
Douglas Fir Growth Is Constrained by Drought: Delineating the Climatic Limits of Timber Species under Seasonally Dry Conditions
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
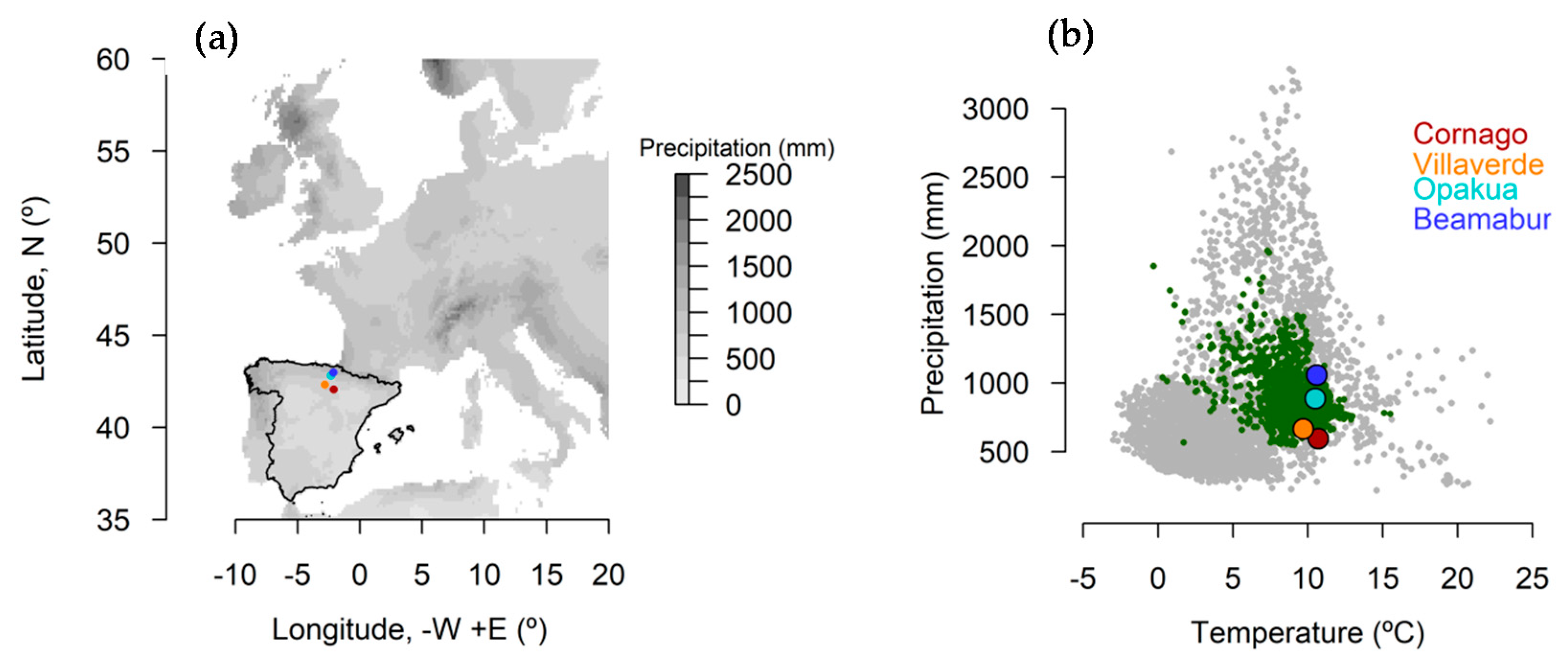
2.1. Study Sites
2.2. Field Sampling
2.3. Tree-Ring Width Data
2.4. Climate and Drought Data
2.5. Building Tree-Ring Width Chronologies
2.6. Climate–Growth Relationships at Tree Level
2.7. Wood Density Data
2.8. Relationships between Climate and Tree-Ring Width or Density Series
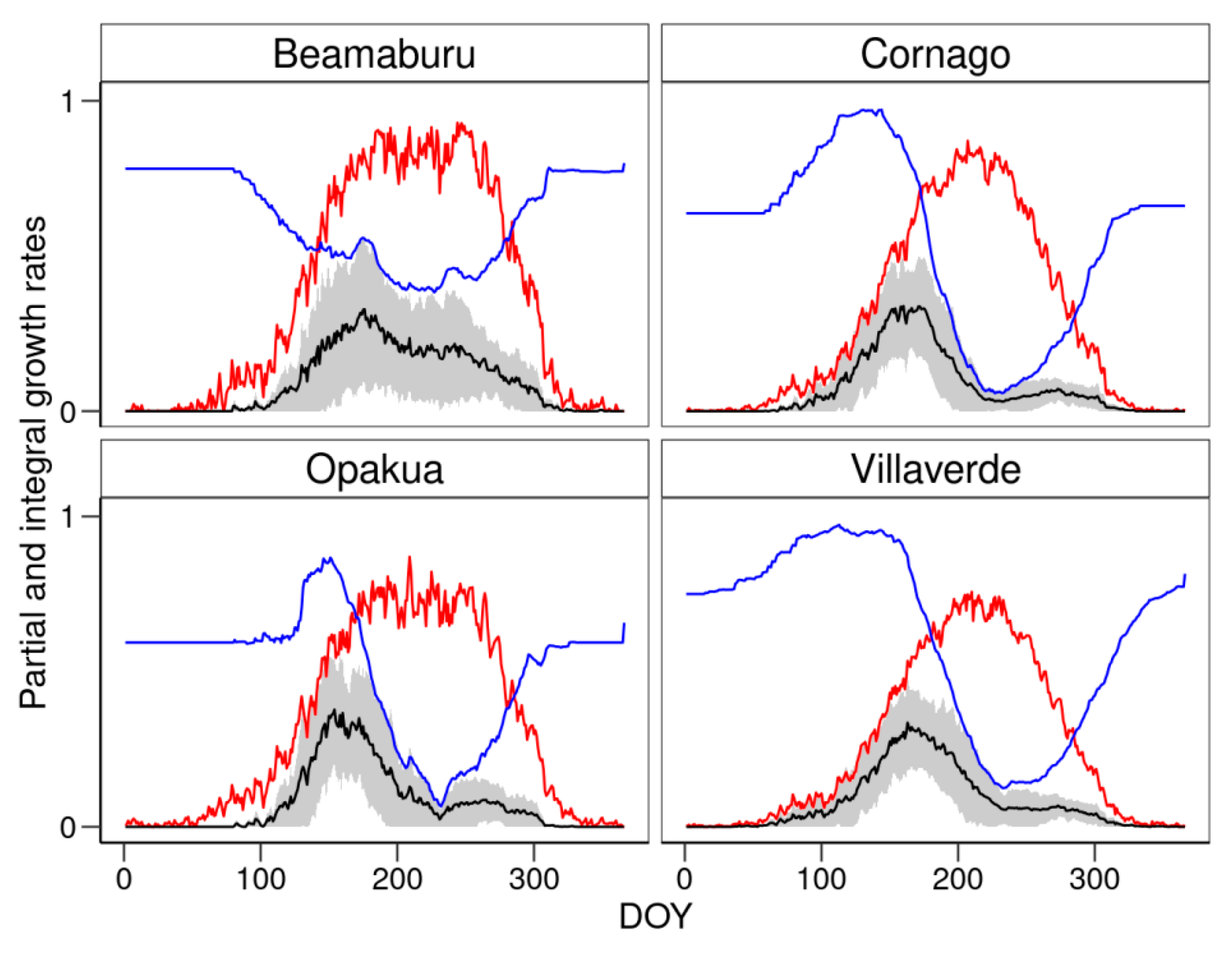
2.9. Process-Based Growth Model
2.10. Statistical Analyses
3. Results
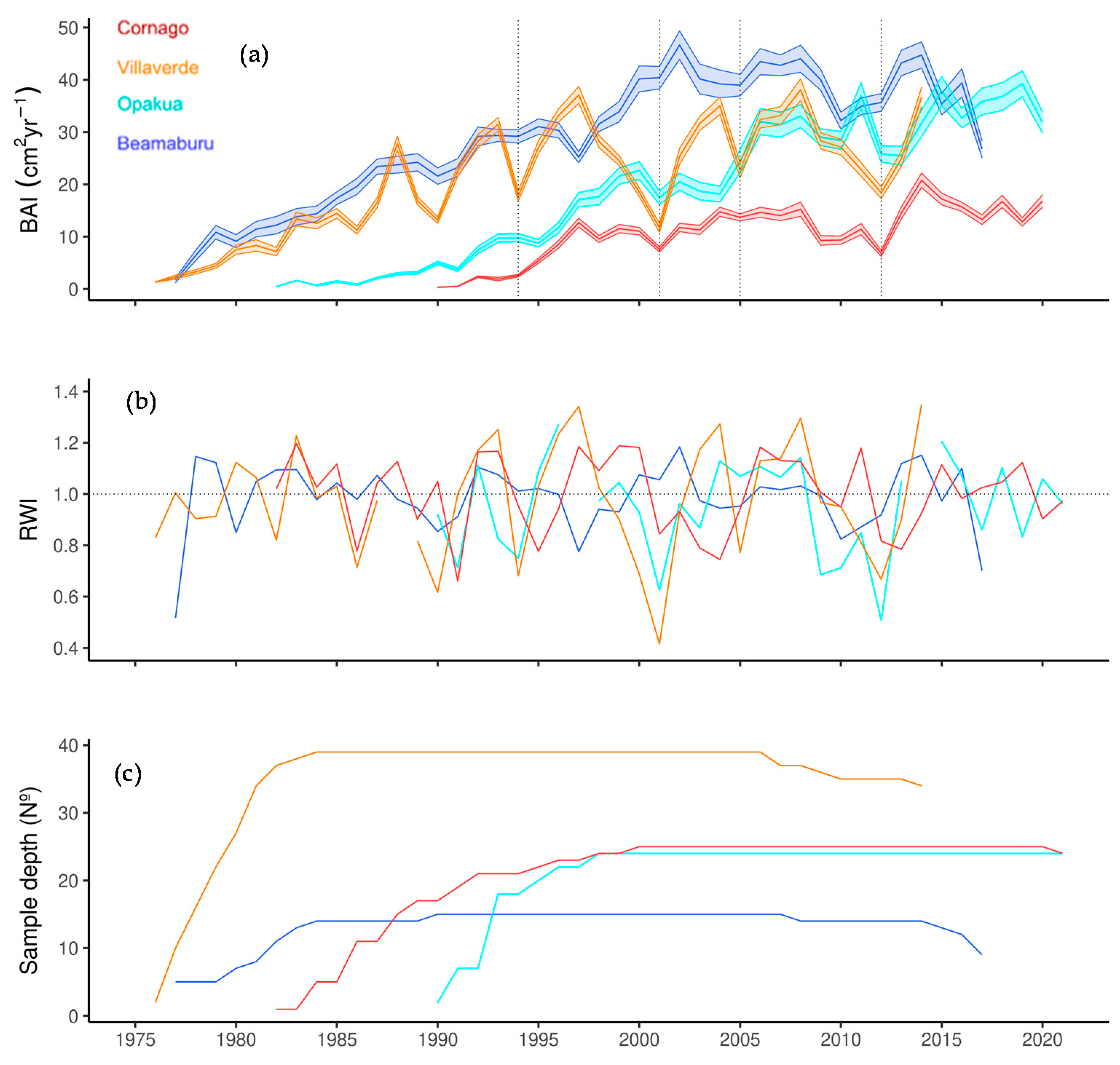
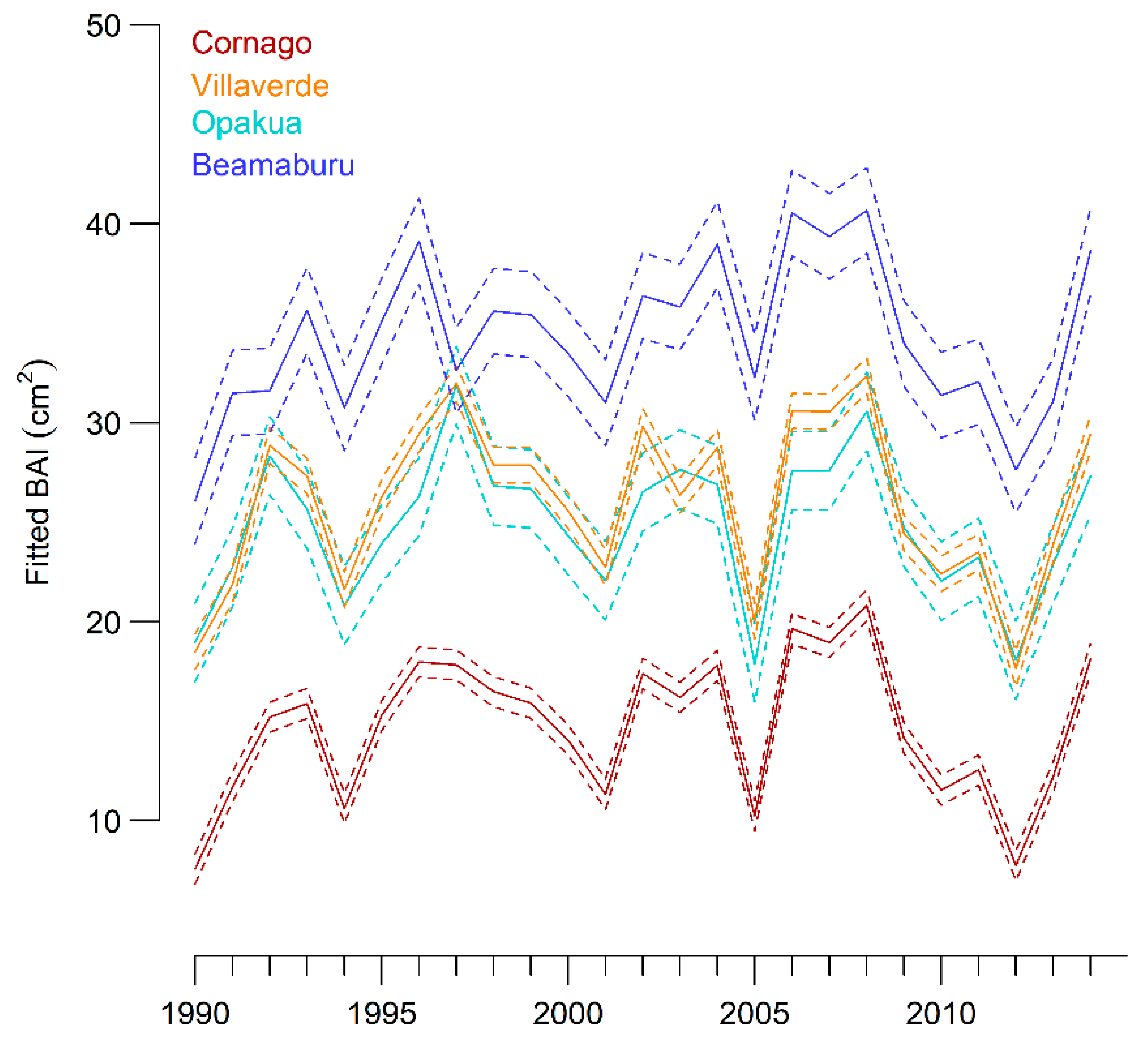
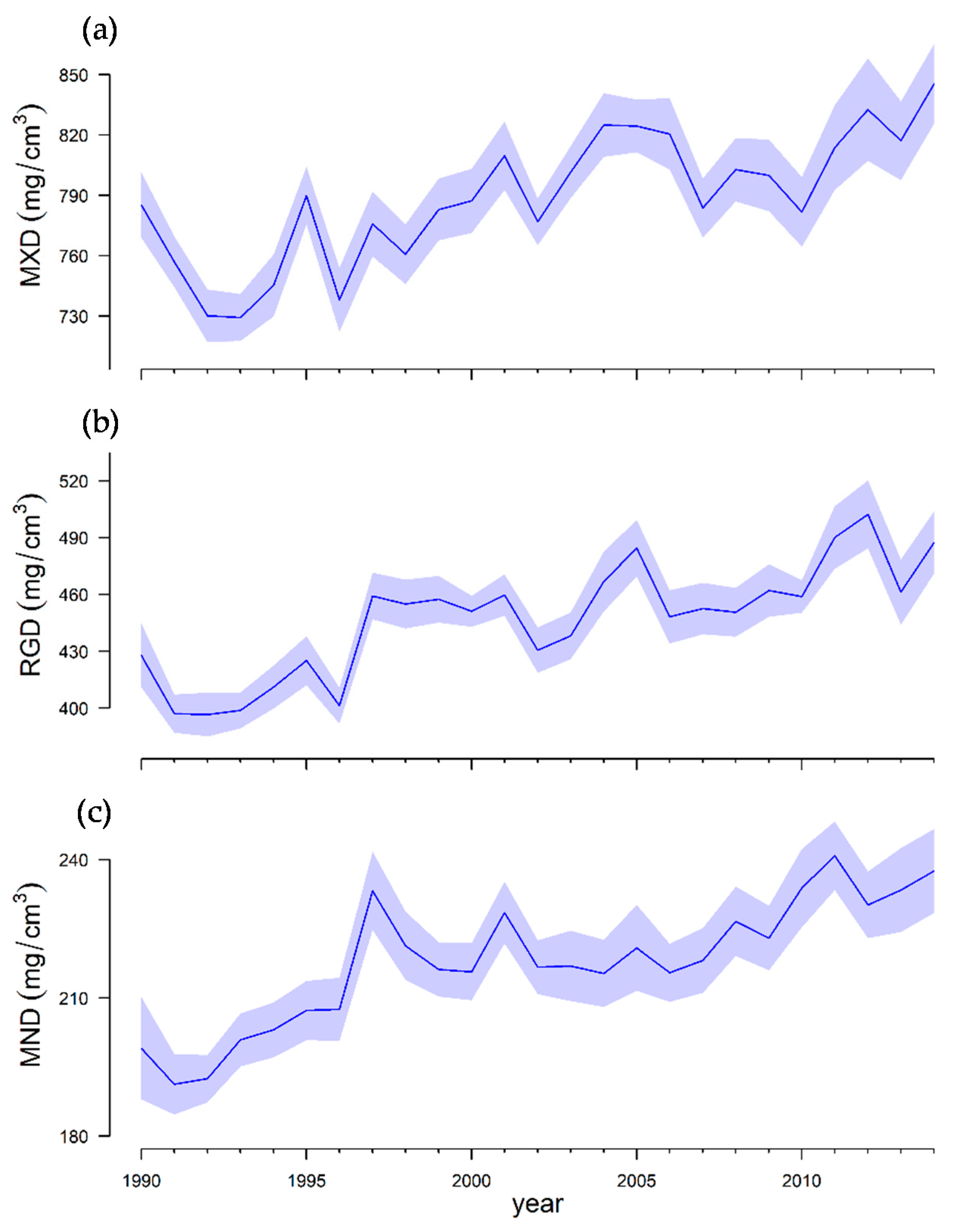
3.1. Growth Variability between Sites and Years
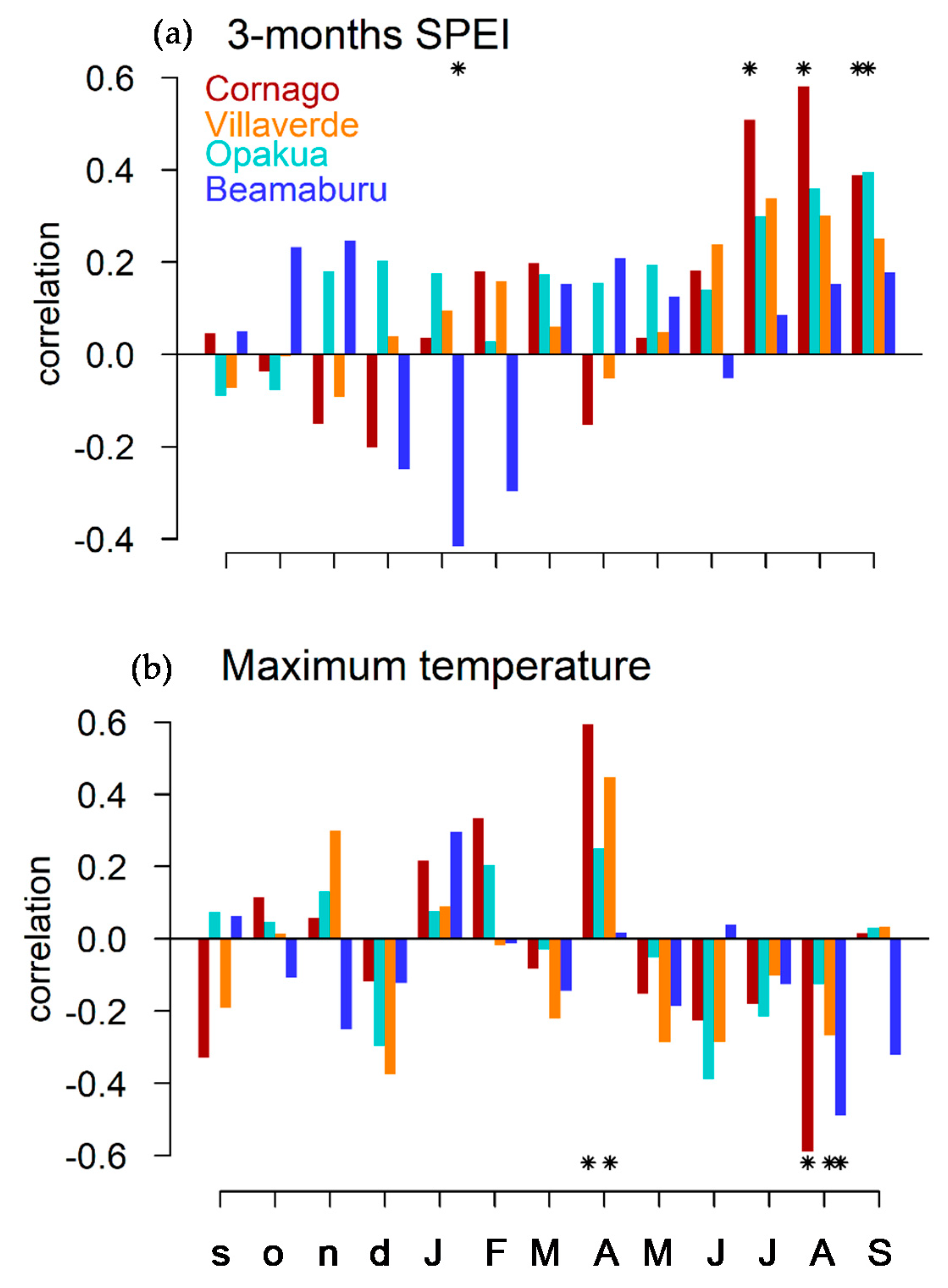
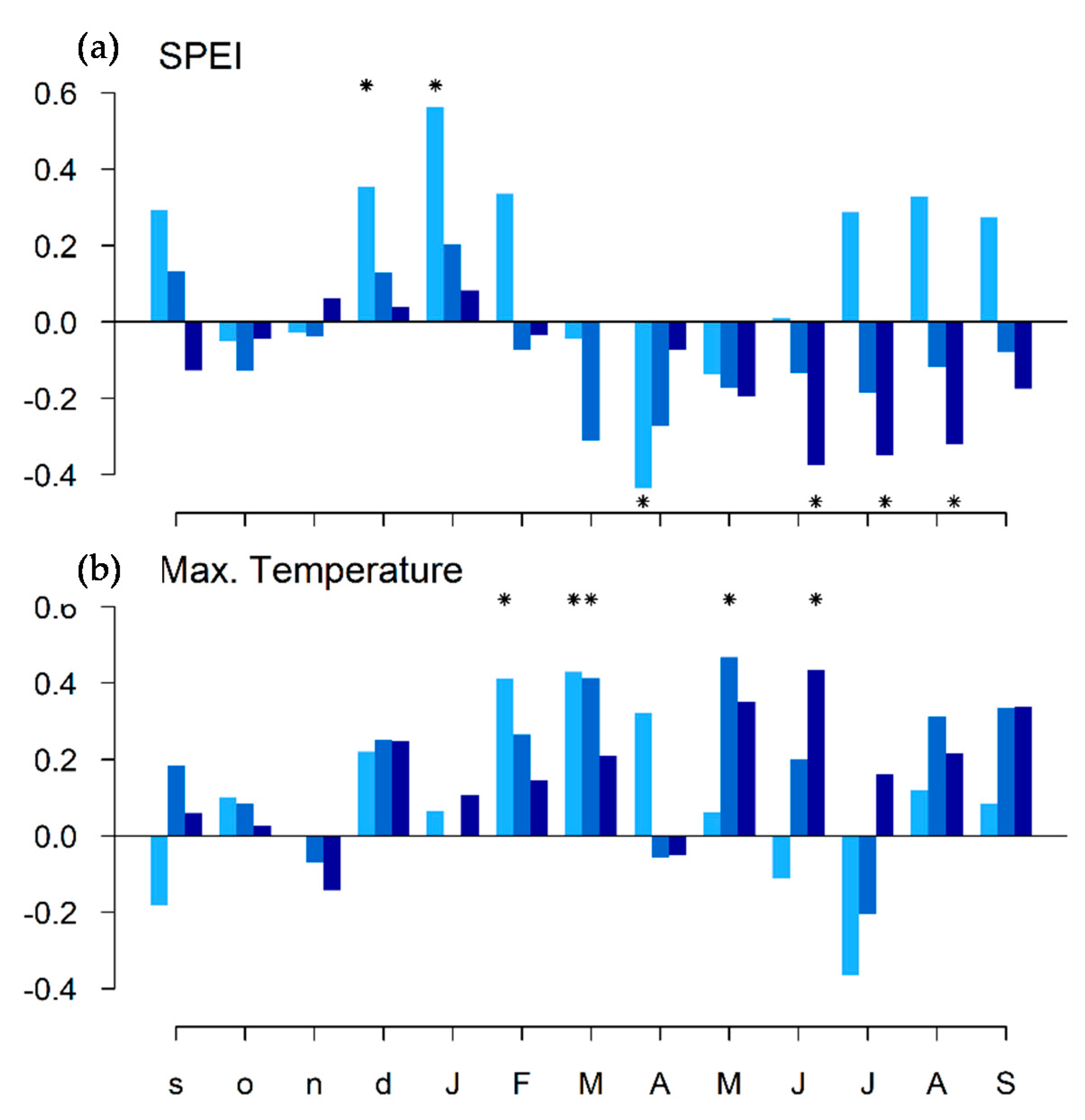
3.2. Relationships between Tree Growth and Climate
3.3. Relationships between Wood Density and Climate
3.4. Climatic Limitations of Tree Growth
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- García-Valdés, R.; Estrada, A.; Early, R.; Lehsten, V.; Morin, X. Climate change impacts on long-term forest productivity might be driven by species turnover rather than by changes in tree growth. Glob. Ecol. Biogeogr. 2020, 29, 1360–1372. [Google Scholar] [CrossRef]
- Buras, A.; Menzel, A. Projecting tree species composition changes of European forests for 2061–2090 under RCP 4.5 and RCP 8.5 scenarios. Front. Plant Sci. 2019, 9, 1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiecker, H. Executive Summary. In Douglas-fir—An option for Europe; Spiecker, H., Lindner, M., Schuler, J., Eds.; European Forest Institute: Joensuu, Finland, 2019; pp. 21–31. [Google Scholar]
- Martínez-Sancho, E.; Rellstab, C.; Guillaume, F.; Bigler, C.; Fonti, P.; Wohlgemuth, T.; Vitasse, Y. Post-glacial re-colonization and natural selection have shaped growth responses of silver fir across Europe. Sci. Tot. Env. 2021, 779, 146393. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J. x Global change and forest disturbances in the Mediterranean Basin: Breakthroughs, knowledge gaps, and recommendations. Forests 2021, 12, 603. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Vergarechea, M.; Alfaro-Sánchez, R.; Cattaneo, N.; Vicente-Serrano, S.M. Tree growth is more limited by drought in rear-edge forests most of the times. For. Ecosyst. 2021, 8, 25. [Google Scholar] [CrossRef]
- Babst, F.; Bouriaud, O.; Poulter, B.; Trouet, V.; Girardin, M.P.; Frank, D.C. Twentieth century redistribution in climatic drivers of global tree growth. Sci. Adv. 2019, 5, eaat4313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the anthropocene. Ecosphere 2015, 6, 129. [Google Scholar] [CrossRef]
- Buras, A.; Rammig, A.; Zang, C.S. Quantifying impacts of the 2018 drought on European ecosystems in comparison to 2003. Biogeosciences 2020, 17, 1655–1672. [Google Scholar] [CrossRef] [Green Version]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Change Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Pötzelsberger, E.; Spiecker, H.; Neophytou, C.; Mohren, F.; Gazda, A.; Hasenauer, H. Growing non-native trees in European forests brings benefits and opportunities but also has its risks and limits. Curr. For. Rep. 2020, 6, 339–353. [Google Scholar] [CrossRef]
- Brus, R.; Pötzelsberger, E.; Lapin, K.; Brundu, G.; Orazio, C.; Straigyte, L.; Hasenauer, H. Extent, distribution and origin of non-native forest tree species in Europe. Scand. J. For. Res. 2019, 34, 533–544. [Google Scholar] [CrossRef]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Ch. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Pautasso, M.; & Holdenrieder, O. Ecological consequences of Douglas fir (Pseudotsuga menziesii) cultivation in Europe. Eur. J. For. Res. 2014, 133, 13–29. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.M.; Rzepecki, A.; Werner, W. Non-native Douglas fir (Pseudotsuga menziesii) in Central Europe: Ecology, performance and nature conservation. For. Ecol. Manag. 2022, 506, 119956. [Google Scholar] [CrossRef]
- Sergent, A.-S.; Rozenberg, P.; Bréda, N. Douglas-fir is vulnerable to exceptional and recurrent drought episodes and recovers less well on less fertile sites. Ann. For. Sci. 2014, 71, 697–708. [Google Scholar] [CrossRef]
- Bastien, J.-C. Douglas Fir Distribution in Europe. In Douglas-fir—An option for Europe; Spiecker, H., Lindner, M., Schuler, J., Eds.; European Forest Institute: Joensuu, Finland, 2019; pp. 21–31. [Google Scholar]
- Klesse, S.; DeRose, R.J.; Babst, F.; Black, B.A.; Anderegg, L.D.L.; Axelson, J.; Ettinger, A. Continental-scale tree-ring-based projection of Douglas-fir growth: Testing the limits of space-for-time substitution. Glob. Ch. Biol. 2020, 26, 5146–5163. [Google Scholar] [CrossRef]
- Castaldi, C.; Marchi, M.; Vacchiano, G.; Corona, P. Douglas-fir climate sensitivity at two contrasting sites along the southern limit of the European planting range. J. For. Res. 2020, 31, 2193–2204. [Google Scholar] [CrossRef] [Green Version]
- Dalla Salda, G.; Martinez-Meier, A.; Cochard, H.; Rozenberg, P. Variation of wood density and hydraulic properties of Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) clones related to a heat and drought waves in France. For. Ecol. Manag. 2009, 257, 182–189. [Google Scholar] [CrossRef]
- Chauvin, T.; Cochard, H.; Segura, V.; Rozenberg, P. Native-source climate determines the Douglas-fir potential of adaptation to drought. For. Ecol. Manage. 2019, 444, 9–20. [Google Scholar] [CrossRef]
- Vejpustková, M.; Čihák, T. Climate response of Douglas fir reveals recently increased sensitivity to drought stress in Central Europe. Forests 2019, 10, 97. [Google Scholar] [CrossRef]
- Valeriano, C.; Gazol, A.; Colangelo, M.; González de Andrés, E.; Camarero, J.J. Modeling climate impacts on tree growth to assess tree vulnerability to drought during forest dieback. Front. Plant Sci. 2021, 26, 1810. [Google Scholar] [CrossRef] [PubMed]
- Björklund, J.; Seftigen, K.; Schweingruber, F.; Fonti, P.; von Arx, G.; Bryukhanova, M.V.; Cuny, H.E.; Carrer, M.; Castagneri, D.; Frank, D.C. Cell size and wall dimensions drive distinct variability of earlywood and latewood density in Northern Hemisphere conifers. New Phytol. 2017, 216, 728–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathgeber, C.B.K. Conifer tree-ring density interannual variability—Anatomical, physiological and environmental determinants. New Phytol. 2017, 216, 621–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellizzari, E.; Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Carrer, M. Wood anatomy and carbon-isotope discrimination support long-term hydraulic deterioration as a major cause of drought-induced dieback. Glob. Ch. Biol. 2016, 22, 2125–2137. [Google Scholar] [CrossRef] [PubMed]
- Little, E. Atlas of United States Trees. Volume 1. Conifers and Important Hardwoods; Department of Agriculture, Forest Service: Washington, DC, USA, 1971. [Google Scholar]
- Brus, D.J.; Hengeveld, G.M.; Walvoort, D.J.J.; Goedhart, P.W.; Heidema, A.H.; Nabuurs, G.J.; Gunia, K. Statistical mapping of tree species over Europe. Eur. J. For. Res. 2012, 131, 145–157. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente-Serrano, S.M.; Lopez-Moreno, J.I.; Beguería, S.; Lorenzo-Lacruz, J.; Sanchez-Lorenzo, A.; García-Ruiz, J.M.; Azorin-Molina, C.; Morán-Tejeda, E.; Revuelto, J.; Trigo, R.; et al. Evidence of increasing drought severity caused by temperature rise in southern Europe. Env. Res. Lett. 2014, 9, 044001. [Google Scholar] [CrossRef]
- Klesse, S. Critical note on the application of the “two-third” spline. Dendrochronologia 2021, 65, 125786. [Google Scholar] [CrossRef]
- Hevia, A.; Sánchez-Salguero, R.; Camarero, J.J.; Querejeta, J.I.; Sangüesa-Barreda, G.; Gazol, A. Long-term nutrient imbalances linked to drought-triggered forest dieback. Sci. Tot. Env. 2019, 690, 1254–1267. [Google Scholar] [CrossRef]
- Camarero, J.J.; Rozas, V.; Olano, J.M. Minimum wood density of Juniperus thurifera is a robust proxy of spring water availability in a continental Mediterranean climate. J. Biogeogr. 2014, 41, 1105–1114. [Google Scholar] [CrossRef]
- Camarero, J.J.; Fernández-Pérez, L.; Kirdyanov, A.V.; Shestakova, T.A.; Knorre, A.A.; Kukarskih, V.V.; Voltas, J. Minimum wood density of conifers portrays changes in early season precipitation at dry and cold Eurasian regions. Trees Struct. Funct. 2017, 7, 214–215. [Google Scholar] [CrossRef] [Green Version]
- Zang, C.; Biondi, F. treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2015, 38, 431–436. [Google Scholar] [CrossRef]
- Shishov, V.V.; Tychkov, I.I.; Popkova, M.I.; Ilyin, V.A.; Bryukhanova, M.V.; Kirdyanov, A.V. VS-oscilloscope: A new tool to parameterize tree radial growth based on climate conditions. Dendrochronologia 2016, 39, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Tychkov, I.I.; Sviderskaya, I.V.; Babushkina, E.A.; Popkova, M.I.; Vaganov, E.A.; Shishov, V.V. How can the parameterization of a process-based model help us understand real tree-ring growth? Trees Struct. Funct. 2019, 33, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Tumajer, J.; Kašpar, J.; Kuželová, H.; Shishov, V.V.; Tychkov, I.I.; Popkova, M.I.; Vaganov, E.A.; Treml, V. Forward modeling reveals multidecadal trends in cambial kinetics and phenology at treeline. Front. Plant Sci. 2021, 12, 32. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000. [Google Scholar]
- Mehtätalo, L.; Peltola, H.; Kilpeläinen, A.; Ikonen, V.P. The response of basal area growth of Scots pine to thinning: A longitudinal analysis of tree-specific series using a nonlinear mixed-effects model. For. Sci. 2014, 60, 636–644. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference, a Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed effects models and extensions in ecology with R. Springer: New York, NY, USA, 2009. [Google Scholar]
- Nakagawa, S.; Johnson, P.C.D.; Schielzeth, H. The coefficient of determination R2 and intra-class correlation coefficient from generalized linear mixed-effects models revisited and expanded. J. R. Soc. Interf. 2017, 14, 20170213. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R. RStudio, PBC; RStudio Team: Boston, MA, USA, 2021. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C. dplR: Dendrochronology Program Library in R. R package version 1.7.2. 2021. Available online: https://CRAN.R-project.org/package=dplR (accessed on 22 August 2022).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–51. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1.42.1. 2018. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 8 June 2020).
- Length, R.V. emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.7.2. 2022. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 22 August 2022).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SAGE Publications, Inc: Thousand Oaks, CA, USA.
- Eckhart, T.; Pötzelsberger, E.; Koeck, R.; Thom, D.; Lair, G.J.; van Loo, M.; Hasenauer, H. Forest stand productivity derived from site conditions: An assessment of old Douglas-fir stands (Pseudotsuga menziesii (Mirb.) Franco var. menziesii) in Central Europe. Ann. For. Sci. 2019, 76, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretzsch, H. The course of tree growth. Theory and reality. For. Ecol. Manage. 2020, 478, 118508. [Google Scholar]
- Gazol, A.; Camarero, J.J.; Vicente-Serrano, S.M.; Sánchez-Salguero, R.; Gutiérrez, E.; de Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; Tíscar, P.A.; et al. Forest resilience to drought varies across biomes. Glob. Change Biol. 2018, 24, 2143–2158. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Cantero, A.; Sánchez-Salguero, R.; Sánchez-Miranda, A.; Granda, E.; Serra-Maluquer, X.; Ibáñez, R. Forest growth responses to drought at short-and long-term scales in Spain: Squeezing the stress memory from tree rings. Front. Ecol. Evol. 2018, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, S.; Pereira, H. Characterization of Douglas-fir grown in Portugal: Heartwood, sapwood, bark, ring width and taper. Eur. J. For. Res. 2017, 136, 597–607. [Google Scholar] [CrossRef]
- Gilson, L.W.; Maguire, D.A. Drivers of productivity differences between Douglas-fir planted within its native range in Oregon and on exotic sites in New Zealand. For. Ecol. Manage. 2021, 498, 119525. [Google Scholar] [CrossRef]
- Thurm, E.A.; Uhl, E.; Pretzsch, H. Mixture reduces climate sensitivity of Douglas-fir stem growth. For. Ecol. Manage. 2016, 376, 205–220. [Google Scholar] [CrossRef]
- Dalla-Salda, G.; Martinez-Meier, A.; Cochard, H.; Rozenberg, P. Genetic variation of xylem hydraulic properties shows that wood density is involved in adaptation to drought in Douglas-fir (Pseudotsuga menziesii (Mirb.)). Ann. For. Sci. 2011, 68, 747–757. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Latitude N | Longitude W | Elevation (m a.s.l.) | Temperature (°C) | Precipitation (mm) | Main Lithology |
|---|---|---|---|---|---|---|
| Villaverde | 42.321 | 2.812 | 886 | 9.8 | 640 | Shale |
| Opakua | 42.818 | 2.323 | 1023 | 9.9 | 990 | Conglomerates |
| Beamaburu | 42.985 | 2.149 | 880 | 12.0 | 1110 | Limestone |
| Cornago | 42.065 | 2.107 | 1023 | 12.1 | 570 | Shales |
| Site | No. Cores (No. Trees) | Period | Autocorrelation | Ring Width (mm) | Rbar | EPS |
|---|---|---|---|---|---|---|
| Villaverde | 39 (20) | 1976–2014 | 0.53 ± 0.17 | 4.49 ± 1.71 b | 0.67 | 0.89 |
| Opakua | 25 (13) | 1982–2021 | 0.57 ± 0.21 | 4.58 ± 1.31 b | 0.52 | 0.87 |
| Beamaburu | 15 (15) | 1977–2017 | 0.60 ± 0.26 | 5.20 ± 1.80 b | 0.47 | 0.85 |
| Cornago | 24 (12) | 1990–2021 | 0.64 ± 0.25 | 3.57 ± 1.25 a | 0.74 | 0.90 |
| Variable | Estimate | Std. Error | df | t Value | p |
|---|---|---|---|---|---|
| Intercept | 34.276 | 1.726 | 74.631 | 19.860 | 0.000 |
| Cornago | −19.584 | 2.684 | 83.189 | −7.297 | 0.000 |
| Opakua | −9.413 | 2.402 | 64.290 | −3.919 | 0.000 |
| Villaverde | −8.287 | 2.117 | 59.871 | −3.914 | 0.000 |
| SPEI | −0.487 | 0.891 | 44.433 | −0.546 | 0.588 |
| Tmax | −2.481 | 0.871 | 41.934 | −2.847 | 0.007 |
| Age | 5.740 | 0.919 | 51.780 | 6.243 | 0.000 |
| Cornago: age | −0.865 | 0.875 | 1387.561 | −0.988 | 0.323 |
| Opakua: age | 4.647 | 0.781 | 1399.288 | 5.948 | 0.000 |
| Villaverde: age | −3.607 | 0.695 | 1388.601 | −5.189 | 0.000 |
| Cornago: SPEI | 1.625 | 0.774 | 1378.864 | 2.099 | 0.036 |
| Opakua: SPEI | 2.948 | 0.734 | 1366.048 | 4.018 | 0.000 |
| Villaverde:SPEI | 2.576 | 0.649 | 1367.387 | 3.971 | 0.000 |
| Cornago: Tmax | 1.051 | 0.729 | 1375.562 | 1.440 | 0.150 |
| Opakua: Tmax | 2.566 | 0.708 | 1362.926 | 3.626 | 0.000 |
| Villaverde: Tmax | 1.058 | 0.629 | 1375.644 | 1.683 | 0.093 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gazol, A.; Valeriano, C.; Cantero, A.; Vergarechea, M.; Camarero, J.J. Douglas Fir Growth Is Constrained by Drought: Delineating the Climatic Limits of Timber Species under Seasonally Dry Conditions. Forests 2022, 13, 1796. https://doi.org/10.3390/f13111796
Gazol A, Valeriano C, Cantero A, Vergarechea M, Camarero JJ. Douglas Fir Growth Is Constrained by Drought: Delineating the Climatic Limits of Timber Species under Seasonally Dry Conditions. Forests. 2022; 13(11):1796. https://doi.org/10.3390/f13111796
Chicago/Turabian StyleGazol, Antonio, Cristina Valeriano, Alejandro Cantero, Marta Vergarechea, and Jesús Julio Camarero. 2022. "Douglas Fir Growth Is Constrained by Drought: Delineating the Climatic Limits of Timber Species under Seasonally Dry Conditions" Forests 13, no. 11: 1796. https://doi.org/10.3390/f13111796