
Microcosm Study on Allelopathic Effects of Leaf Litter Leachates and Purified Condensed Tannins from Kandelia obovata on Germination and Growth of Aegiceras corniculatum

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Sampling Site
2.2. Plant Materials and Culture Conditions
2.3. Preparation for Allelopathic Agonists
2.3.1. Leaf Litter Leachates
2.3.2. Purified Condensed Tannins
2.4. Experimental Design
2.4.1. Treatments
2.4.2. Establishment of Physiological Indices
2.5. Statistical Analysis
3. Results
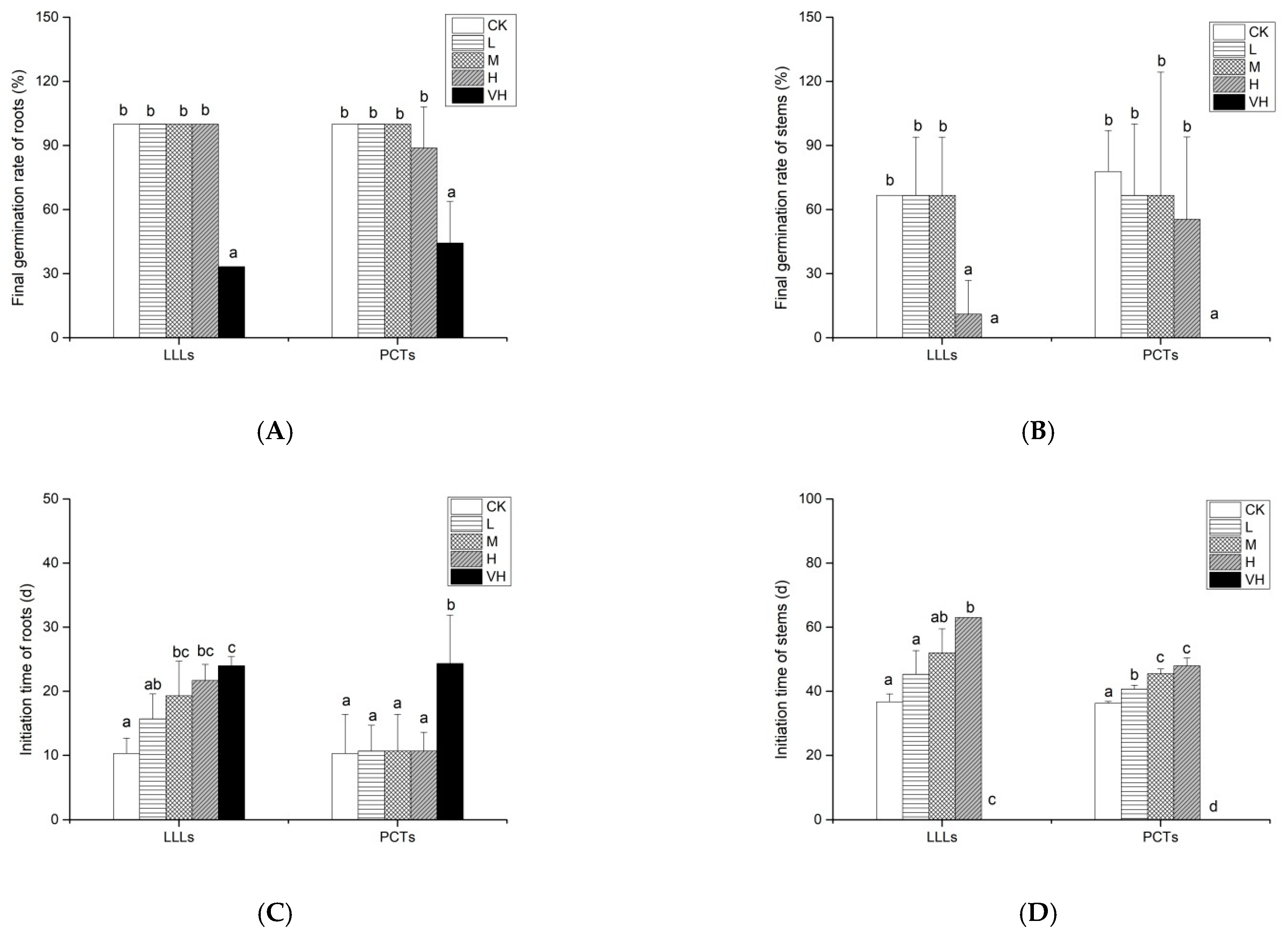
3.1. Effects of LLLs and PCTs from K. obovata on Final Germination Rates and Initiation Times of A. corniculatum Roots and Stems
3.2. Effects of LLLs and PCTs from K. obovata on the Numbers of A. corniculatum Fine Roots, Nutritive Roots, and Leaves and the Lengths of A. corniculatum Fine Roots, Nutritive Roots, and Stems
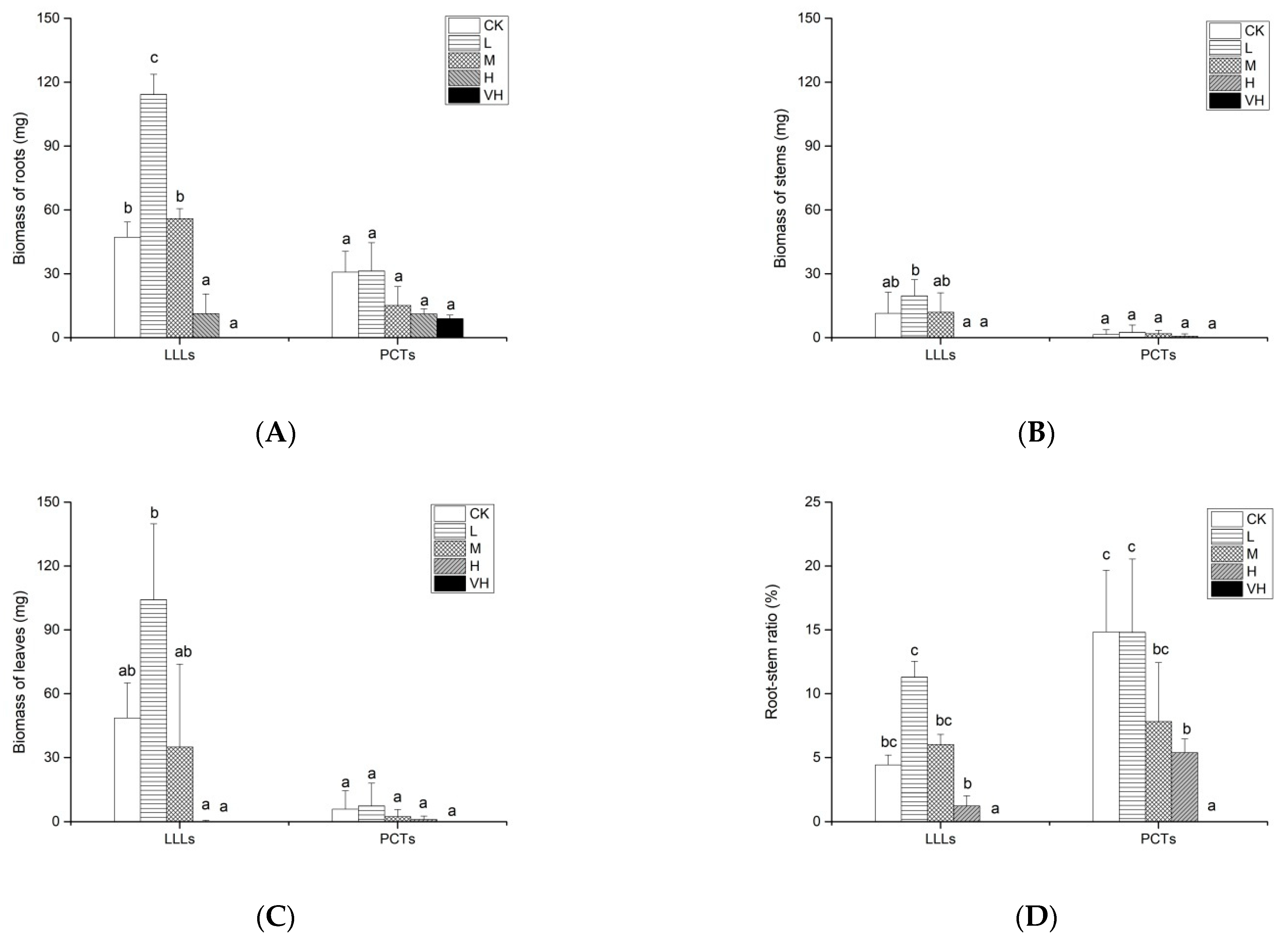
3.3. Effects of LLLs and PCTs from K. obovata on Biomasses of A. corniculatum Roots, Stems, Leaves, and Root–Stem Ratio
3.4. Allelopathic Response Indices of A. corniculatum’s Physiological Performance under LLL and PCT Treatments from K. obovata
3.5. Distinguishing Effects between LLL and PCT Treatments from K. obovata on Physiological Performance of A. corniculatum
4. Discussion
4.1. Condensed Tannins Extracted from Leaf Litter Primarily Contribute to Allelopathy in Mangrove Forests
4.2. A Challenge to Utilize Allelopathic Mechanisms in Mangrove Rehabilitation or Restoration and Conservation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- May, F.E.; Ash, J.E. An assessment of the allelopathic potential of eucalyptus. Aust. J. Bot. 1990, 38, 245–254. [Google Scholar] [CrossRef]
- Mushtaq, M.N.; Sunohara, Y.; Matsumoto, H. Allelochemical L-DOPA induces quinoprotein adducts and inhibits NADH dehydrogenase activity and root growth of cucumber. Plant Physiol. Biochem. 2013, 70, 374–378. [Google Scholar] [CrossRef]
- Si, C.C.; Liu, X.Y.; Wang, C.Y.; Wang, L.; Dai, Z.C.; Qi, S.S.; Du, D.L. Different degrees of plant invasion significantly affect the richness of the soil fungal community. PLoS ONE 2013, 8, e85490. [Google Scholar] [CrossRef]
- Das, R.; Geethangili, M.; Majhi, A.; Das, B.; Rao, Y.K.; Tzeng, Y.M. A new highly oxygenated pseudoguaianolide from a collection of the flowers of Parthenium hysterophorus. Chem. Pharm. Bull. 2005, 53, 861–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef]
- Meiners, S.J.; Kong, C.H.; Ladwig, L.M.; Pisula, N.L.; Lang, K.A. Developing an ecological context for allelopathy. Plant Ecol. 2012, 213, 1861–1867. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Vitousek, P. The role of polyphenols in terrestrial ecosystem nutrient cycling. Trends Ecol. Evol. 2000, 15, 238–243. [Google Scholar] [CrossRef]
- Fabbro, C.D.; Güsewell, S.; Prati, D. Allelopathic effects of three plant invaders on germination of native species: A field study. Biol. Invasions 2014, 16, 1035–1042. [Google Scholar] [CrossRef]
- Stamp, N. Out of the quagmire of plant defense hypotheses. Q. Rev. Biol. 2003, 78, 23–55. [Google Scholar] [CrossRef] [Green Version]
- Wardle, D.A.; Karban, R.; Callaway, R.M. The ecosystem and evolutionary contexts of allelopathy. Trends Ecol. Evol. 2011, 26, 655–662. [Google Scholar]
- Torawane, S.; Mokat, D. Allelopathic effects of weed Neanotis montholonii on seed germination and metabolism of mungbean and rice. Allelopath. J. 2020, 49, 151–164. [Google Scholar]
- Wang, D.; Chen, J.; Xiong, X.; Wang, S.; Liu, J. Allelopathic effects of Cinnamomum migao on seed germination and seedling growth of its associated species Liquidambar formosana. Forests 2019, 10, 535. [Google Scholar] [CrossRef] [Green Version]
- Legare, S.; Bergeron, Y.; Pare, D. Effect of aspen (Populus tremuloides) as a companion species on the growth of black spruce (Picea mariana) in the southwestern boreal forest of Quebec. For. Ecol. Manag. 2005, 208, 211–222. [Google Scholar]
- Bayen, S. Occurrence, bioavailability and toxic effects of trace metals and organic contaminants in mangrove ecosystems: A review. Environ. Int. 2012, 48, 84–101. [Google Scholar] [CrossRef] [PubMed]
- Brander, L.M.; Wagtendonk, A.J.; Hussain, S.S.; McVittie, A.; Verburg, P.H.; de Groot, R.S.; van der Ploeg, S. Ecosystem service values for mangroves in Southeast Asia: A meta-analysis and value transfer application. Ecosyst. Serv. 2012, 1, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Lovelock, C.E.; Cahoon, D.R.; Friess, D.A.; Guntenspergen, G.R.; Krauss, K.W.; Reef, R.; Rogers, K.; Saunders, M.L.; Sidik, F.; Swales, A.; et al. The vulnerability of Indo-Pacific mangrove forests to sea-level rise. Nature 2015, 526, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Nagelkerken, I.; Blaber, S.; Bouillon, S.; Green, P.; Haywood, M.; Kirton, L.; Meynecke, J.O.; Pawlik, J.; Penrose, H.; Sasekumar, A. The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat. Bot. 2008, 89, 155–185. [Google Scholar] [CrossRef] [Green Version]
- Tamin, N.M.; Zakaria, R.; Hashim, R.; Yin, Y. Establishment of Avicennia marina mangroves on accreting coastline at Sungai Haji Dorani, Selangor, Malaysia. Estuar. Coast. Shelf Sci. 2011, 94, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.D.; Lin, Y.M.; Liao, M.M.; Zhou, H.C.; Li, Y.Y. Characterization and antioxidative properties of condensed tannins from the mangrove plant Aegiceras corniculatum. J. Appl. Polym. Sci. 2012, 124, 2463–2472. [Google Scholar] [CrossRef]
- Chen, S.; Wang, Q.; Lu, H.; Li, J.; Yang, D.; Liu, J.; Yan, C. Phenolic metabolism and related heavy metal tolerance mechanism in Kandelia Obovata under Cd and Zn stress. Ecotoxicol. Environ. Saf. 2019, 169, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Xiao, H.; Sun, D.; Duan, S.S. Investigation of the inhibitory effects of mangrove leaves and analysis of their active components on Phaeocystis globosa during different stages of leaf age. Int. J. Environ. Res. Public Health 2018, 15, 2434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.W.; Tian, F.; Duan, L.Y.; An, M.; Duan, S.S. Allelopathic effects of mangrove plant Bruguiera gymnorrhiza on microalgae. Allelopath. J. 2012, 30, 291–298. [Google Scholar]
- Salminen, J.P. Two-dimensional tannin fingerprints by liquid chromatography tandem mass spectrometry offer a new dimension to plant tannin analyses and help to visualize the tannin diversity in plants. J. Agric. Food Chem. 2018, 66, 9162–9171. [Google Scholar] [CrossRef]
- Chou, C.H.; Leu, L.L. Allelopathic substances and interactions of Delonix regia (BOJ) RAF. J. Chem. Ecol. 1992, 18, 2285–2303. [Google Scholar] [CrossRef]
- Rawat, M.S.M.; Pant, G.; Prasad, D.T.; Joshi, R.; Pande, C.B. Plant growth inhibitors (Proanthocyanidins) from Prunus armeniaca. Biochem. Syst. Ecol. 1998, 26, 13–23. [Google Scholar] [CrossRef]
- Batista, E.F.; Costa, D.M.; Guilhon, G.M.; Muller, A.H.; Santos, L.S.; Arruda, M.S.P.; Arruda, A.C.; Silva, M.N.; Silva, G.K.R.; Secco, R.; et al. Chemical constituents and allelopathic and antioxidant activities of Alchorneopsis floribunda Müll. Arg. (Euphorbiaceae). Nat. Prod. Res. 2011, 27, 1–8. [Google Scholar] [CrossRef]
- Lang, T.; Sun, H.M.; Li, N.Y.; Lu, Y.J.; Shen, Z.D.; Jing, X.S.; Xiang, M.; Shen, X.; Chen, S.L. Multiple signaling networks of extracellular ATP, hydrogen peroxide, calcium, and nitric oxide in the mediation of root ion fluxes in secretor and non-secretor mangroves under salt stress. Aquat. Bot. 2014, 119, 33–43. [Google Scholar] [CrossRef]
- Zhang, L.L.; Lin, Y.M.; Zhou, H.C.; Wei, S.D.; Chen, J.H. Condensed tannins from mangrove species Kandelia candel and Rhizophora mangle and their antioxidant activity. Molecules 2010, 15, 420–431. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.C.; Tam, N.F.Y.; Lin, Y.M.; Wei, S.D.; Li, Y.Y. Changes of condensed tannins during decomposition of leaves of Kandelia obovata in a subtropical mangrove swamp in China. Soil Biol. Biochem. 2012, 44, 113–121. [Google Scholar] [CrossRef]
- Harris, R.W. Root shoot ratios. J. Arboricult. 1992, 18, 39–42. [Google Scholar]
- Latif, S.; Chiapusio, G.; Weston, L.A. Allelopathy and the role of allelochemicals in plant defence. Adv. Bot. Res. 2017, 82, 19–54. [Google Scholar]
- Zhang, Z.; Liu, Y.; Yuan, L.; Weber, E.; van Kleunen, M. Effect of allelopathy on plant performance: A meta-analysis. Ecol Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar] [CrossRef]
- Castells, E. Indirect Effects of Phenolics on Plant Performance by Altering Nitrogen Cycling: Another Mechanism of Plant–Plant Negative Interactions. In Allelopathy in Sustainable Agriculture and Forestry; Zeng, R.S., Mallik, A.U., Luo, S.M., Eds.; Springer: New York, NY, USA, 2008; pp. 137–156. [Google Scholar]
- Huang, Y.J.; Ge, Y.Y.; Wang, Q.L.; Zhou, H.; Liu, W.X.; Christie, P. Allelopathic Effects of Aqueous Extracts of Alternanthera philoxeroides on the Growth of Zoysia matrella. Pol. J. Environ. Stud. 2017, 26, 97–105. [Google Scholar] [CrossRef]
- Zhang, K.M.; Shen, Y.; Fang, Y.M.; Liu, Y. Changes in gametophyte physiology of Pteris multifida induced by the leaf leachate treatment of the invasive Bidens pilosa. Environ. Sci. Pollut. R. 2016, 23, 3578–3585. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy of Lantana camara as an invasive plant. Plants 2021, 10, 1028. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.D.; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [Green Version]
- Lalremsang, P.; Gopichand, B.; Upadhyaya, K.; Remlalpeka, C.; Lungmuana, S.; Singh, B.P. Allelopathic effects of Flemingia semialata Roxb. on seedling growth of maize (Zea mays L.) and rice (Oryza sativa L.). Allelopath. J. 2020, 50, 173–183. [Google Scholar] [CrossRef]
- Farooq, M.; Jabran, K.; Cheema, Z.A.; Wahid, A.; Siddique, K.H. The role of allelopathy in agricultural pest management. Pest Manag. Sci. 2011, 67, 493–506. [Google Scholar] [CrossRef]
- Sunulahpašić, A.; Čekić, S.; Golijan, J.; Hamidović, S. The ecological role of interactions between plants in agroecosystems. Agro-Knowl. J. 2017, 18, 293–305. [Google Scholar] [CrossRef]
- Macías, F.A.; Mejías, F.J.R.; Molinillo, J.M.G. Recent advances in allelopathy for weed control: From knowledge to applications. Pest Manag. Sci. 2019, 75, 2413–2436. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Feagin, R.A.; Innocenti, R.A.; Hu, B.; He, M.; Li, H. Invasion and ecological effects of exotic smooth cordgrass Spartina alterniflora in China. Ecol. Eng. 2020, 143, 105670. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Indices (A. corniculatum) | Leaf Litter Leachates (LLLs) | Purified Condensed Tannins (PCTs) | ||||||
|---|---|---|---|---|---|---|---|---|
| Low (L) | Medium (M) | High (H) | Very High (VH) | Low (L) | Medium (M) | High (H) | Very High (VH) | |
| Final germination of roots | 0 | 0 | 0 | −0.67 | 0 | 0 | −0.11 | −0.56 |
| Final germination of stems | 0 | 0 | −0.88 | −1.00 | −0.14 | −0.14 | −0.29 | −1.00 |
| Number of fine roots | −0.02 | −0.12 | −0.86 | −1.00 | −0.04 | −0.36 | −0.46 | −0.92 |
| Number of nutritive roots | −0.34 | −0.43 | −0.41 | −0.95 | 0.13 | 0.20 | −0.13 | −0.21 |
| Number of leaves | 0.22 | −0.33 | −0.14 | – | −0.25 | −0.75 | −0.75 | – |
| Length of fine roots | 0.48 | 0.09 | −0.68 | −1.00 | 0.05 | −0.23 | −0.47 | −0.69 |
| Length of nutritive roots | 0.47 | 0.24 | −0.50 | −0.88 | 0.17 | −0.15 | −0.23 | −0.55 |
| Length of stems | −0.02 | −0.49 | – | – | −0.40 | 0.26 | −0.18 | – |
| Biomass of roots | 0.59 | 0.16 | −0.76 | −1.00 | −0.02 | −0.51 | −0.64 | −0.71 |
| Biomass of stems | 0.13 | −0.30 | −1.00 | −1.00 | −0.38 | −0.19 | −0.53 | – |
| Biomass of leaves | 0.53 | −0.28 | −0.99 | −1.00 | 0.19 | −0.60 | −0.81 | – |
| Physiological Indices (A. corniculatum) | Concentration Levels | ||||
|---|---|---|---|---|---|
| Control (CK) | Low (L) | Medium (M) | High (H) | Very High (VH) | |
| Final germination of roots | NSD | NSD | NSD | NSD | NSD |
| Final germination of stems | NSD | NSD | NSD | NSD | – |
| Initiation time of roots | NSD | NSD | NSD | * | NSD |
| Initiation time of stems | NSD | NSD | NSD | * | – |
| Number of fine roots | NSD | NSD | NSD | ** | NSD |
| Number of nutritive roots | NSD | NSD | * | NSD | *** |
| Number of leaves | NSD | NSD | NSD | NSD | – |
| Length of fine roots | NSD | NSD | NSD | NSD | NSD |
| Length of nutritive roots | NSD | NSD | NSD | NSD | NSD |
| Length of stems | NSD | NSD | NSD | NSD | – |
| Biomass of roots | NSD | NSD | NSD | ** | ** |
| Biomass of stems | NSD | NSD | NSD | NSD | – |
| Biomass of leaves | NSD | NSD | NSD | NSD | NSD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, T.; Wei, P.; Chen, X.; Fu, Y.; Tam, N.F.-y.; Hu, Z.; Chen, Z.; Li, F.; Zhou, H. Microcosm Study on Allelopathic Effects of Leaf Litter Leachates and Purified Condensed Tannins from Kandelia obovata on Germination and Growth of Aegiceras corniculatum. Forests 2021, 12, 1000. https://doi.org/10.3390/f12081000
Lang T, Wei P, Chen X, Fu Y, Tam NF-y, Hu Z, Chen Z, Li F, Zhou H. Microcosm Study on Allelopathic Effects of Leaf Litter Leachates and Purified Condensed Tannins from Kandelia obovata on Germination and Growth of Aegiceras corniculatum. Forests. 2021; 12(8):1000. https://doi.org/10.3390/f12081000
Chicago/Turabian StyleLang, Tao, Pingping Wei, Xiaoxia Chen, Yijian Fu, Nora Fung-yee Tam, Zhangli Hu, Zhiteng Chen, Fenglan Li, and Haichao Zhou. 2021. "Microcosm Study on Allelopathic Effects of Leaf Litter Leachates and Purified Condensed Tannins from Kandelia obovata on Germination and Growth of Aegiceras corniculatum" Forests 12, no. 8: 1000. https://doi.org/10.3390/f12081000