Wang, Y.Y.; Chang, H.; Zeng, S.C.; Jin, J.H.; Xin, G.R.; Chen, B.M.; et al. Integrated Biological Surveys on Mount Danxia, Guangdong; Sciences Press: Beijing, China, 2011. [Google Scholar]
- Yan, L.B.; Kasanin-Grubin, M. Land degradation and management of red beds in China: Two case studies. J. Mt. Sci. 2019, 16, 2591–2604. [Google Scholar] [CrossRef]
- FAO-UNESCO. FAO-UNESCO Soil Map of the World; UNESCO: Paris, France, 1974. [Google Scholar]
- Lu, R.K. Methods for Soil Agrochemical Analysis; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Guan, S.Y. Soil Enzyme and Research Method; China Agricultural Press: Beijing, China, 1986. [Google Scholar]
- Welsh, M.K.; Mcmillan, S.K.; Vidon, P.G. Denitrification along the stream-riparian continuum in restored and unrestored agricultural streams. J. Environ. Qual. 2017, 46, 1010–1019. [Google Scholar] [CrossRef]
- Liu, Y.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. Root exudates shift how N mineralization and N fixation contribute to the plant-available N supply in low fertility soils. Soil Biol. Biochem. 2022, 165, 108541. [Google Scholar] [CrossRef]
- Khalil, M.I.; Hossain, M.B.; Schmidhalter, U. Carbon and nitrogen mineralization in different upland soils of the subtropics treated with organic material. Soil Biol. Biochem. 2005, 37, 1507–1518. [Google Scholar] [CrossRef]
- Cheng, Y.; Cai, Z.C.; Chang, S.X.; Wang, J.; Zhang, J.B. Wheat straw and its biochar have contrasting effects on inorganic N retention and N2O production in a cultivated Black Chernozem. Biol. Fert. Soils 2012, 48, 941–946. [Google Scholar] [CrossRef]
- Zheng, X.Z.; Lin, C.; Guo, B.L.; Yu, H.H.; Ding, H.; Peng, S.Y.; Zhang, J.B.; Ireland, E.; Chen, D.L.; Mvller, C.; et al. Mechanisms behind soil N dynamics following cover restoration in degraded land in subtropical China. J. Soil Sediments 2020, 20, 1897–1905. [Google Scholar] [CrossRef]
- Berg, M.P.; Verhoef, H.A.; Bolger, T.; Mccarthy, F.; Anderson, J.M.; Splatt, P.; Willison, T. Effects of air pollutant temperature interactions on mineral N dynamics and cation leaching in reciplicate forest soil transplantation experiments. Biogeochemistry 1997, 39, 295–326. [Google Scholar] [CrossRef]
- Li, S.S.; Tong, Y.W.; Wang, Z.W. Species and genetic diversity affect leaf litter decomposition in subtropical forest in southern China. J. Plant Ecol. 2017, 10, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Qiu, S.J.; Peng, P.Q.; Li, L.; He, P.; Liu, Q.; Wu, J.S.; Christie, P.; Ju, X.T. Effects of applied urea and straw on various nitrogen fractions in two Chinese paddy soils with differing clay mineralogy. Biol. Fert. Soils 2012, 48, 161–172. [Google Scholar] [CrossRef]
- Li, L.; Wilson, C.B.; He, H.; Zhang, X.; Zhou, F.; Schaeffer, S.M. Physical, biochemical, and microbial controls on amino sugar accumulation in soils under long- term cover cropping and no-tillage farming. term cover cropping and no-tillage farming. Soil Biol. Biochem. 2019, 135, 369–378. [Google Scholar] [CrossRef]
- Wagai, R.; Mayer, L.M.; Kitayama, K.; Shirato, Y. Association of organic matter with iron and aluminum across a range of soils determined via selective dissolution techniques coupled with dissolved nitrogen analysis. Biogeochemistry 2013, 112, 95–109. [Google Scholar] [CrossRef]
- Guan, X.K.; Wei, L.; Turner, N.C.; Ma, S.C.; Yang, M.D.; Wang, T.C. Improved straw management practices promote in situ straw decomposition and nutrient release, and increase crop production. J. Clean. Prod. 2020, 250, 119514. [Google Scholar] [CrossRef]
- Osborne, B.B.; Soper, F.M.; Nasto, M.K.; Bru, D.; Hwang, S.; Machmuller, M.B.; Morales, M.L.; Philippot, L.; Sullivan, B.W.; Asner, G.P. Litter inputs drive patterns of soil nitrogen heterogeneity in a diverse tropical forest: Results from a litter manip-ulation experiment. Soil Biol. Biochem. 2021, 158, 108247. [Google Scholar] [CrossRef]
- Barton, L.; Schipper, L.A.; Smith, C.T.; Mcaly, C.D.A. Denitrification enzyme activity is limited by soil aeration in a wastewater-irrigated forest soil. Biol. Fert. Soils. 2000, 32, 385–389. [Google Scholar] [CrossRef]
- Judy, S.; Michael, D.; Rodica, P.; Arthur, G.; Heinz, R. Nitrogen nutrition of beech forests in a changing climate: Importance of plant-soil-microbe water carbon, and nitrogen interactions. Plant Soil 2017, 418, 89–114. [Google Scholar]
- Han, H.; Chen, C.; Bai, M.H.; Xu, T.; Yang, H.F.; Shi, A.M.; Ding, G.C.; Li, J. Abundance and diversity of denitrifying bacterial communities associated with N2O emission under long-term organic farming. Eur. J. Soil Biol. 2020, 97, 103153. [Google Scholar] [CrossRef]
- Šimek, M.J.; Jı́šová, L.; Hopkins, D.W. What is the so-called optimum pH for denitrification in soil. Soil Biol. Biochem. 2002, 34, 1227–1234. [Google Scholar] [CrossRef]
- Fu, G.P.; Wu, J.F.; Han, J.Y.; Zhao, L.; Chan, G.; Leong, K.F. Effects of substrate type on denitrification efficiency and microbial community structure in constructed wetlands. Bioresour. Technol. 2020, 307, 123222. [Google Scholar] [CrossRef] [PubMed]
- Kappler, A.; Bryce, C.; Mansor, M.; Lueder, U.; Byrne, J.M.; Swanner, E.D. An evolving view on biogeochemical cycling of iron. Nat. Rev. Microbiol. 2021, 19, 360–374. [Google Scholar] [CrossRef] [PubMed]
- Krause, L.; Klumpp, E.; Nofz, I.; Missong, A.; Amelung, W.; Siebers, N. Colloidal iron and organic carbon control soil aggregate formation and stability in arable Luvisols. Geoderma 2020, 374, 114421. [Google Scholar] [CrossRef]
- Chen, C.M.; Hall, S.J.; Coward, E.; Thompson, A. Iron-mediated organic matter decomposition in humid soils can counteract protection. Nat. Commun. 2020, 11, 2255. [Google Scholar] [CrossRef]
- Chen, C.M.; James, J.D.; Wang, J.; Sparks, D.L. Properties of Fe-organic matter associations via coprecipitation versus ad-sorption. Environ. Sci. Technol. 2014, 48, 13751–13759. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Hammerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of microbial nitrogen use efficiency to carbon: Nitrogen imbalances regulates soil nitrogen cycling. Nat. Commun. 2014, 5, 3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerard, H.R. Predicting soil N mineralization using organic matter fractions and soil properties: A re-analysis of literature data. Soil Biol. Biochem. 2012, 45, 132–135. [Google Scholar]
- Farrell, M.; Prendergast-Miller, M.; Jones, D.L.; Hill, P.W.; Condron, L.M. Soil microbial organic nitrogen uptake is regulated by carbon availability. Soil Biol. Biochem. 2014, 77, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Shahid, M.; Nayak, A.K.; Puree, C.; Tripathi, R.; Lal, B.; Gautam, P.; Bhattacharyya, P.; Mohanty, S.; Kumar, A.; Panda, B.B.; et al. Carbon and nitrogen fractions and stocks under 41 years of chemical and organic fertilization in a sub-humid tropical rice soil. Soil Tillage Res. 2017, 170, 136–146. [Google Scholar] [CrossRef]
- Jin, H.M.; Sun, O.J.; Liu, J.F. Changes in soil microbial biomass and community structure with addition of contrasting types of plant litter in a semiarid grassland ecosystem. J. Plant Ecol. 2010, 3, 209–217. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Depth (cm) | Vegetation Restoration | pH | BD (g cm−3) | DOC (mg kg−1) | MBC (mg kg−1) | SOC (g kg−1) | C:N | Feo (g kg−1) | Sand (%) | Silt (%) | Clay (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0–10 | AF | 5.21 ± 0.04 c | 1.18 ± 0.01 c | 291.10 ± 5.95 a | 179.16 ± 8.97 a | 14.13 ± 0.81 b | 17.36 ± 1.22 a | 2.42 ± 0.13 b | 20.21 ± 0.94 c | 46.71 ± 2.08 b | 33.08 ± 2.34 a |
| SL | 5.16 ± 0.17 c | 1.19 ± 0.00 b | 274.09 ± 12.81 ab | 157.10 ± 10.58 b | 24.32 ± 0.84 a | 7.32 ± 0.26 b | 1.72 ± 0.07 c | 23.16 ± 1.58 c | 45.06 ± 1.23 b | 31.79 ± 2.19 a | |
| GL | 6.28 ± 0.13 a | 1.15 ± 0.03 a | 275.82 ± 7.65 ab | 143.36 ± 10.44 b | 4.56 ± 0.39 c | 16.35 ± 0.95 a | 1.05 ± 0.06 d | 37.33 ± 4.47 a | 54.57 ± 4.85 a | 8.09 ± 1.13 c | |
| CL | 6.01 ± 0.12 b | 1.05 ± 0.02 a | 260.20 ± 14.75 b | 40.00 ± 3.04 c | 13.55 ± 0.56 b | 3.13 ± 0.34 c | 2.84 ± 0.07 a | 32.29 ± 1.30 b | 49.15 ± 1.19 b | 18.56 ± 0.14 b | |
| 10–20 | AF | 4.80 ± 0.03 d | 1.20 ± 0.01 b | 230.69 ± 6.15 bc | 96.19 ± 8.09 a | 7.41 ± 0.56 b | 13.16 ± 1.44 a | 0.99 ± 0.07 d | 17.91 ± 1.22 c | 43.89 ± 1.51 c | 38.20 ± 0.29 b |
| SL | 5.29 ± 0.02 c | 1.12 ± 0.01 b | 254.25 ± 15.27 a | 60.92 ± 2.37 b | 14.29 ± 0.84 a | 8.47 ± 0.33 b | 1.30 ± 0.09 b | 17.53 ± 0.19 c | 41.63 ± 0.52 c | 40.84 ± 0.35 a | |
| GL | 6.77 ± 0.15 a | 1.21 ± 0.01 a | 215.40 ± 8.25 c | 37.13 ± 5.65 c | 4.83 ± 0.44 c | 12.90 ± 1.39 a | 1.12 ± 0.02 c | 41.00 ± 1.76 a | 50.57 ± 1.77 b | 8.43 ± 0.42 c | |
| CL | 5.91 ± 0.06 b | 1.13 ± 0.02 a | 238.22 ± 11.04 ab | 28.31 ± 4.82 c | 8.41 ± 0.44 b | 3.94 ± 0.17 c | 2.81 ± 0.06 a | 30.00 ± 1.22 b | 66.19 ± 0.58 a | 3.81 ± 0.65 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Yang, Q.; Zhang, C.; Zhou, B.; Li, X.; Zhang, X.; Chen, J.; Liu, K. Soil Organic Nitrogen Components and N−Cycling Enzyme Activities Following Vegetation Restoration of Cropland in Danxia Degraded Region. Forests 2022, 13, 1917. https://doi.org/10.3390/f13111917
Wang C, Yang Q, Zhang C, Zhou B, Li X, Zhang X, Chen J, Liu K. Soil Organic Nitrogen Components and N−Cycling Enzyme Activities Following Vegetation Restoration of Cropland in Danxia Degraded Region. Forests. 2022; 13(11):1917. https://doi.org/10.3390/f13111917
Chicago/Turabian StyleWang, Chao, Qiannan Yang, Chi Zhang, Bo Zhou, Xiangdong Li, Xiaolong Zhang, Jing Chen, and Kexue Liu. 2022. "Soil Organic Nitrogen Components and N−Cycling Enzyme Activities Following Vegetation Restoration of Cropland in Danxia Degraded Region" Forests 13, no. 11: 1917. https://doi.org/10.3390/f13111917