
Population Demographic History of a Rare and Endangered Tree Magnolia sprengeri Pamp. in East Asia Revealed by Molecular Data and Ecological Niche Analysis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Investigation and Samples Collection
2.2. DNA Extraction and Sequencing
2.3. Genetic Diversity and Population Structure Analyses
2.4. Species Distribution Modeling
3. Results
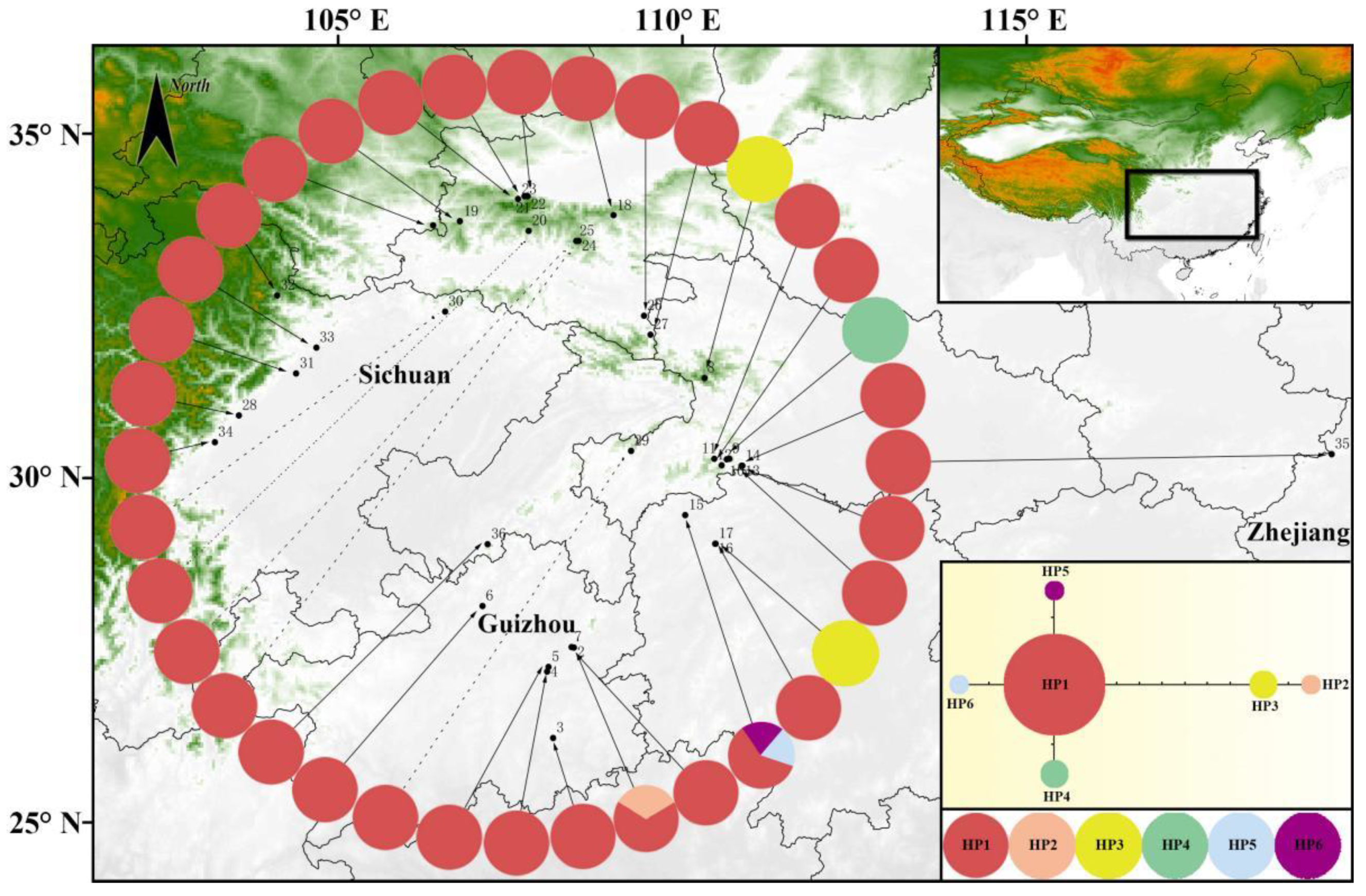
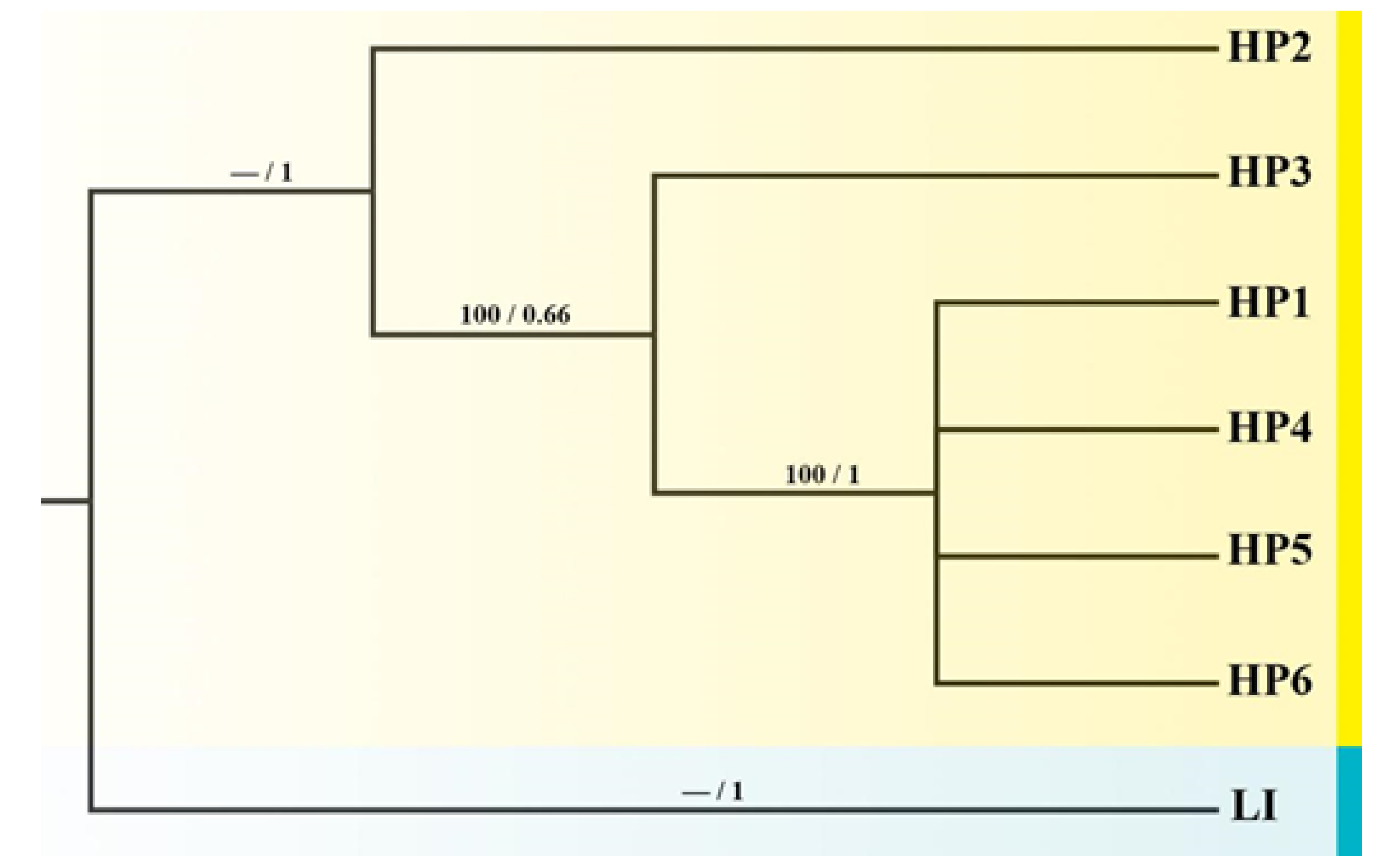
3.1. The Distribution and Relationships of Chloroplast DNA (cpDNA) Haplotypes
3.2. Genetic Differentiation and Population Structure
3.3. Species Distribution Simulation Results
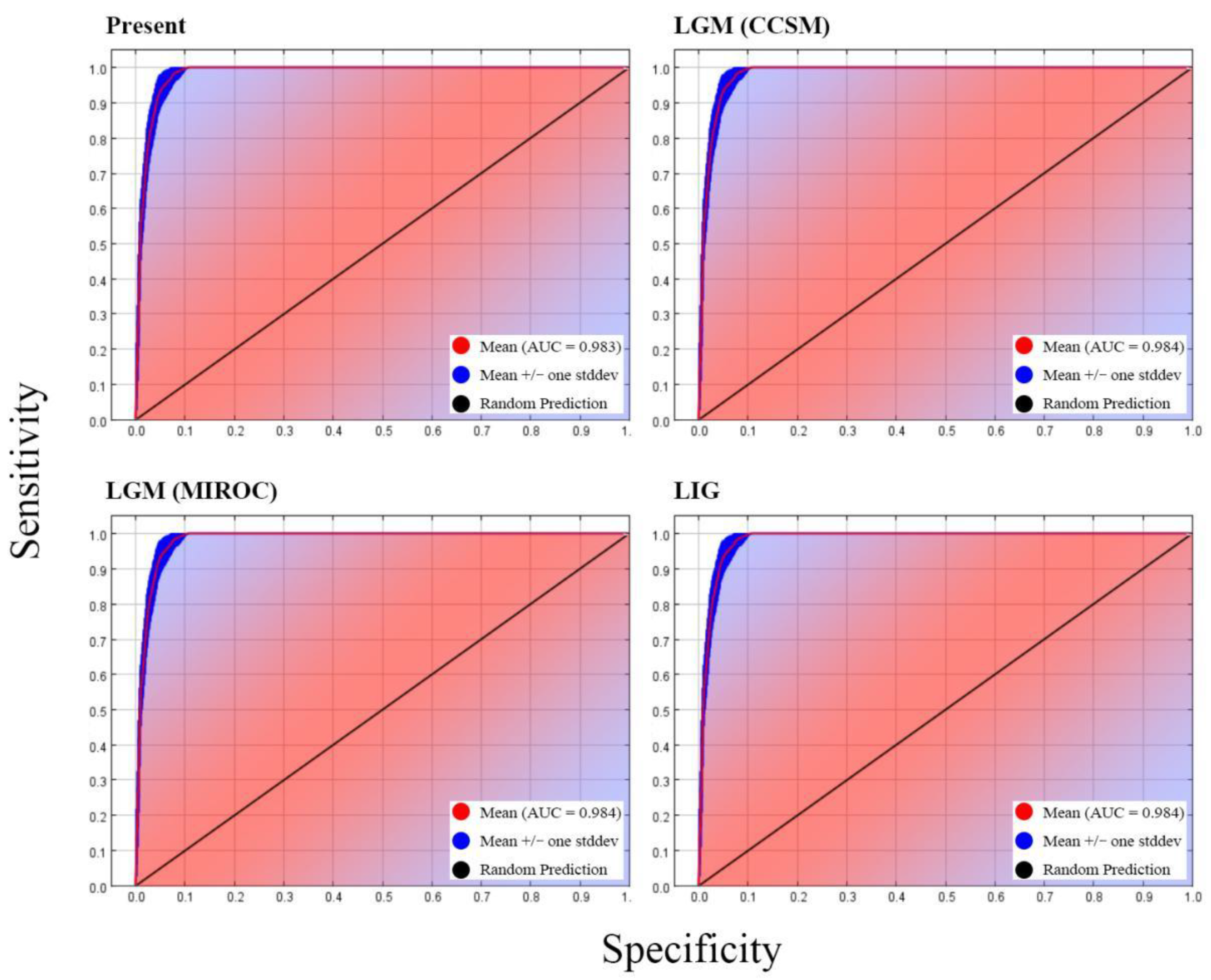
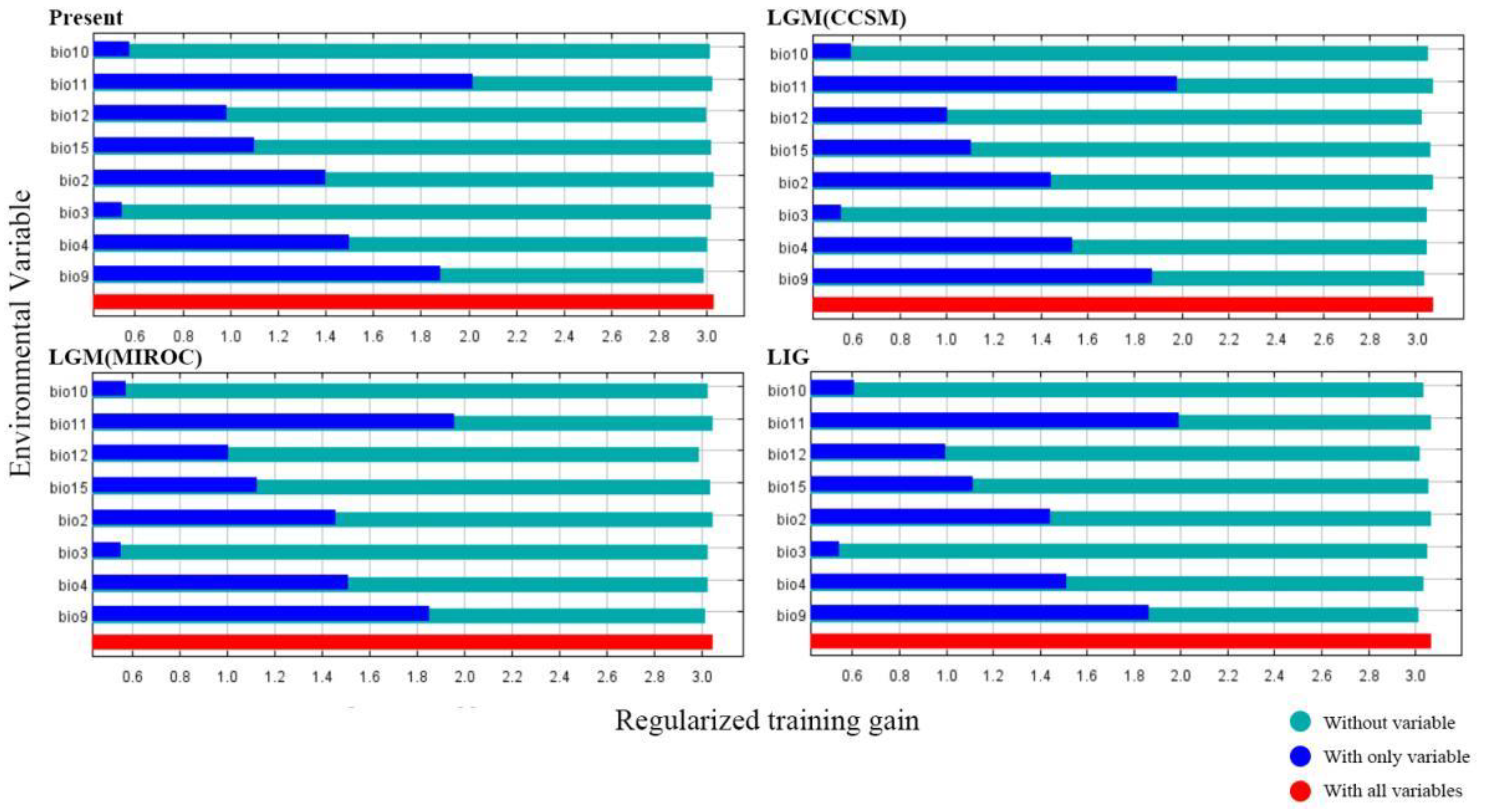
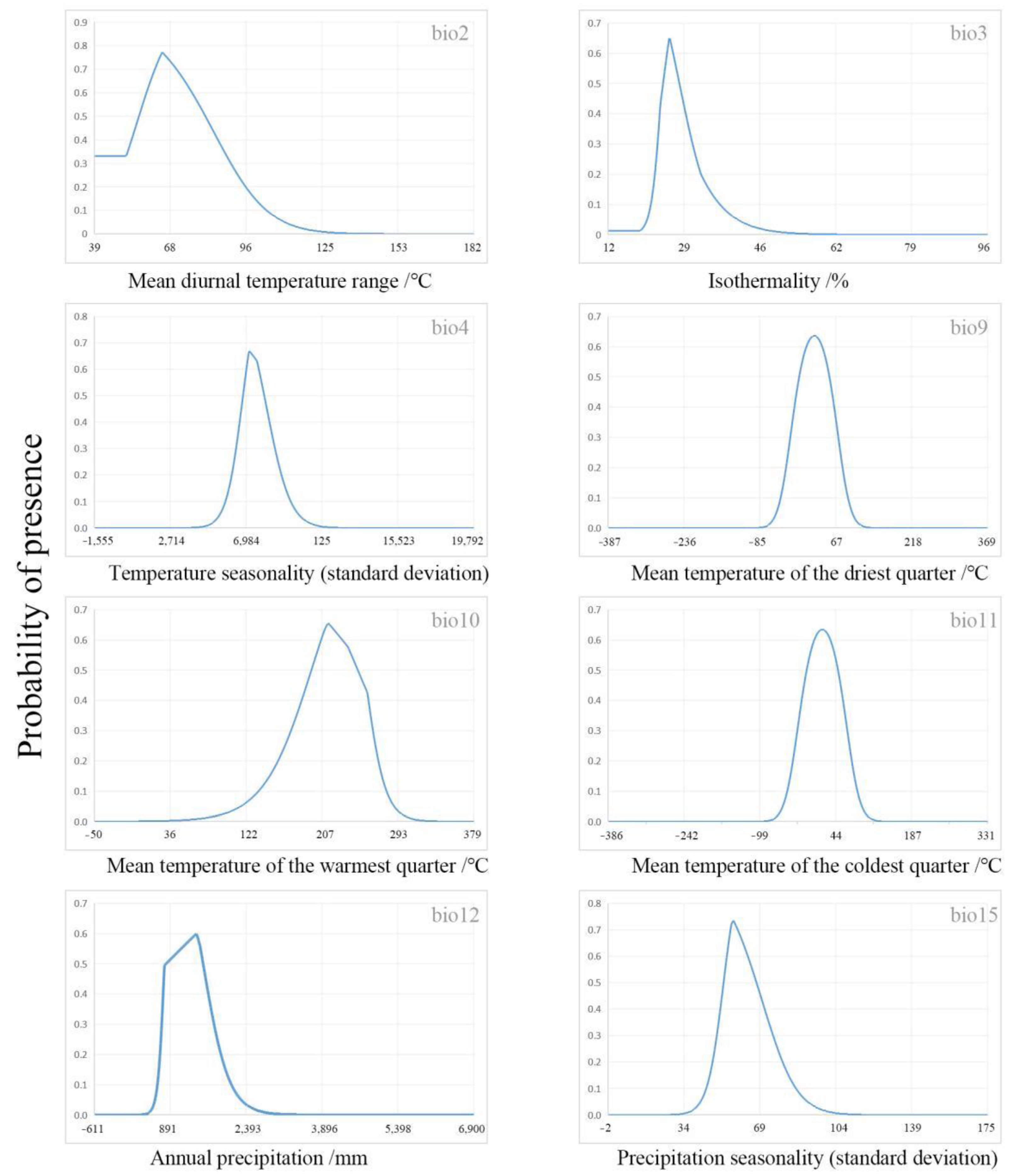
3.3.1. Model Performance and Contributions of Variables
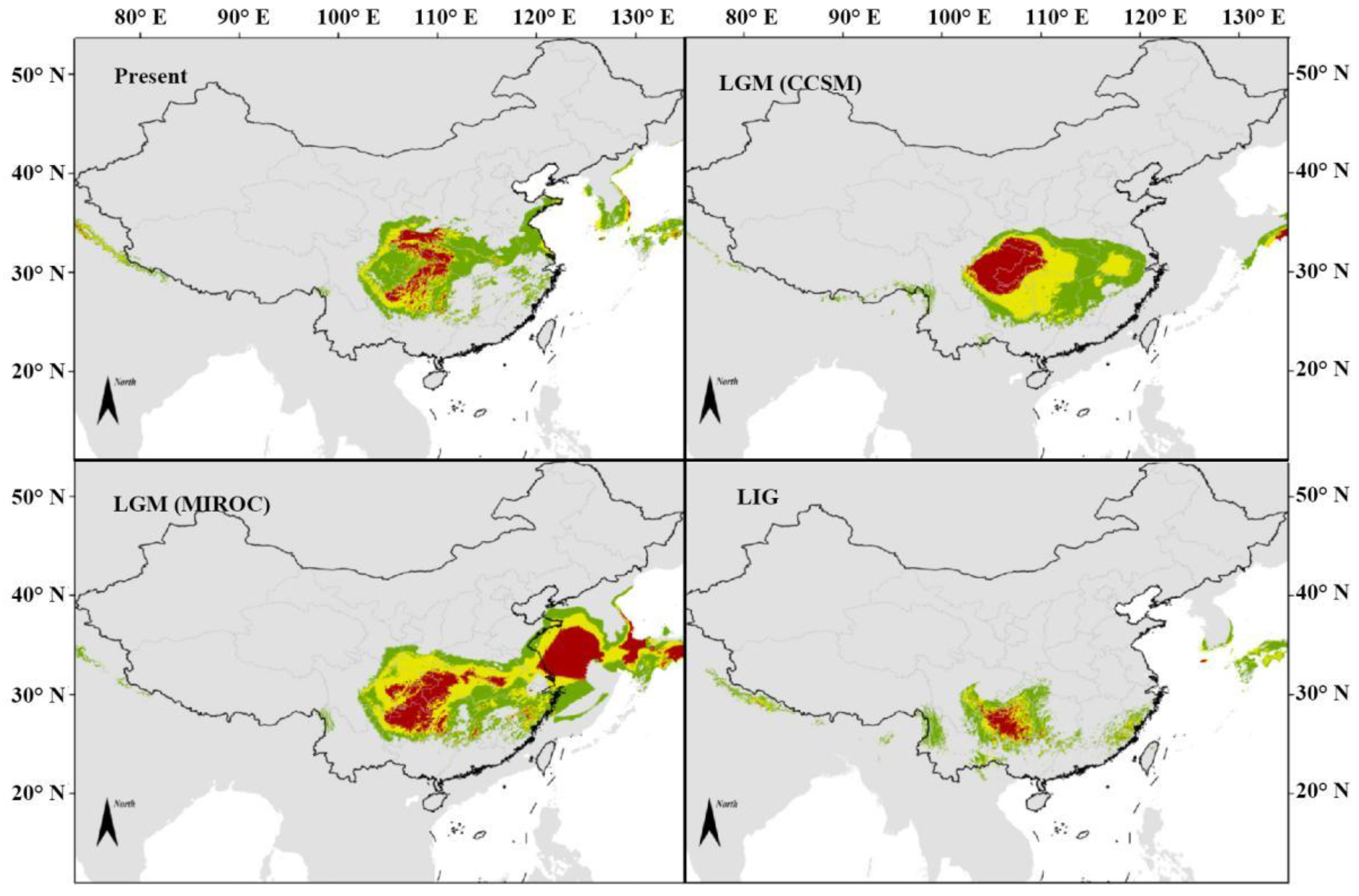
3.3.2. M. sprengeri Potential Distribution
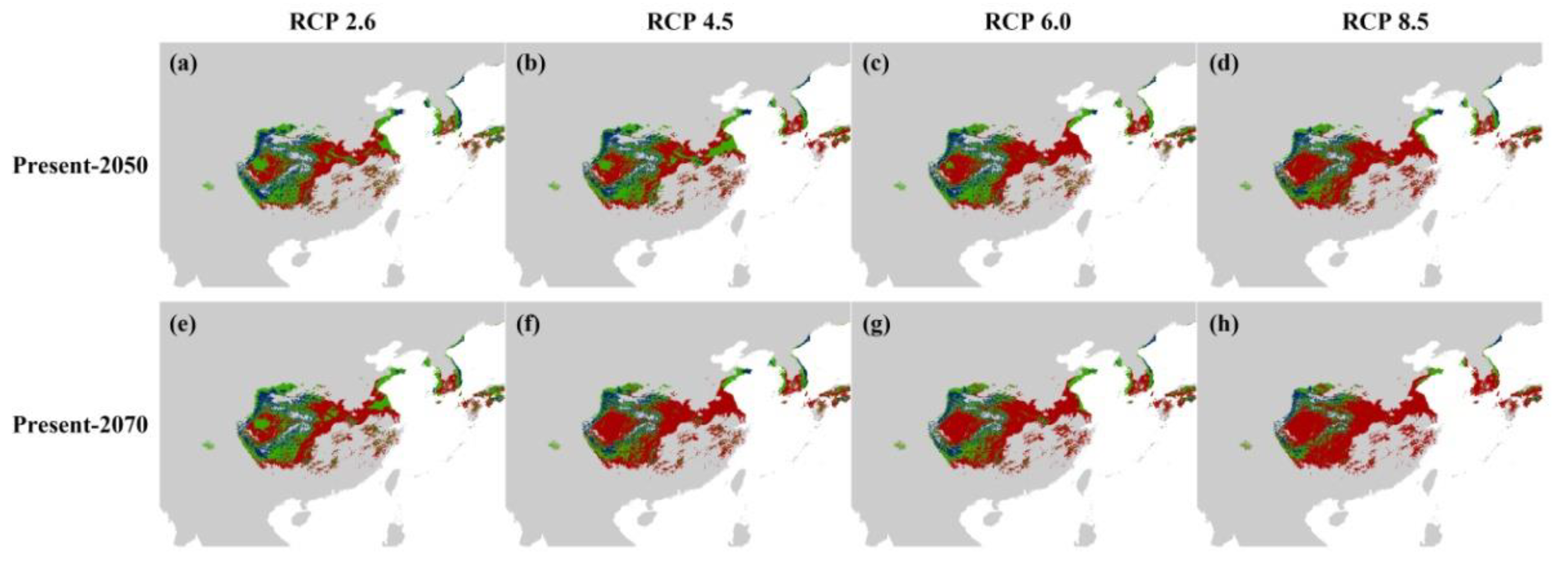
3.3.3. Suitable Distribution under Future Climate Scenarios
4. Discussion
4.1. Diversity and Genetic Structure
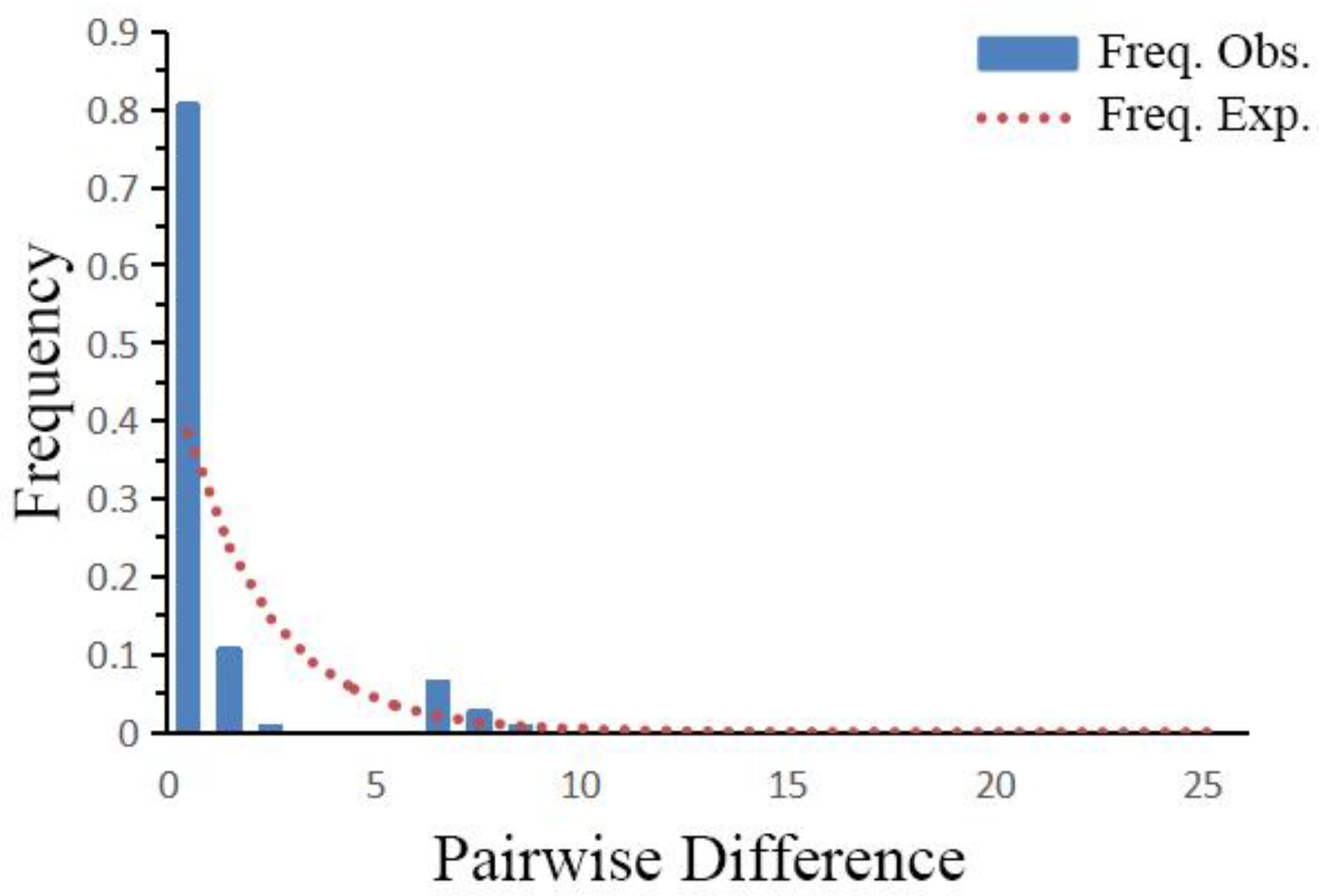
4.2. Population History and Species Distribution Fluctuations
4.3. Conservation Strategies and Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, M.B. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewitt, G.M. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc. Lond. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Suarez-Gonzalez, A.; Sutton, J.T.; Trant, A.J.; Zamlynny, E.; Good, S.V. Rethinking refugia: Tree topology, divergence dates, and demographic history trace the distribution of the endangered Plymouth gentian (Sabatia kennedyana) from the Pleistocene glaciation to present day. Am. J. Bot. 2015, 102, 609–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petit, R.J.; Kremer, A.; Wagner, D.B. Geographic structure of chloroplast DNA polymorphisms in European oaks. Theor. Appl. Genet. 1993, 87, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Tzedakis, P.C.; Lawson, I.T.; Frogley, M.R.; Hewitt, G.M.; Preece, R.C. Buffered tree population changes in a Quaternary refugium: Evolutionary implications. Science 2002, 297, 2044–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Deng, M.; Jiang, X.L.; Westwood, M.; Song, Y.G.; Turkington, R. Phylogeography of Quercus glauca (Fagaceae), a dominant tree of East Asian subtropical evergreen forests, based on three chloroplast DNA interspace sequences. Tree Genet. Genomes 2015, 11, 1–17. [Google Scholar] [CrossRef]
- Michalczyk, I.M.; Opgenoorth, L.; Luecke, Y.; Huck, S.; Ziegenhagen, B. Genetic support for perglacial survival of Juniperus communis L. in Central Europe. Holocene 2010, 20, 887–894. [Google Scholar] [CrossRef]
- Sommer, R.S.; Zachos, F.E. Fossil evidence and phylogeography of temperate species: ‘glacial refugia’ and post-glacial recolonization. J. Biogeogr. 2010, 36, 2013–2020. [Google Scholar] [CrossRef]
- Petit, R.J.; Brewer, S.; Bordacs, S.; Burg, K.; Cheddadi, R.; Coart, E.; Cottrell, J.; Csaikl, U.M.; Van Dam, B.; Deans, J.D.; et al. Identification of refugia and post-glacial colonisation routes of European white oaks based on chloroplast DNA and fossil pollen evidence. For. Ecol. Manag. 2002, 156, 49–74. [Google Scholar] [CrossRef]
- Kitamura, A. Quaternary climatic changes and species diversity: Responses of marine organisms to glacial-interglacial cycles. Fossils 2004, 75, 54–59. [Google Scholar]
- Becquet, V.; Simon-Bouhet, B.; Pante, E.; Hummel, H.; Garcia, P. Glacial refugium versus range limit: Conservation genetics of Macoma balthica, a key species in the Bay of Biscay (France). J. Exp. Mar. Biol. Ecol. 2012, 432, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Carstens, B.C.; Richards, C.L. Integrating coalescent and ecological niche modeling in comparative phylogeography. Evolution 2007, 61, 1439–1454. [Google Scholar] [CrossRef]
- Su, Z.M. The subtropical mid-montane coniferous and broad-leaf mixed forest in Guangxi. Guihaia 1983, 3, 33–42. [Google Scholar]
- Liu, H.Y.; Jin, X.L.; Xue, H.W.; Luo, X.M.; Zhang, R.Q. Research Progress on Community Characteristics and Population Ecology of Davidia involucrate Baill. Chin. Agric. Sci. Bull. 2012, 28, 1–4. [Google Scholar]
- Wan, Q.; Zheng, Z.; Huang, K.; Guichoux, E.; Petit, R.J. Genetic divergence within the monotypic tree genus Platycarya (Juglandaceae) and its implications for species’ past dynamics in subtropical china. Tree Genet. Genomes 2017, 13, 73. [Google Scholar] [CrossRef]
- Shi, Y.; Cui, Z.; Su, Z. The Quaternary Glaciations and Environmental Variations in China, 1st ed.; Hebei Science and Technology Press: Shijiazhuang, China, 2006; pp. 65–101. [Google Scholar]
- Wang, J. Refugia within refugia: The case study of a canopy tree (Eurycorymbus cavaleriei) in subtropical China. J. Biogeogr. 2010, 36, 2156–2164. [Google Scholar] [CrossRef]
- Chen, C.; Lu, R.S.; Zhu, S.S.; Tamaki, I.; Qiu, Y.X. Population structure and historical demography of Dipteronia dyeriana (Sapindaceae), an extremely narrow palaeoendemic plant from China: Implications for conservation in a biodiversity hot spot. Heredity 2017, 119, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, S.; Lei, S.Q.; Hu, W.; Deng, L.L.; Li, B.; Meng, Q.L.; Soltis, D.E.; Soltis, P.S.; Fan, D.M.; Zhang, Z.Y. Repeated range expansions and inter-/postglacial recolonization routes of Sargentodoxa cuneata (Oliv.) Rehd. et Wils. (Lardizabalaceae) in subtropical China revealed by chloroplast phylogeography. Mol. Phylogenet. Evol. 2015, 85, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.G.; Chen, J.; Zhu, H.; Li, Y.F.; Li, X.X.; Li, M.; Duan, Y.F.; Chen, L.; Wang, X.R. Phylogeography and the population genetic structure of flowering cherry Cerasus serrulata (Rosaceae) in subtropical and temperate China. Ecol. Evol. 2020, 10, 11262–11276. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, X.; Kang, H.; Sun, X.; Yin, S.; Du, H.; Yamanaka, N.; Gapare, W.; Wu, H.X.; Liu, C. Phylogeography of Quercus variabilis based on chloroplast DNA sequence in East Asia: Multiple glacial refugia and Mainland-migrated island populations. PLoS ONE 2012, 7, e47268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ming, L.; Wang, Q.; Wu, Z.J.; López-Pujol, J.; Li, D.Z.; Zhang, Z.Y. Molecular phylogeography of Fagus engleriana (Fagaceae) in subtropical China: Limited admixture among multiple refugia. Tree Genet. Genomes 2012, 8, 1203–1212. [Google Scholar]
- Fu, D.L.; Xiong, B.H.; Chen, X.; Tian, W.X.; Du, L. Analyses of chloroplast genomic and morphological evolutionomy of Yulania sprengeri and two new confusable species (Magnoliaceae). J. Agric. For. 2019, 7, 212. [Google Scholar] [CrossRef]
- China Flora Editorial Board. Flora of China, 3rd ed.; Beijing Science Press: Beijing, China, 2008; Volume 7, p. 73. [Google Scholar]
- Song, C.; Liu, H. Habitat differentiation and conservation gap of Magnolia biondii, M. denudata, and M. sprengeri in China. PeerJ 2019, 6, e6126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.X.; Ejder, E. Magnolia sprengeri Pamp.: Morphological variation and geographical distribution. Plant Biosyst. 2011, 145, 906–923. [Google Scholar] [CrossRef]
- Peng, X.; Chen, K. Research on Popularization and Application of Si Chuan Native Tree Magnolia sprengeri Pamp. in Landscape Architecture. Archit. Cult. 2018, 5, 114–116. [Google Scholar]
- Yang, D.L. Use of Magnolia (Magnolia grandiflora) Seeds in Medicine, and Possible Mechanisms of Action. Nuts Seeds Health Dis. Prev. 2011, 86, 727–732. [Google Scholar]
- Rivers, M.C.; Beech, E.; Murphy, L.; Oldfield, S. The Red List of Magnoliaceae—Revised and Extended, 1st ed.; Botanic Gardens Conservation International: Surrey, UK, 2016; p. 48. [Google Scholar]
- Ho, V.T.; Tran, T.K.P.; Vu, T.T.T.; Widiarsih, S. Comparison of matK and rbcL DNA barcodes for genetic classification of jewel orchid accessions in Vietnam. J. Genet. Eng. Biotechnol. 2021, 19, 93. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [Green Version]
- Massoni, J.; Forest, F.; Sauquet, H. Increased sampling of both genes and taxa improves resolution of phylogenetic relationships within Magnoliidae, a large and early-diverging clade of angiosperms. Mol. Phylogenet. Evol. 2014, 70, 84–93. [Google Scholar] [CrossRef]
- Azuma, H.; García-Franco, J.G.; Rico-Gray, V.; Thien, L.B. Molecular phylogeny of the Magnoliaceae: The biogeography of tropical and temperate disjunctions. Am. J. Bot. 2001, 88, 2275–2285. [Google Scholar] [CrossRef]
- Sangtae, K.; Youngbae, S. Phylogeny of Magnoliaceae Based on Ten Chloroplast DNA Regions. J. Plant Bio. 2013, 56, 290–305. [Google Scholar]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef]
- Jiang, C.; Cao, L.; Yuan, Y.; Chen, M.; Jin, Y.; Huang, L. Barcoding melting curve analysis for rapid, sensitive, and discriminating authentication of saffron (Crocus sativus L.) from its adulterants. Biomed Res. Int. 2014, 2014, 809037. [Google Scholar] [CrossRef] [Green Version]
- Swift, J.F.; Lance, R.F.; Guan, X.; Britzke, E.R.; Lindsay, D.L.; Edwards, C.E. Multifaceted DNA metabarcoding: Validation of a noninvasive, next-generation approach to studying bat populations. Evol. Appl. 2018, 11, 1120–1138. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.A.; Simpson, B.B. Breeding system evolution in Tarasa (Malvaceae) and selection for reduced pollen grain size in the polyploid species. Am. J. Bot. 2004, 91, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Erickson, D.L. A two-locus global DNA barcode for land plants: The coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS ONE 2007, 2, e508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Res. 1999, 41, 95–98. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Zheng, Q.J.; Qian, Z.Q.; Yang, J.; Zhang, Y.P.; Li, Z.H.; Zhao, G.F. Genetic Structure and Evolutionary History of Three Alpine Sclerophyllous Oaks in East Himalaya-Hengduan Mountains and Adjacent Regions. Front. Plant Sci. 2016, 7, 1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2007, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research--an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Cockerham, C.C. Drift and mutation with a finite number of allelic states. Proc. Natl. Acad. Sci. USA 1984, 81, 530–534. [Google Scholar] [CrossRef] [Green Version]
- Pons, O.; Petit, R.J. Measuring and testing genetic differentiation with ordered versus unordered alleles. Genetics 1996, 144, 1237–1245. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Sun, L.; Jiang, Z.; Wan, X.; Zou, X.; Yao, X.; Wang, Y.; Yin, Z. The complete chloroplast genome of Magnolia polytepala: Comparative analyses offer implication for genetics and phylogeny of Yulania. Gene 2020, 736, 144410. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FigTree v1.3.1. 2006–2009. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 29 November 2012).
- Otto-Bliesner, B.L.; Marshall, S.J.; Overpeck, J.T.; Miller, G.H.; Hu, A. Simulating Arctic climate warmth and icefield retreat in the last interglaciation. Science 2006, 311, 1751–1753. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.P.; Wang, G.Y. Key findings and assessment results of ipcc wgi fifth assessment report. J. Glaciol. Geocryol. 2013, 5, 10–18. [Google Scholar]
- Steven, J.P.; Robert, P.A.; Robert, E.S. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar]
- Worth, J.; Williamson, G.J.; Sakaguchi, S.; Nevill, P.G.; Jordan, G.J. Environmental niche modeling fails to predict Last Glacial Maximum refugia: Niche shifts, microrefugia or incorrect palaeoclimate estimates? Glob. Ecol. Biogeogr. 2015, 23, 1186–1197. [Google Scholar] [CrossRef]
- Cao, B.; Bai, C.; Zhang, L.; Li, G.; Mao, M. Modeling habitat distribution of Cornus officinalis with MaxEnt modeling and fuzzy logics in China. J. Plant Ecol. 2016, 9, 742–751. [Google Scholar] [CrossRef] [Green Version]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Shah, T.; Hao, Z.; Taba, S.; Zhang, S.; Gao, S.; Liu, J.; Cao, M.; Wang, J.; Prakash, A.B.; et al. Comparative SNP and haplotype analysis reveals a higher genetic diversity and rapider LD decay in tropical than temperate germplasm in maize. PLoS ONE 2011, 6, e24861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.J.; Chiang, Y.C.; Chou, C.H.; Chiang, T.Y. Nested clade analysis of Dunnia sinensis (Rubiaceae), a monotypic genus from China based on organelle DNA sequences. Conservat. Genet. 2002, 3, 351–362. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M. Effects of Life History Traits on Genetic Diversity in Plant Species. Philos. Trans. Biol. Sci. 1996, 351, 1291–1298. [Google Scholar]
- Shu, X.; Yang, Z.; Duan, H.; Yang, X.; Yu, H. Seed germinating characteristics of endangered plant Magnolia officinalis. China J. Chin. Mater. Med. 2010, 35, 419–422. [Google Scholar]
- Lopez-Pujol, J.L.; Zhang, F.M.; Sun, H.Q.; Ying, T.S.; Ge, S. Mountains of Southern China as “Plant Museums” and “Plant Cradles”: Evolutionary and Conservation Insights. Mt. Res. Dev. 2011, 31, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Rost, K.T. Pleistocene paleoenvironmental changes in the high mountain ranges of central China and adjacent regions. Quat. Int. 2000, 65, 147–160. [Google Scholar] [CrossRef]
- Petrosian, V.G.; Tokarskaia, O.N.; Kashentseva, T.A.; Korochkin, L.I.; Ryskov, A.P. Assessment of population differentiation using DNA fingerprinting and modified Wright’s FST-statistics. Genetika 2003, 39, 229–235. [Google Scholar]
- Sork, V.L.; Nason, J.; Campbell, D.R.; Fernandez, J.F. Landscape approaches to historical and contemporary gene flow in plants. Trends Ecol. Evol. 1999, 14, 219–224. [Google Scholar] [CrossRef]
- Corral-Aguirre, J.; Sánchez-Velásquez, L.R. Seed ecology and germination treatments in Magnolia dealbata: An endangered species. Flora 2006, 201, 227–232. [Google Scholar] [CrossRef]
- Tian, K.; Zhang, G.X.; Cheng, X.F.; He, S.J.; Yang, Y.X. The habitat fragility of Manglietiastrum sinicum. Acta Bot. Yunnanica 2003, 25, 551–556. [Google Scholar]
- Vovides, A.P.; Iglesias, C.G. Seed Germination of Magnolia dealbata Zucc. (Magnoliaceae), an endangered species from Mexico. HortScience 1996, 31, 877. [Google Scholar] [CrossRef] [Green Version]
- He, H.L.; Cai, Y.L.; Gao, T.X.; Li, B.; Wan, T.; Wang, Y. SSR evaluation of genetic diversity of seven wild Prunus tomentosa populations in Shaanxi. J. Northwest A F Univ. 2015, 43, 193–198. [Google Scholar]
- Sur, D.K.; Plesa, M.L. Treatment of Allergic Rhinitis. Asian Pac. J. Allergy Immunol. 2015, 92, 985–992. [Google Scholar]
- Lacy, R.C. Loss of genetic diversity from managed population: Interacting effects of drift, mutation, immigration, Selection and population subdivision. Conserv. Biol. 1987, 1, 143–158. [Google Scholar] [CrossRef]
- Qiu, Y.X.; Fu, C.X.; Comes, H.P. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenet. Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef]
- Wang, D.Q.; Zhang, Y.C. Diurnal variations of precipitation and circulation simulated by model for interdisciplinary research on climate. J. Nanjing Univ. 2009, 45, 724–733. [Google Scholar]
- Bhagwat, S.A.; Willis, K.J. Species persistence in northerly glacial refugia of Europe: A matter of chance or biogeographical traits? J. Biogeogr. 2008, 35, 464–482. [Google Scholar] [CrossRef]
- Mackay, A. Climate change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. J. Environ. Qual. 2007, 36, 2407. [Google Scholar] [CrossRef]
- Olsen, S.L.; Töpper, J.P.; Skarpaas, O.; Vandvik, V.; Klanderud, K. From facilitation to competition: Temperature-driven shift in dominant plant interactions affects population dynamics in seminatural grasslands. Glob. Chang. Biol. 2016, 22, 1915–1926. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Sun, L.; Tao, J. Impact of climate change on the distribution of Euscaphis japonica (Staphyleaceae) trees. Forests 2020, 11, 525. [Google Scholar] [CrossRef]
- Dulias, R. Drift Sand Fields as a Result of Past and Curent Deforestation in the Silesian-Cracow Upland, Poland. Land Degrad. Dev. 2018, 29, 1530–1539. [Google Scholar] [CrossRef]
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.; De Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Fay, M.F. Conservation of rare and endangered plants using in vitro methods. In Vitro Cell. Dev. Biol. Plant 1992, 28, 1–4. [Google Scholar] [CrossRef]
- Liu, Y.H.; Zhou, R.Z.; Zeng, Q.W. Ex situ conservation of Magnoliaceae including its rare and endangered species. J. Trop. Subtrop. Bot. 1997, 5, 1–12. [Google Scholar]
- Fang, H.; Guo, Q.; Su, W.; Deng, F.; Wang, K.Q. RP—HPLC Determination of Magnolin in Chinese Medicine Xinyi. Chin. J. Pharm. Anal. 2002, 22, 342–345. [Google Scholar]
- Shi, S.G.; Li, S.J.; Kang, Y.X.; Liu, J.J. Molecular characterization and expression analyses of an anthocyanin synthase gene from Magnolia sprengeri Pamp. Appl. Biochem. Biotechnol. 2015, 175, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Shi, S.G.; Liu, W.; Zhang, M.; Gou, L.; Kang, Y.X.; Liu, J.J. Phenotypic variation and diversity of Magnolia sprengeri Pamp. In native habitat. Genet. Mol. Res. 2015, 14, 6495–6508. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, J.R.; Quinn, R.M.; Lawton, J.H.; Eversham, B.C.; Gibbons, D.W. Rare species, the coincidence of diversity hotspots and conservation strategies. Nature 1993, 365, 335–337. [Google Scholar] [CrossRef]
- Chen, T.; Chen, Q.; Luo, Y.; Huang, Z.L.; Zhang, J.; Tang, H.R.; Pan, D.M.; Wang, X.R. Phylogeography of Chinese cherry (Prunus pseudocerasus Lindl.) inferred from chloroplast and nuclear DNA: Insights into evolutionary patterns and demographic history. Plant biol. 2015, 17, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Gerlee, P.; Lundh, T. Productivity and diversity in a cross-feeding population of artificial organisms. Evolution 2010, 64, 2716–2730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagl, W.; Habermann, T.; Fusenig, H.P. Nuclear DNA contents in four primitive angiosperms. Plant Syst. Evol. 1977, 127, 103–105. [Google Scholar] [CrossRef]
- Zhang, X.M.; Wen, J.; Dao, Z.L.; Motley, T.J.; Long, C.L. Genetic variation and conservation assessment of Chinese populations of Magnolia cathcartii (Magnoliaceae), a rare evergreen tree from the South-Central China hotspot in the Eastern Himalayas. J. Plant Res. 2010, 123, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.H.; Yang, Z.L.; Sun, B.; Liu, R.N. Genetic diversity and relationship of endangered plant Magnolia officinalis (Magnoliaceae) assessed with ISSR polymorphisms. Biochem. Syst. Ecol. 2011, 39, 71–78. [Google Scholar] [CrossRef]
- Li, J.M.; Jin, Z.X. Genetic structure of endangered Emmenopterys henryi Oliv. Based on ISSR polymorphism and implications for its conservation. Genetica 2008, 133, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Al-Turki, T.A.; Al-Namazi, A.A.; Al-Ammari, B.S.; Al-Mosallam, M.S.; Basahi, M.A. Ex-situ conservation of wheat genetic resources from Saudi Arabia. Saudi J. Biol. Sci. 2020, 27, 2318–2324. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N 1 | S 2 | H 3 | Hd 4 | π 5 | Tajima’s D | HS 6 | HT 7 | GST | NST |
|---|---|---|---|---|---|---|---|---|---|
| 87 | 10 | 6 | 0.196 | 0.0669 | −1.73 | 0.068 | 0.169 | 0.595 | 0.268 |
| Source of Variation | d.f. | Variation Components | Percentage of Variation | p |
|---|---|---|---|---|
| Among populations | 19 | 0.047 | 27.61% 1 | ≤0.005 |
| Within populations | 51 | 0.123 | 72.39% | ≤0.005 |
| Type | Variable | Code |
|---|---|---|
| climate | Mean diurnal temperature range/°C | bio2 |
| Isothermality/% | bio3 | |
| Temperature seasonality (standard deviation) | bio4 | |
| Mean temperature of the driest quarter/°C | bio9 | |
| Mean temperature of the warmest quarter/°C | bio10 | |
| Mean temperature of the coldest quarter/°C | bio11 | |
| Annual precipitation/mm | bio12 | |
| Precipitation seasonality (standard deviation) | bio15 |
| Period | Bio2 | Bio3 | Bio4 | Bio9 | Bio10 | Bio11 | Bio12 | Bio15 | |
|---|---|---|---|---|---|---|---|---|---|
| Percent contribution | LIG | 27.7 | 0.9 | 5.5 | 30.1 | 9.3 | 7 | 18.2 | 1.3 |
| LGM (CCSM) | 28.5 | 0.9 | 4.6 | 25.1 | 9.8 | 11.8 | 18.6 | 0.7 | |
| LGM (MIROC) | 30.7 | 1 | 4.1 | 29.2 | 10.2 | 5.1 | 18.5 | 1.2 | |
| Present | 26.3 | 0.5 | 4.7 | 35.2 | 10.1 | 3.3 | 18.5 | 1.3 | |
| Permutation importance | LIG | 1 | 3.7 | 34.3 | 55.3 | 2 | 0.5 | 2.4 | 0.7 |
| LGM (CCSM) | 0.2 | 10.9 | 31.4 | 49.1 | 1.9 | 1.5 | 4.2 | 0.6 | |
| LGM (MIROC) | 1.2 | 13.2 | 30.6 | 39.6 | 2.2 | 6 | 6.6 | 0.7 | |
| Present | 2.7 | 6.2 | 30.3 | 48.7 | 1.9 | 5.1 | 4.5 | 0.7 |
| Period | Area of Each Suitable Region (×104 km2) | |||
|---|---|---|---|---|
| Marginally Suitable Region | Moderately Suitable Region | Highly Suitable Region | Total Sutable Region | |
| LIG | 32.48958 | 13.72222 | 7.80208 | 54.01388 |
| LGM (CCSM) | 56.49132 | 43.99653 | 26.97917 | 127.46702 |
| LGM (MIROC) | 51.88021 | 57.42188 | 29.22743 | 138.52952 |
| Present | 71.30556 | 29.97049 | 15.03125 | 116.30730 |
| Emission | Description |
|---|---|
| RCP 1 8.5 | The radiative forcing rose to 8.5 W/m2, and the CO2 equivalent concentration reached about 1370 mL/m3 in 2100. |
| RCP6.0 | The radiative forcing stabilized at 6.0 W/m2, and the CO2 equivalent concentration stabilized at about 850 mL/m3 after 2100. |
| RCP4.5 | The radiative forcing stabilized at 4.5 W/m2, and the CO2 equivalent concentration stabilized at about 600 mL/m3 after 2100. |
| RCP2.6 | The radiative forcing reached its peak before 2100 and decreased to 2.6 W/m2 by 2100. The peak CO2 equivalent concentration was about 490 mL/m3. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, T.; Huang, X.-J.; Zhang, S.-Z.; Wang, Y.; Wang, Y.-J.; Liu, W.-Z.; Wang, Y.-L.; Zou, J.-B.; Li, Z.-H. Population Demographic History of a Rare and Endangered Tree Magnolia sprengeri Pamp. in East Asia Revealed by Molecular Data and Ecological Niche Analysis. Forests 2021, 12, 931. https://doi.org/10.3390/f12070931
Zhou T, Huang X-J, Zhang S-Z, Wang Y, Wang Y-J, Liu W-Z, Wang Y-L, Zou J-B, Li Z-H. Population Demographic History of a Rare and Endangered Tree Magnolia sprengeri Pamp. in East Asia Revealed by Molecular Data and Ecological Niche Analysis. Forests. 2021; 12(7):931. https://doi.org/10.3390/f12070931
Chicago/Turabian StyleZhou, Tong, Xiao-Juan Huang, Shou-Zhou Zhang, Yuan Wang, Ying-Juan Wang, Wen-Zhe Liu, Ya-Ling Wang, Jia-Bin Zou, and Zhong-Hu Li. 2021. "Population Demographic History of a Rare and Endangered Tree Magnolia sprengeri Pamp. in East Asia Revealed by Molecular Data and Ecological Niche Analysis" Forests 12, no. 7: 931. https://doi.org/10.3390/f12070931