
Estimation of Biomass Increase and CUE at a Young Temperate Scots Pine Stand Concerning Drought Occurrence by Combining Eddy Covariance and Biometric Methods

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
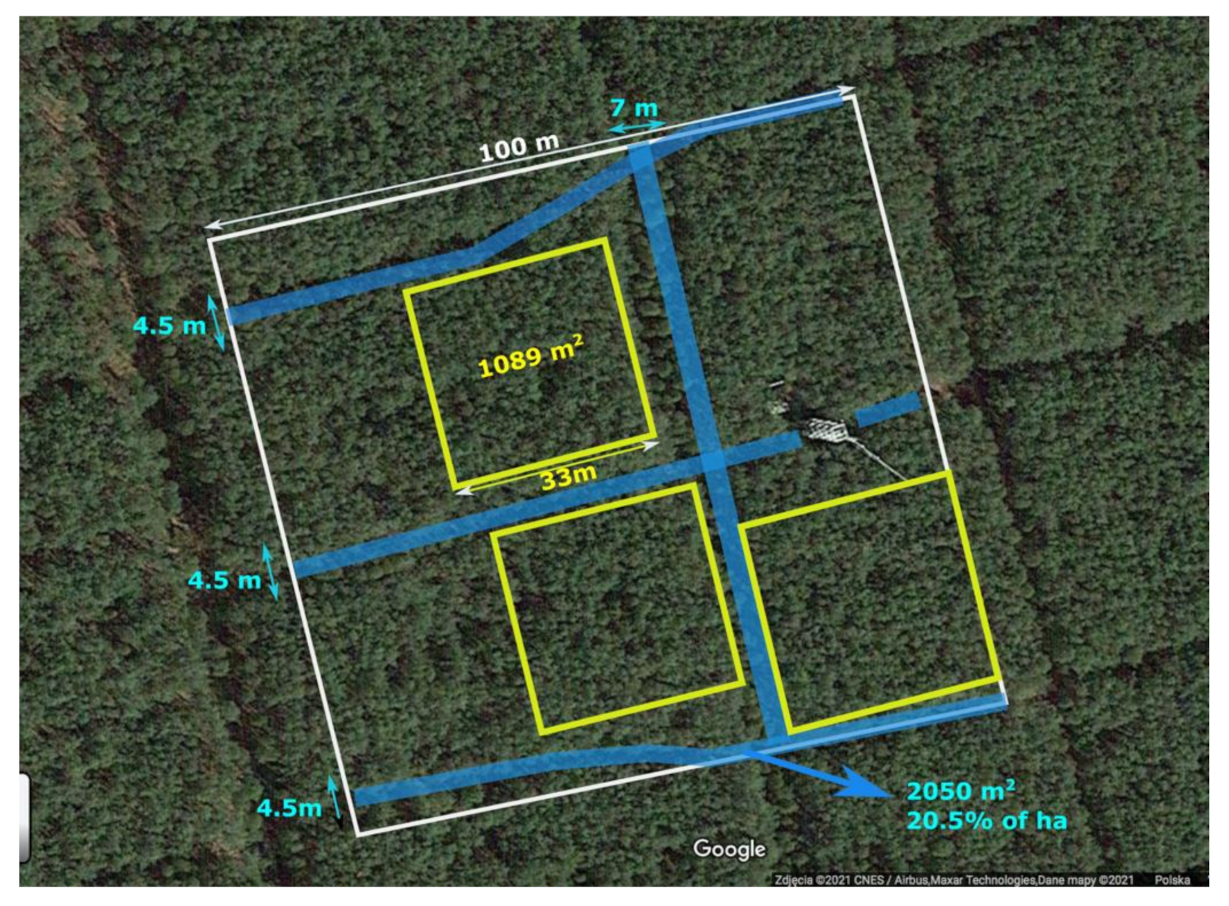
2.1. Site Description
2.2. Eddy Covariance and Meteorological Measurements
2.3. Dendrometer Measurements and Tree Circumference Increment Calculations
2.4. Biomass Inventory of the Individual Parts of the Model Trees
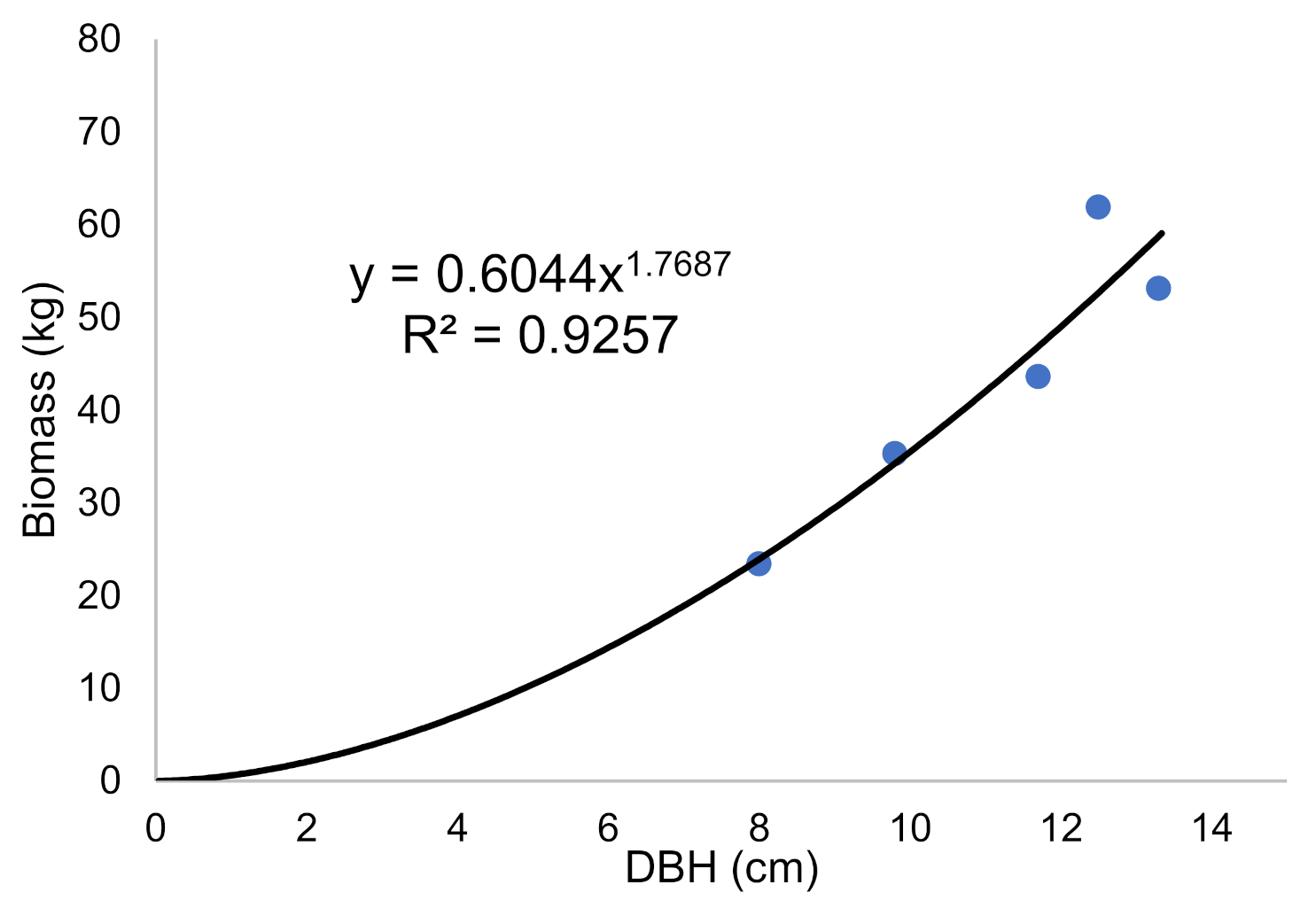
2.5. In Situ Allometric Equations—The Relationship between Biomass of Tree Components and Their DBH
2.6. Eddy Covariance Data Processing
2.7. Carbon Use Efficiency (CUE) and Definition of Vegetation Period
2.8. Drought Conditions Estimates
3. Results
3.1. Drought Occurrence Detected by SPEI Index Monitoring
3.2. Meteorological Conditions and CO2 and H2O Fluxes Courses
3.3. Identification of Specific Periods during Wood Growth
3.4. Stand Biomass and CUE Estimations
4. Discussion
4.1. NPP Biometric Estimates
4.2. Differences between Meteorological Conditions in 2019 and 2020 and Their Impact on NPP and GPP, and CUE Values
4.3. Difficulties in Calculating NPP as a Part of GPP Using Different Approaches
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wegiel, A.; Polowy, K. Aboveground carbon content and storage in mature scots pine stands of different densities. Forests 2020, 11, 240. [Google Scholar] [CrossRef] [Green Version]
- Peichl, M.; Brodeur, J.J.; Khomik, M.; Arain, M.A. Biometric and eddy-covariance based estimates of carbon fluxes in an age-sequence of temperate pine forests. Agric. For. Meteorol. 2010, 150, 952–965. [Google Scholar] [CrossRef]
- Dixon, R.K.; Solomon, A.M.; Brown, S.; Houghton, R.A.; Trexier, M.C.; Wisniewski, J. Carbon Pools and Flux of Global Forest Ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef]
- Denman, K.L.; Brasseur, G.; Chidthaisong, A.; Ciais, P.; Cox, P.M.; Dickinson, R.E.; Hauglustaine, D.; Heinze, C.; Holland, E.; Jacob, D.; et al. Couplings Between Changes in the Climate System and Biogeochemistry. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 501–568. [Google Scholar]
- Urban, J.; Čermák, J.; Ceulemans, R. Above- and below-ground biomass, surface and volume, and stored water in a mature Scots pine stand. Eur. J. For. Res. 2015, 134, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Manickam, V.; Krishna, I.V.M.; Shanti, S.K.; Radhika, R.; Campus, B.V.; Pradesh, A. Biomass Calculations for Carbon Sequestration in Forest Ecosystem. J. Energy Chem. Eng. 2014, 2, 30–38. [Google Scholar]
- Kunert, N.; El-Madany, T.S.; Aparecido, L.M.T.; Wolf, S.; Potvin, C. Understanding the controls over forest carbon use efficiency on small spatial scales: Effects of forest disturbance and tree diversity. Agric. For. Meteorol. 2019, 269–270, 136–144. [Google Scholar] [CrossRef]
- Juráň, S.; Grace, J.; Urban, O. Temporal Changes in Ozone Concentrations and Their Impact on Vegetation. Atmosphere 2021, 12, 82. [Google Scholar] [CrossRef]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kumar, M. Estimation of Biomass and Soil Carbon Stock in the Hydroelectric Catchment of India and its Implementation to Climate Change. J. Sustain. For. 2020, 39, 1–16. [Google Scholar] [CrossRef]
- Rai, P.; Vineeta; Shukla, G.; Manohar K, A.; Bhat, J.A.; Kumar, A.; Kumar, M.; Cabral-Pinto, M.; Chakravarty, S. Carbon storage of single tree and mixed tree dominant species stands in a reserve forest—Case study of the eastern sub-himalayan region of india. Land 2021, 10, 435. [Google Scholar] [CrossRef]
- Zianis, D.; Muukkonen, P.; Mäkipää, R.; Mencuccini, M. Biomass and Stem Volume Equations for Tree Species in Europe; The Finnish Society of Forest Science: Helsinki, Finland; The Finnish Forest Research Institute: Helsinki, Finland, 2005; Volume 4, ISBN 9514019830. [Google Scholar]
- Wohlfahrt, G.; Gu, L. The many meanings of gross photosynthesis and their implication for photosynthesis research from leaf to globe. Plant Cell Environ. 2015, 38, 2500–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, J.Q.; dos Santos, J.; Ribeiro, R.; Higuchi, N. Tree damage, allometric relationships, and aboveground net primary production in a tropical forest. For. Ecol. Manag. 2001, 152, 73–84. [Google Scholar] [CrossRef]
- Chapin, F.S.; Woodwell, G.M.; Randerson, J.T.; Rastetter, E.B.; Lovett, G.M.; Baldocchi, D.D.; Clark, D.A.; Harmon, M.E.; Schimel, D.S.; Valentini, R.; et al. Reconciling carbon-cycle concepts, terminology, and methods. Ecosystems 2006, 9, 1041–1050. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.A.; Brown, S.; Kicklighter, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, J. Measuring net primary production in forests: Concepts and field methods. Ecol. Appl. 2001, 11, 356–370. [Google Scholar] [CrossRef]
- Drake, J.E.; Davis, S.C.; Raetz, L.M.; Delucia, E.H. Mechanisms of age-related changes in forest production: The influence of physiological and successional changes. Glob. Chang. Biol. 2011, 17, 1522–1535. [Google Scholar] [CrossRef]
- Bruchwald, A. New Empirical Formulae for Determi- nation of Volume of Scots Pine Stands. Folia For. Pol. Ser. 1996, A 38, 5–10. [Google Scholar]
- DeLucia, E.H.; Drake, J.E.; Thomas, R.B.; Gonzalez-Meler, M. Forest carbon use efficiency: Is respiration a constant fraction of gross primary production? Glob. Chang. Biol. 2007, 13, 1157–1167. [Google Scholar] [CrossRef] [Green Version]
- Zanotelli, D.; Montagnani, L.; Manca, G.; Tagliavini, M. Net primary productivity, allocation pattern and carbon use efficiency in an apple orchard assessed by integrating eddy covariance, biometric and continuous soil chamber measurements. Biogeosciences 2013, 10, 3089–3108. [Google Scholar] [CrossRef] [Green Version]
- Law, B.E.; Sun, O.J.; Campbell, J.; Van Tuyl, S.; Thornton, P.E. Changes in carbon storage and fluxes in a chronosequence of ponderosa pine. Glob. Chang. Biol. 2003, 9, 510–524. [Google Scholar] [CrossRef]
- Doughty, C.E.; Goldsmith, G.R.; Raab, N.; Girardin, C.A.J.; Farfan-Amezquita, F.; Huaraca-Huasco, W.; Silva-Espejo, J.E.; Araujo-Murakami, A.; da Costa, A.C.L.; Rocha, W.; et al. What controls variation in carbon use efficiency among Amazonian tropical forests? Biotropica 2018, 50, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Anić, M.; Ostrogović Sever, M.Z.; Alberti, G.; Balenović, I.; Paladinić, E.; Peressotti, A.; Tijan, G.; Večenaj, Ž.; Vuletić, D.; Marjanović, H. Eddy covariance vs. biometric based estimates of net primary productivity of Pedunculate Oak (Quercus robur L.) forest in Croatia during ten years. Forests 2018, 9, 764. [Google Scholar] [CrossRef] [Green Version]
- Campioli, M.; Gielen, B.; Göckede, M.; Papale, D.; Bouriaud, O.; Granier, A. Temporal variability of the NPP-GPP ratio at seasonal and interannual time scales in a temperate beech forest. Biogeosciences 2011, 8, 2481–2492. [Google Scholar] [CrossRef] [Green Version]
- Małek, S.; Pająk, M.; Wosś, B.; Jasik, M. Zapas węgla w biomasie drzewostanów sosnowych o różnym wieku wokół stacji pomiarowych: Tuczno, Mezyk, Tlen1 i Tlen2. In Rola Lasu w Pochłanianiu Dwutlenku Węgla z Atmosfery; Olejnik, J., Małek, S., Eds.; Wydawnictwo Uniwersytetu Przyrodniczego w Poznaniu: Poznań, Poland, 2020; pp. 123–134. ISBN 978-83-7160-971-8. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Picard, N.; Saint-André, L.; Henry, M. Manual for Building Tree Volume and Biomass Allometric Equations: From Field Measurement to Prediction; Food and Agricultural Organization of the United Nations, Rome, and Centre de Coopération Internationale en Recherche Agronomique pour le Développement: Montpellier, France, 2012; ISBN 9789251073476. [Google Scholar]
- Mauder, M.; Foken, T. Impact of post-field data processing on eddy covariance flux estimates and energy balance closure. Meteorol. Zeitschrift 2006, 15, 597–609. [Google Scholar] [CrossRef]
- Barr, A.G.; Richardson, A.D.; Hollinger, D.Y.; Papale, D.; Arain, M.A.; Black, T.A.; Bohrer, G.; Dragoni, D.; Fischer, M.L.; Gu, L.; et al. Use of change-point detection for friction–velocity threshold evaluation in eddy-covariance studies. Agric. For. Meteorol. 2013, 171–172, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Šigut, L.; Menzer, O.; Reichstein, M. Basic and extensible post-processing of eddy covariance flux data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Dancho, M.; Vaughan, D. anomalize: Tidy Anomaly Detection. 2020. Available online: https://github.com/business-science/anomalize (accessed on 29 June 2021).
- LI COR, I. EddyPro® Software, Lincoln, Nebraska. 2019. Available online: https://www.licor.com/env/support/EddyPro/software.html (accessed on 29 June 2021).
- Morgenstern, K.; Black, T.A.; Humphreys, E.R.; Griffis, T.J.; Drewitt, G.B.; Cai, T.; Nesic, Z.; Spittlehouse, D.L.; Livingston, N.J. Sensitivity and uncertainty of the carbon balance of a Pacific Northwest Douglas-fir forest during an El Niño/La Niña cycle. Agric. For. Meteorol. 2004, 123, 201–219. [Google Scholar] [CrossRef]
- Ziemblińska, K.; Urbaniak, M.; Chojnicki, B.H.; Black, T.A.; Niu, S.; Olejnik, J. Net ecosystem productivity and its environmental controls in a mature Scots pine stand in north-western Poland. Agric. For. Meteorol. 2016, 228–229, 60–72. [Google Scholar] [CrossRef]
- Falge, E.M.; Clement, R.J.; Baldocchi, D.D.; Dolman, H.; Burba, G.G.; Anthoni, P.; Aubinet, M.; Olson, R.; Bernhofer, C.; Ceulemans, R.; et al. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agric. For. Meteorol. 2001, 107, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Gilmanov, T.G.; Johnson, D.A.; Saliendra, N.Z. Growing season CO2 fluxes in a sagebrush-steppe ecosystem in Idaho: Bowen ratio/energy balance measurements and modeling. Basic Appl. Ecol. 2003, 4, 167–183. [Google Scholar] [CrossRef]
- Körner, C. Leaf diffusive conductances in the major vegetation types of the globe. In Ecophysiology of Photosynthesis; Schulze, E.-D., Caldwell, M.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1995; pp. 463–490. ISBN 9788578110796. [Google Scholar]
- Lasslop, G.; Reichstein, M.; Papale, D.; Richardson, A.D.; Aarneth, A.; Barr, A.; Stoy, P.; Wohlfart, G. Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: Critical issues and global evaluation. Glob. Chang. Biol. 2010, 16, 187–208. [Google Scholar] [CrossRef] [Green Version]
- Zweifel, R.; Haeni, M.; Buchmann, N.; Eugster, W. Are trees able to grow in periods of stem shrinkage? New Phytol. 2016, 211, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Hinckley, T.M.; Bruckerhoff, D.N. The effects of drought on water relations and stem shrinkage of Quercus alba. Can. J. Bot. 1975, 53, 62–72. [Google Scholar] [CrossRef]
- Zweifel, R.; Zimmermann, L.; Newbery, D.M. Modeling tree water deficit from microclimate: An approach to quantifying drought stress. Tree Physiol. 2005, 25, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Beguería, S.; Latorre, B.; Reig, F.; Vicente-Serrano, S.M. SPEI Global Drought Monitor. Available online: https://spei.csic.es/map/maps.html#months=1%23month=10%23year=2020 (accessed on 6 January 2020).
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef] [Green Version]
- Beguería, S.; Vicente-Serrano, S.M.; Angulo-Martínez, M. A multiscalar global drought dataset: The SPEI base: A new gridded product for the analysis of drought variability and impacts. Bull. Am. Meteorol. Soc. 2010, 91, 1351–1356. [Google Scholar] [CrossRef] [Green Version]
- Zweifel, R.; Eugster, W.; Etzold, S.; Dobbertin, M.; Buchmann, N.; Häsler, R. Link between continuous stem radius changes and net ecosystem productivity of a subalpine Norway spruce forest in the Swiss Alps. New Phytol. 2010, 187, 819–830. [Google Scholar] [CrossRef]
- Xu, B.; Yang, Y.; Li, P.; Shen, H.; Fang, J. Global patterns of ecosystem carbon flux in forests: A biometric data-based synthesis. Glob. Biogeochem. Cycles 2014, 28, 962–973. [Google Scholar] [CrossRef]
- Ryan, M.G.; Phillips, N.; Bond, B.J. The hydraulic limitation hypothesis revisited. Plant Cell Environ. 2006, 29, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Ball, J.T.; Woodrow, I.E.; Berry, J.A. A Model Predicting Stomatal Conductance and Its Contribution to the Control of Photosynthesis under Different Environmental Conditions; Biggins, J., Ed.; Springer: Amsterdam, The Netherlands, 1987. [Google Scholar]
- Maseyk, K.; Grünzweig, J.M.; Rotenberg, E.; Yakir, D. Respiration acclimation contributes to high carbon-use efficiency in a seasonally dry pine forest. Glob. Chang. Biol. 2008, 14, 1553–1567. [Google Scholar] [CrossRef]
- Waring, R.H.; Landsberg, J.J.; Williams, M. Net primary production of forests: A constant fraction of gross primary production? Tree Physiol. 1998, 18, 129–134. [Google Scholar] [CrossRef]
- Gifford, R.M. Plant respiration in productivity models: Conceptualisation, representation and issues for global terrestrial carbon-cycle research. Funct. Plant Biol. 2003, 30, 171–186. [Google Scholar] [CrossRef]
- Thornley, J.H.M.; Cannell, M.G.R. Modelling the components of plant respiration: Representation and realism. Ann. Bot. 2000, 85, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Cannell, M.G.R.; Dewar, R.C. Carbon Allocation in Trees: A Review of Concepts for Modelling. Adv. Ecol. Res. 1994, 25, 59–104. [Google Scholar]
- Leuschner, C.; Backes, K.; Hertel, D.; Schipka, F.; Schmitt, U.; Terborg, O.; Runge, M. Drought responses at leaf, stem and fine root levels of competitive Fagus sylvatica L. and Quercus petraea (Matt.) Liebl. trees in dry and wet years. For. Ecol. Manag. 2001, 149, 33–46. [Google Scholar] [CrossRef]
- Granier, A.; Reichstein, M.; Bréda, N.; Janssens, I.A.; Falge, E.; Ciais, P.; Grünwald, T.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; et al. Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year: 2003. Agric. For. Meteorol. 2007, 143, 123–145. [Google Scholar] [CrossRef]
- Luyssaert, S.; Inglima, I.; Jung, M.; Richardson, A.D.; Reichstein, M.; Papale, D.; Piao, S.L.; Schulze, E.-D.; Wingate, L.; Matteucci, G.; et al. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Glob. Chang. Biol. 2007, 13, 2509–2537. [Google Scholar] [CrossRef] [Green Version]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Lacointe, A. Carbon allocation among tree organs: A review of basic processes and representation in functional-structural tree models. Ann. For. Sci. 2000, 57, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Curtis, P.S.; Hanson, P.J.; Bolstad, P.; Barford, C.; Randolph, J.; Schmid, H.; Wilson, K.B. Biometric and eddy-covariance based estimates of annual carbon storage in five eastern North American deciduous forests. Agric. For. Meteorol. 2002, 113, 3–19. [Google Scholar] [CrossRef]
- Ketterings, Q.M.; Coe, R.; Van Noordwijk, M.; Ambagau’, Y.; Palm, C.A. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. For. Ecol. Manag. 2001, 146, 199–209. [Google Scholar] [CrossRef]
- Black, K.; Bolger, T.; Davis, P.; Nieuwenhuis, M.; Reidy, B.; Saiz, G.; Tobin, B.; Osborne, B. Inventory and eddy covariance-based estimates of annual carbon sequestration in a Sitka spruce (Picea sitchensis (Bong.) Carr.) forest ecosystem. Eur. J. For. Res. 2005, 126, 167–178. [Google Scholar] [CrossRef]
- Gough, C.M.; Vogel, C.S.; Schmid, H.P.; Su, H.B.; Curtis, P.S. Multi-year convergence of biometric and meteorological estimates of forest carbon storage. Agric. For. Meteorol. 2008, 148, 158–170. [Google Scholar] [CrossRef]
- Ehman, J.L.; Schmid, H.P.; Grimmond, C.S.B.; Randolph, J.C.; Hanson, P.J.; Wayson, C.A.; Cropley, F.D. An initial intercomparison of micrometeorological and ecological inventory estimates of carbon exchange in a mid-latitude deciduous forest. Glob. Chang. Biol. 2002, 8, 575–589. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Saigusa, N.; Koizumi, H. On linking multiyear biometric measurements of tree growth with eddy covariance-based net ecosystem production. Glob. Chang. Biol. 2009, 15, 1015–1024. [Google Scholar] [CrossRef]
- Keith, H.; Leuning, R.; Jacobsen, K.L.; Cleugh, H.A.; van Gorsel, E.; Raison, R.J.; Medlyn, B.E.; Winters, A.; Keitel, C. Multiple measurements constrain estimates of net carbon exchange by a Eucalyptus forest. Agric. For. Meteorol. 2009, 149, 535–558. [Google Scholar] [CrossRef]
- Collalti, A.; Prentice, I.C. Is NPP proportional to GPP? Waring’s hypothesis 20 years on. Tree Physiol. 2019, 39, 1473–1483. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hollinger, D.Y.; Burba, G.G.; Davis, K.J.; Flanagan, L.B.; Katul, G.G.; William Munger, J.; Ricciuto, D.M.; Stoy, P.C.; Suyker, A.E.; et al. A multi-site analysis of random error in tower-based measurements of carbon and energy fluxes. Agric. For. Meteorol. 2006, 136, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Haslwanter, A.; Hammerle, A.; Wohlfahrt, G. Open-path vs. closed-path eddy covariance measurements of the net ecosystem carbon dioxide and water vapour exchange: A long-term perspective. Agric. For. Meteorol. 2009, 149, 291–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deventer, M.J.; Roman, T.; Bogoev, I.; Kolka, R.K.; Erickson, M.; Lee, X.; Baker, J.M.; Millet, D.B.; Griffis, T.J. Biases in open-path carbon dioxide flux measurements: Roles of instrument surface heat exchange and analyzer temperature sensitivity. Agric. For. Meteorol. 2021, 296. [Google Scholar] [CrossRef]
- Tanaka, A. Photosynthetic activity in winter needles of the evergreen tree Taxus cuspidata at low temperatures. Tree Physiol. 2007, 27, 641–648. [Google Scholar] [CrossRef]
- Goldstein, G.; Nobel, P.S. Water relations and low-temperature acclimation for cactus species varying in freezing tolerance. Plant Physiol. 1994, 104, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Uemura, M.; Steponkus, P.L. A contrast of the plasma membrane lipid composition of oat and rye leaves in relation to freezing tolerance. Plant Physiol. 1994, 104, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Steponkus, P.L.; Uemura, M.; Joseph, R.A.; Gilmour, S.J.; Thomashow, M.F. Mode of action of the COR15a gene on the freezing tolerance of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1998, 95, 14570–14575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hare, P.D.; Cress, W.A.; Van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Thomashow, M.F. PLANT COLD ACCLIMATION: Freezing Tolerance Genes and Regulatory Mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | DBH | Stem with Bark | Bark | Main Branches | Fine Branches | Needles | Fine Roots | Main Roots | Total (Whole Tree) | AGB (% DB) | BGB (% DB) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 8.0 | 12.9 | 1.7 | 2.3 | 1.5 | 1.2 | 0.95 | 2.8 | 23.43 | 84.0 | 16.0 |
| 2 | 9.8 | 17.5 | 2.3 | 4.3 | 3.2 | 2.8 | 0.94 | 4.3 | 35.32 | 85.1 | 14.9 |
| 3 | 11.7 | 23.5 | 3.1 | 6.1 | 2.9 | 2.5 | 0.98 | 4.6 | 43.61 | 87.3 | 12.7 |
| 4 | 12.5 | 28.4 | 4.3 | 12.7 | 4.2 | 3.1 | 1.02 | 8.2 | 61.89 | 85.0 | 15.0 |
| 5 | 13.3 | 29.9 | 4.6 | 6.0 | 3.5 | 2.8 | 1.01 | 7.9 | 53.13 | 88.2 | 11.8 |
| Average | 11.1 | 22.4 | 3.2 | 6.3 | 3.1 | 2.5 | 0.98 | 5.6 | 43.47 | 85.9 | 14.1 |
| YEAR | P (mm) | Tair (°C) | VPD (kPa) | Rg (Wm−2) | SWC (%) | GPP (gCm−2) | Reco (gCm−2) | ET (mm) |
|---|---|---|---|---|---|---|---|---|
| 2019 | 509 | 9.87 | 0.37 | 127.7 | 6.7 | 1724 | 1284 | 479 |
| 2020 | 624 | 9.73 | 0.33 | 127.2 | 8.6 | 1727 | 1316 | 493 |
| Spring 2019 | 116 | 9.32 | 0.49 | 165.3 | 7.7 | 536 | 301 | 129 |
| Spring 2020 | 98 | 7.97 | 0.39 | 191.4 | 9.9 | 511 | 259 | 140 |
| Summer 2019 | 70 | 18.48 | 0.76 | 238.4 | 5.1 | 776 | 527 | 168 |
| Summer 2020 | 232 | 17.45 | 0.58 | 209.7 | 5.5 | 765 | 575 | 181 |
| Autumn 2019 | 191 | 9.64 | 0.12 | 71.8 | 7.1 | 314 | 330 | 112 |
| Autumn 2020 | 132 | 10.32 | 0.25 | 75.8 | 7.0 | 317 | 352 | 100 |
| Year | 2019 | 2020 | ||
|---|---|---|---|---|
| M1 | M2 | M1 | M2 | |
| Total stem increment of an average tree (cm)—dendrometers | 0.142 ± 0.01 | - | 0.164 ± 0.01 | - |
| Total increase in dry biomass of an average tree (kg) | 0.942 ± 0.070 | 0.978 ± 0.072 | 1.206 ± 0.070 | 1.255 ± 0.073 |
| Total biomass of the stand at the end of growing seasons (t ha−1) | 203.387 ± 0.295 | 204.017 ± 0.306 | 208.861 ± 0.298 | 209.714 ± 0.310 |
| Total increase in dry biomass of the stand (t ha−1) | 4.273 ± 0.295 | 4.439 ± 0.306 | 5.473 ± 0.298 | 5.697 ± 0.310 |
| Total NPP (t C ha−1) | 2.137 ± 0.148 | 2.220 ± 0.153 | 2.737 ± 0.149 | 2.849 ± 0.155 |
| GPP total during vegetation period (t C ha−1) | 14.70 | 14.49 | ||
| NEP total during vegetation period (t C ha−1) | 5.00 | 4.50 | ||
| CUE for vegetation period | 0.15 | 0.15 | 0.19 | 0.20 |
| GPP total during B (t C ha−1) | 11.79 | 11.74 | ||
| NEP total during B (t C ha−1) | 3.70 | 3.19 | ||
| CUE for B | 0.18 | 0.19 | 0.23 | 0.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dukat, P.; Ziemblińska, K.; Olejnik, J.; Małek, S.; Vesala, T.; Urbaniak, M. Estimation of Biomass Increase and CUE at a Young Temperate Scots Pine Stand Concerning Drought Occurrence by Combining Eddy Covariance and Biometric Methods. Forests 2021, 12, 867. https://doi.org/10.3390/f12070867
Dukat P, Ziemblińska K, Olejnik J, Małek S, Vesala T, Urbaniak M. Estimation of Biomass Increase and CUE at a Young Temperate Scots Pine Stand Concerning Drought Occurrence by Combining Eddy Covariance and Biometric Methods. Forests. 2021; 12(7):867. https://doi.org/10.3390/f12070867
Chicago/Turabian StyleDukat, Paulina, Klaudia Ziemblińska, Janusz Olejnik, Stanisław Małek, Timo Vesala, and Marek Urbaniak. 2021. "Estimation of Biomass Increase and CUE at a Young Temperate Scots Pine Stand Concerning Drought Occurrence by Combining Eddy Covariance and Biometric Methods" Forests 12, no. 7: 867. https://doi.org/10.3390/f12070867