
Mycorrhizal Benefits of Salt-Stressed Cinnamomum camphora (L.) Presl. May Be Related to P and Mn2+ Contents in Shoots, Biomass Allocation, and K+/Na+ in Roots and Shoots

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and AMF Preparation
2.2. Experimental Design and Treatments
2.3. Measurements
2.4. Data Analysis
3. Results
3.1. Mycorrhizal Colonization and Plant Growth
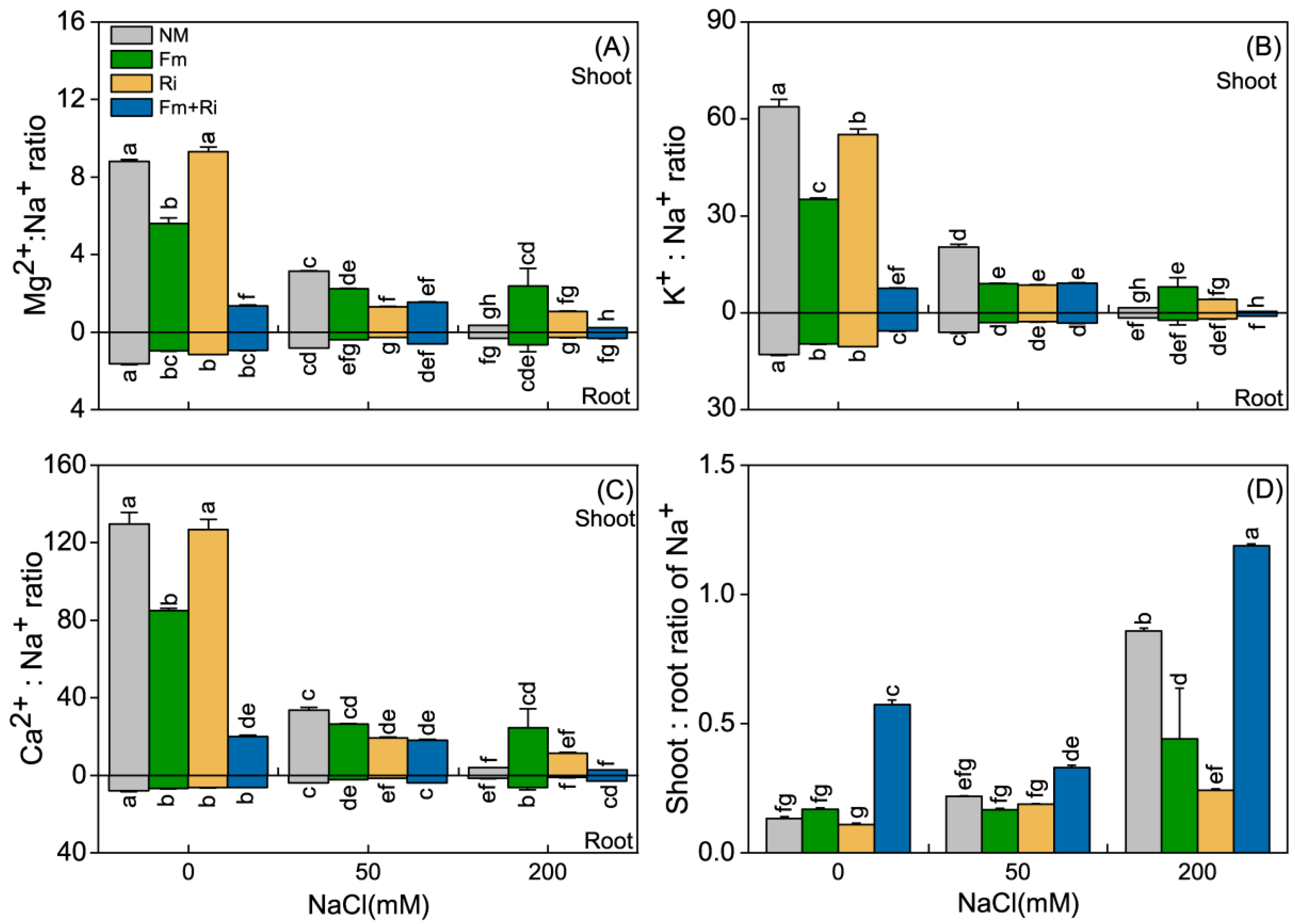
3.2. Nutrient Uptake and Allocation
3.3. Mycorrhizal Dependence and Selective Transporting Capacity
3.4. Mycorrhizal Dependence and Their Relationships with Other Plant Variables
4. Discussion
4.1. Improved Biomass with Mycorrhizal Inoculation
4.2. Adjusting Na+ Uptake and Translocation with Mycorrhizal Inoculation
4.3. Enhanced Nutrient Concentratioin and Ionic Homeostasis with Mycorrhizal Inoculation
4.4. Underlying Mechanism for Mycorrhizal Dependence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMF | arbuscular mycorrhizal fungi |
| AM | arbuscular mycorrhizal |
| AM plants | mycorrhizal plants |
| NM plants | non-mycorrhizal plants |
| EC | Electrical conductivity |
| MD | mycorrhizal dependence |
| STC | selective transporting capacity of K+ over Na+ from roots to shoots |
| LSD | Least significant difference |
References
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi: A review. Agron. Sustain. Dev. 2011, 32, 181–200. [Google Scholar] [CrossRef] [Green Version]
- Amanifar, S.; Toghranegar, Z. The efficiency of arbuscular mycorrhiza for improving tolerance of Valeriana officinalis L. and enhancing valerenic acid accumulation under salinity stress. Ind. Crops Prod. 2020, 147, 112234. [Google Scholar] [CrossRef]
- Dashtebani, F.; Hajiboland, R.; Aliasgharzad, N. Characterization of salt-tolerance mechanisms in mycorrhizal (Claroideoglomus etunicatum) halophytic grass, Puccinellia distans. Acta Physiol. Plant. 2014, 36, 1713–1726. [Google Scholar] [CrossRef]
- Pereira, P.; Barcelo, D.; Panagos, P. Soil and water threats in a changing environment. Environ. Res. 2020, 186, 109501. [Google Scholar] [CrossRef] [PubMed]
- Ondrasek, G.; Rengel, Z. Environmental salinization processes: Detection, implications & solutions. Sci. Total Environ. 2021, 754, 142432. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.M. Sustainability associated coastal eco-environmental problems and coastal science development in China. Bull. Chin. Acad. Sci. 2016, 31, 1133–1142. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Boughattas, S.; Hu, S.; Oh, S.H.; Sa, T. A meta-analysis of arbuscular mycorrhizal effects on plants grown under salt stress. Mycorrhiza 2014, 24, 611–625. [Google Scholar] [CrossRef]
- Dastogeer, K.M.G.; Zahan, M.I.; Tahjib-Ul-Arif, M.; Akter, M.A.; Okazaki, S. Plant salinity tolerance conferred by arbuscular mycorrhizal fungi and associated mechanisms: A meta-analysis. Front. Plant Sci. 2020, 11, 588550. [Google Scholar] [CrossRef]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of salinity stress in plants by arbuscular mycorrhizal symbiosis: Current understanding and new challenges. Front. Plant Sci. 2019, 10, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, UK, 2008; ISBN 978-0-12-370526-6. [Google Scholar]
- Aliasgharzad, N.; Saleh, R.N.; Towfighi, H.; Alizadeh, A. Occurrence of arbuscular mycorrhizal fungi in saline soils of the Tabriz Plain of Iran in relation to some physical and chemical properties of soil. Mycorrhiza 2001, 11, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Juniper, S.; Abbott, L. Vesicular-arbuscualr mycorrhizas and soil salinity. Mycorrhiza 1993, 4, 45–57. [Google Scholar] [CrossRef]
- Porras-Soriano, A.; Soriano-Martin, M.L.; Porras-Piedra, A.; Azcon, R. Arbuscular mycorrhizal fungi increased growth, nutrient uptake and tolerance to salinity in olive trees under nursery conditions. J. Plant Physiol. 2009, 166, 1350–1359. [Google Scholar] [CrossRef]
- Wang, J.P.; Fu, Z.Y.; Ren, Q.; Zhu, L.J.; Lin, J.; Zhang, J.C.; Cheng, X.F.; Ma, J.Y.; Yue, J.M. Effects of arbuscular mycorrhizal fungi on growth, photosynthesis, and nutrient uptake of Zelkova serrata (Thunb.) Makino seedlings under salt stress. Forests 2019, 10, 186. [Google Scholar] [CrossRef] [Green Version]
- Johnson, N.C.; Graham, J.H.; Smith, F.A. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytol. 1997, 135, 575–585. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Hart, M.M.; Reader, R.J. Taxonomic basis for variation in the colonization strategy of arbuscular mycorrhizal fungi. New Phytol. 2002, 153, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Jansa, J.; Mozafar, A.; Frossard, E. Phosphorus acquisition strategies within arbuscular mycorrhizal fungal community of a single field site. Plant Soil 2005, 276, 163–176. [Google Scholar] [CrossRef]
- Feng, G.; Zhang, F.S.; Li, X.L.; Tian, C.Y.; Tang, C.; Rengel, Z. Improved tolereance of maize plants to salt stress by arbuscular mycorrhiza is related to higher accumulation of soluble sugars in roots. Mycorrhiza 2002, 12, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Li, J.; van der Werff, H. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1982. [Google Scholar]
- Luo, Q.; Xu, C.; Zheng, T.; Ma, Y.; Li, Y.; Zuo, Z. Leaf morphological and photosynthetic differences among four chemotypes of Cinnamomum camphora in different seasons. Ind. Crops Prod. 2021, 169, 113651. [Google Scholar] [CrossRef]
- Wang, J.P.; Wang, S.T.; Yue, J.M.; Zhang, J.C.; Zhang, L.; You, Y.H.; Zhao, W.R. Physiological response of Cinnamomum camphora seedlings to NaCl stress. Sci. Soil Water Conserv. 2016, 14, 82–89. [Google Scholar] [CrossRef]
- Yan, M.; Zhong, Z.C. Effects of aluminum stress on growth of Cinnamomum camphora seedligns inoculated with AMF. Sci. Silv. Sin. 2007, 43, 59–65. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, Y.L.; Song, L.L.; Song, X.Z.; Hänninen, H.; Wu, J.S. Biochar enhances nut quality of Torreya grandis and soil fertility under simulated nitrogen deposition. For. Ecol. Manag. 2017, 391, 321–329. [Google Scholar] [CrossRef]
- Wang, Y.H.; Liu, S.Y.; Shao, C.L.; Wu, A.P.; He, X.B.; Xia, L.N.; Wang, X.D.; Qiu, Y.J.; Yu, S.Q.; Pei, J.; et al. Enhancement of photosynthetic parameters and growth of Zelkova serrata by arbuscular mycorrhizal fungi under simulated sulfuric acid rain. Plant Ecol. 2021, 222, 1361–1374. [Google Scholar] [CrossRef]
- McNamara, N.P.; Black, H.I.J.; Beresford, N.A.; Parekh, N.R. Effects of acute gamma irradiation on chemical, physical and biological properties of soils. Appl. Soil Ecol. 2003, 24, 117–132. [Google Scholar] [CrossRef]
- Gong, Z. Chinese Soil Taxonomy; Science Press: Beijing, China, 2001. [Google Scholar]
- Qiu, Y.J.; Zhang, N.L.; Zhang, L.L.; Zhang, X.L.; Wu, A.P.; Huang, J.Y.; Yu, S.Q.; Wang, Y.H. Mediation of arbuscular mycorrhizal fungi on growth and biochemical parameters of Ligustrum vicaryi in response to salinity. Physiol. Mol. Plant Pathol. 2020, 112, 101522. [Google Scholar] [CrossRef]
- Xia, L.N.; Shao, C.L.; Zhang, N.L.; Wu, A.P.; Xie, J.B.; Qiu, Y.J.; He, X.B.; Pei, J.; Wang, X.D.; Wang, Y.H. Improved tolerance of mycorrhizal Torreya grandis seedlings to sulfuric acid rain related to phosphorus and zinc contents in shoots. J. Fungi 2021, 7, 296. [Google Scholar] [CrossRef]
- Wang, Y.H.; Wang, M.Q.; Li, Y.; Wu, A.P.; Huang, J.Y. Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress. PLoS ONE 2018, 13, e0196408. [Google Scholar] [CrossRef]
- Porter, W.M. The ‘most probable number’ method for enumerating infective propagules of vesicular arbuscular mycorrhizal fungi in soil. Aust. J. Soil Res. 1979, 17, 515–519. [Google Scholar] [CrossRef]
- Evelin, H.; Giri, B.; Kapoor, R. Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 2012, 22, 203–217. [Google Scholar] [CrossRef]
- Phillips, J.; Hayman, D. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific Publications: London, UK, 1989. [Google Scholar]
- Heidarianpour, M.B.; Aliasgharzad, N.; Olsson, P.A. Positive effects of co-inoculation with Rhizophagus irregularis and Serendipita indica on tomato growth under saline conditions, and their individual colonization estimated by signature lipids. Mycorrhiza 2020, 30, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.X.; Bai, R.; Nan, M.; Ren, W.; Wang, C.M.; Shabala, S.; Zhang, J.L. Evaluation of salt tolerance of oat cultivars and the mechanism of adaptation to salinity. J. Plant Physiol. 2022, 273, 153708. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Influence of arbuscular mycorrhizal fungi and salinity on growth, biomass and mineral nutrient of Acacia auriculiformis. Biol. Fert. Soils 2003, 38, 170–175. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; He, X.H. Contributions of arbuscular mycorrhizal fungi to growth, photosynthesis, root morphology and ion balance of citrus seedlings under salt stress. Acta Physiol. Plant 2010, 32, 297–304. [Google Scholar] [CrossRef]
- Otgonsuren, B.; Rewald, B.; Godbold, D.L.; Göransson, H. Ectomycorrhizal inoculation of Populus nigra modifies the response of absorptive root respiration and root surface enzyme activity to salinity stress. Flora 2016, 224, 123–129. [Google Scholar] [CrossRef]
- McMillen, B.G.; Juniper, S.; Abbott, L.K. Inhibition of hyphal growth of a vesicular-arbuscular mycorrhizal fungus in soil containing sodium chloride limits the spread of infection from spores. Soil Biol. Biochem. 1998, 30, 1639–1646. [Google Scholar] [CrossRef]
- Pandey, D.; Kehri, H.K.; Zoomi, I.; Akhtar, O.; Singh, A.K. Mycorrhizal fungi: Biodiversity, ecological significance, and industrial applications. In Recent Advancement in White Biotechnology through Fungi: Volume 1: Diversity and Enzymes Perspectives; Yadav, A.N., Mishra, S., Singh, S., Gupta, A., Eds.; Springer: Cham, Switzerland, 2019; pp. 181–199. ISBN 978-030-25506-0. [Google Scholar]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Daei, G.; Ardekani, M.R.; Rejali, F.; Teimuri, S.; Miransari, M. Alleviation of salinity stress on wheat yield, yield components, and nutrient uptake using arbuscular mycorrhizal fungi under field conditions. J. Plant Physiol. 2009, 166, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Hajiboland, R.; Aliasgharzadeh, N.; Laiegh, S.F.; Poschenrieder, C. Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant Soil 2010, 331, 313–327. [Google Scholar] [CrossRef]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Improved tolerance of Acacia nilotica to salt stress by arbuscular mucorrhiza, Glomus fasciculatum, may be partly related to elevated K+/Na+ ratios in root and shoot tissue. Microb. Ecol. 2007, 54, 753–760. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Porcel, R.; Azcon, C.; Aroca, R. Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: New challenges in physiological and molecular studies. J. Exp. Bot. 2012, 63, 4033–4044. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.; He, Y.; Zang, L.; Si, J.; Yang, Y.; Shen, K.; Xia, T.; Tan, Q.; Wu, B.; Guo, Y.; et al. Mycorrhizal networks interacting with litter improves nutrients and growth for one plant through the vary of N/P ratio under karst soil. Phyton Int. J. Exp. Bot. 2021, 90, 701–717. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcón, R. Symbiotic efficiency and infectivity of an autochthonous arbuscular mycorrhizal Glomus sp. from saline soils and Glomus deserticola under salinity. Mycorrhiza 2000, 10, 137–143. [Google Scholar] [CrossRef]
- Johnson, N.C.; Wilson, G.W.T.; Wilson, J.A.; Miller, R.M.; Bowker, M.A. Mycorrhizal phenotypes and the law of the minimum. New Phytol. 2015, 205, 1473–1484. [Google Scholar] [CrossRef]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2009, 98, 139–151. [Google Scholar] [CrossRef]
- He, Y.; Cornelissen, J.H.; Zhong, Z.; Dong, M.; Jiang, C. How interacting fungal species and mineral nitrogen inputs affect transfer of nitrogen from litter via arbuscular mycorrhizal mycelium. Environ. Sci. Pollut. Res. 2017, 24, 9791–9801. [Google Scholar] [CrossRef]
- Hajiboland, R. Role of arbuscular mycorrhiza in amelioration of salinity. In Salt Stress in Plants: Singnaling, Omics and Adaptations; Ahmad, P., Azzoz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 301–354. ISBN 978-1-4614-6108-1. [Google Scholar]
- van der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, T.; Wu, A.; Li, Y.; Zhang, N. Mycorrhizal Benefits of Salt-Stressed Cinnamomum camphora (L.) Presl. May Be Related to P and Mn2+ Contents in Shoots, Biomass Allocation, and K+/Na+ in Roots and Shoots. Forests 2022, 13, 1882. https://doi.org/10.3390/f13111882
Wang Y, Li T, Wu A, Li Y, Zhang N. Mycorrhizal Benefits of Salt-Stressed Cinnamomum camphora (L.) Presl. May Be Related to P and Mn2+ Contents in Shoots, Biomass Allocation, and K+/Na+ in Roots and Shoots. Forests. 2022; 13(11):1882. https://doi.org/10.3390/f13111882
Chicago/Turabian StyleWang, Yanhong, Tiantian Li, Aiping Wu, Yan Li, and Naili Zhang. 2022. "Mycorrhizal Benefits of Salt-Stressed Cinnamomum camphora (L.) Presl. May Be Related to P and Mn2+ Contents in Shoots, Biomass Allocation, and K+/Na+ in Roots and Shoots" Forests 13, no. 11: 1882. https://doi.org/10.3390/f13111882