
Transcriptomic and Anatomic Profiling Reveal Etiolation Promotes Adventitious Rooting by Exogenous Application of 1-Naphthalene Acetic Acid in Robinia pseudoacacia L.


 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Histological Changes for the Process of Adventitious Root Formation in R. pseudoacacia
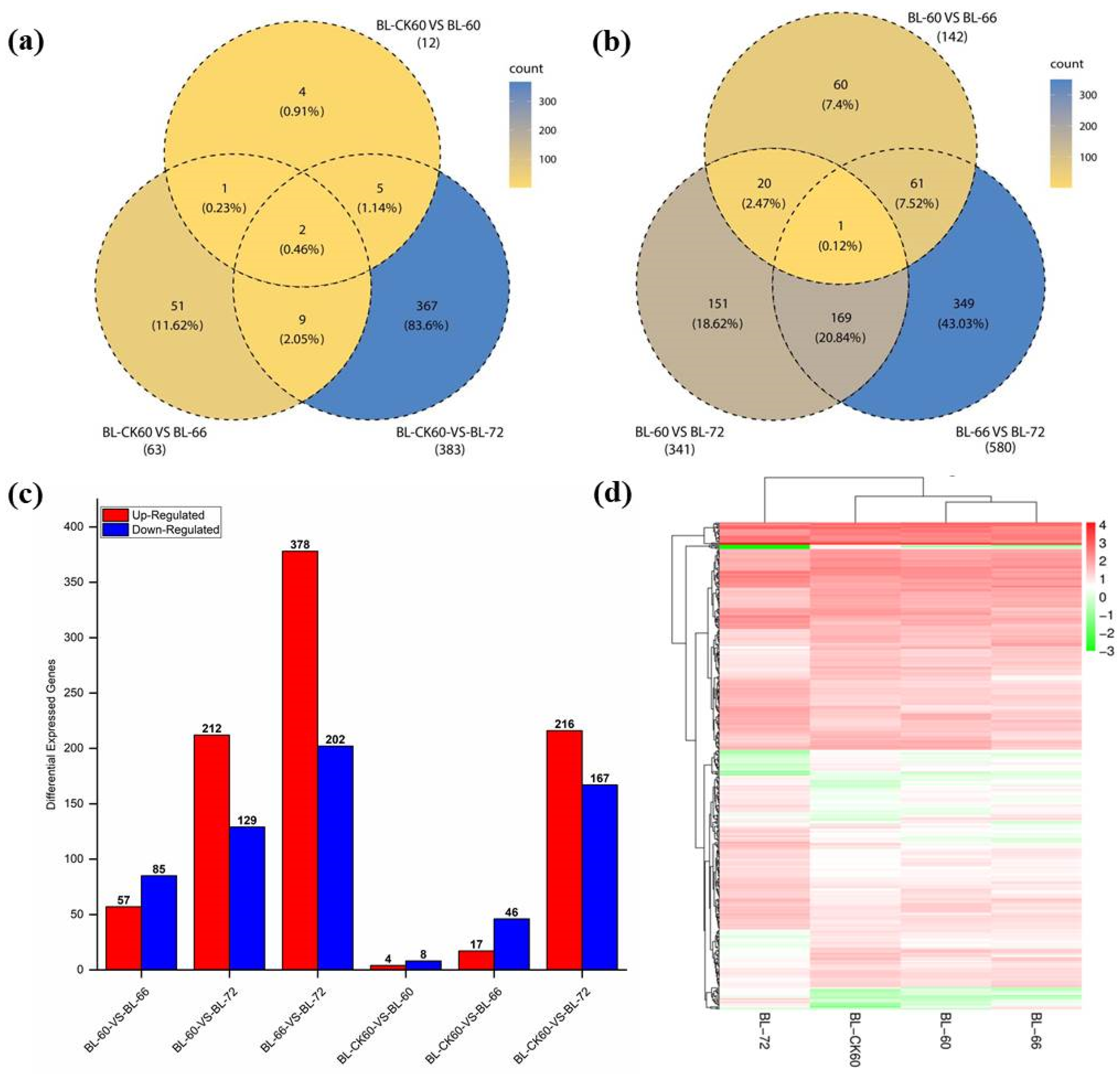
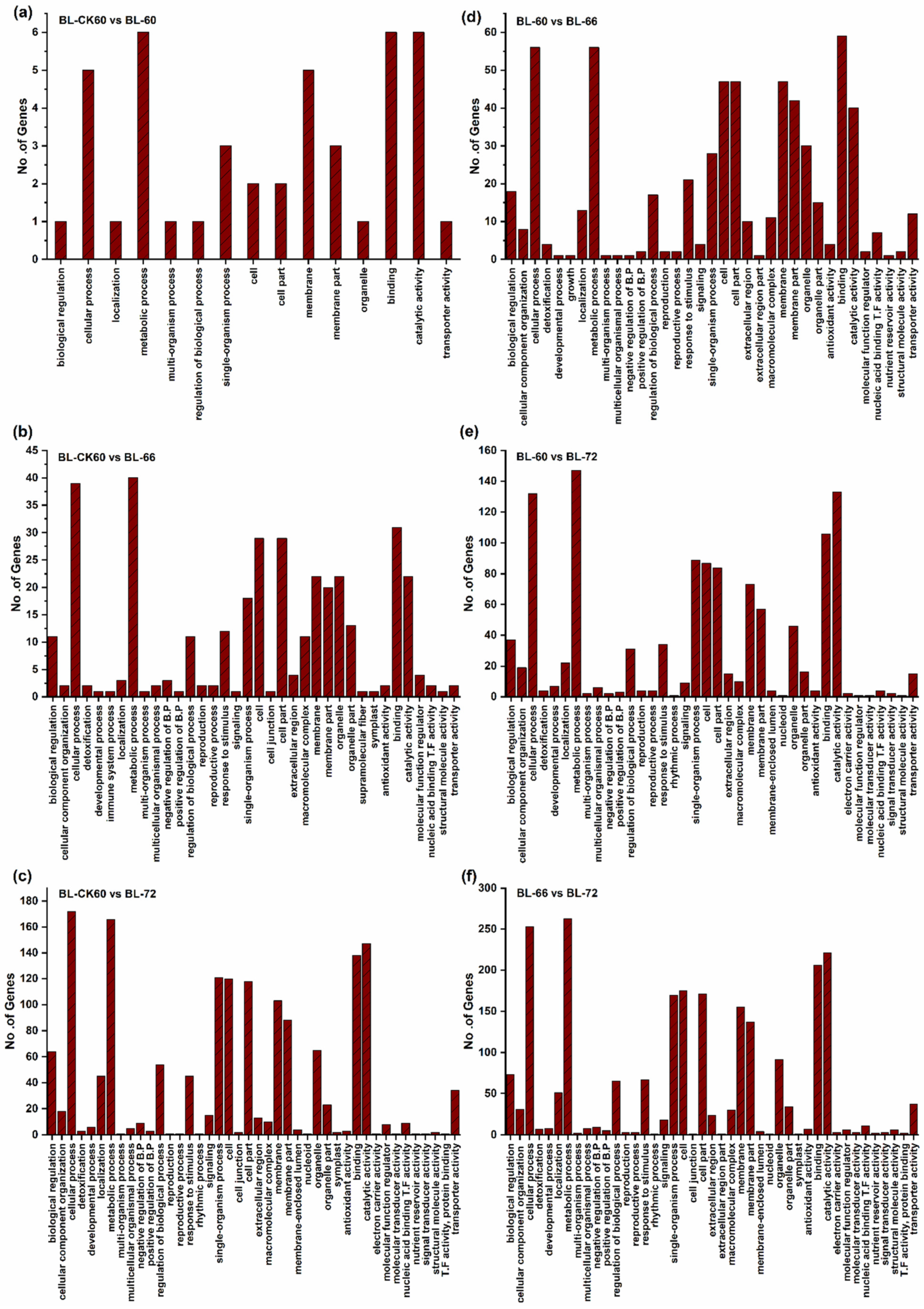
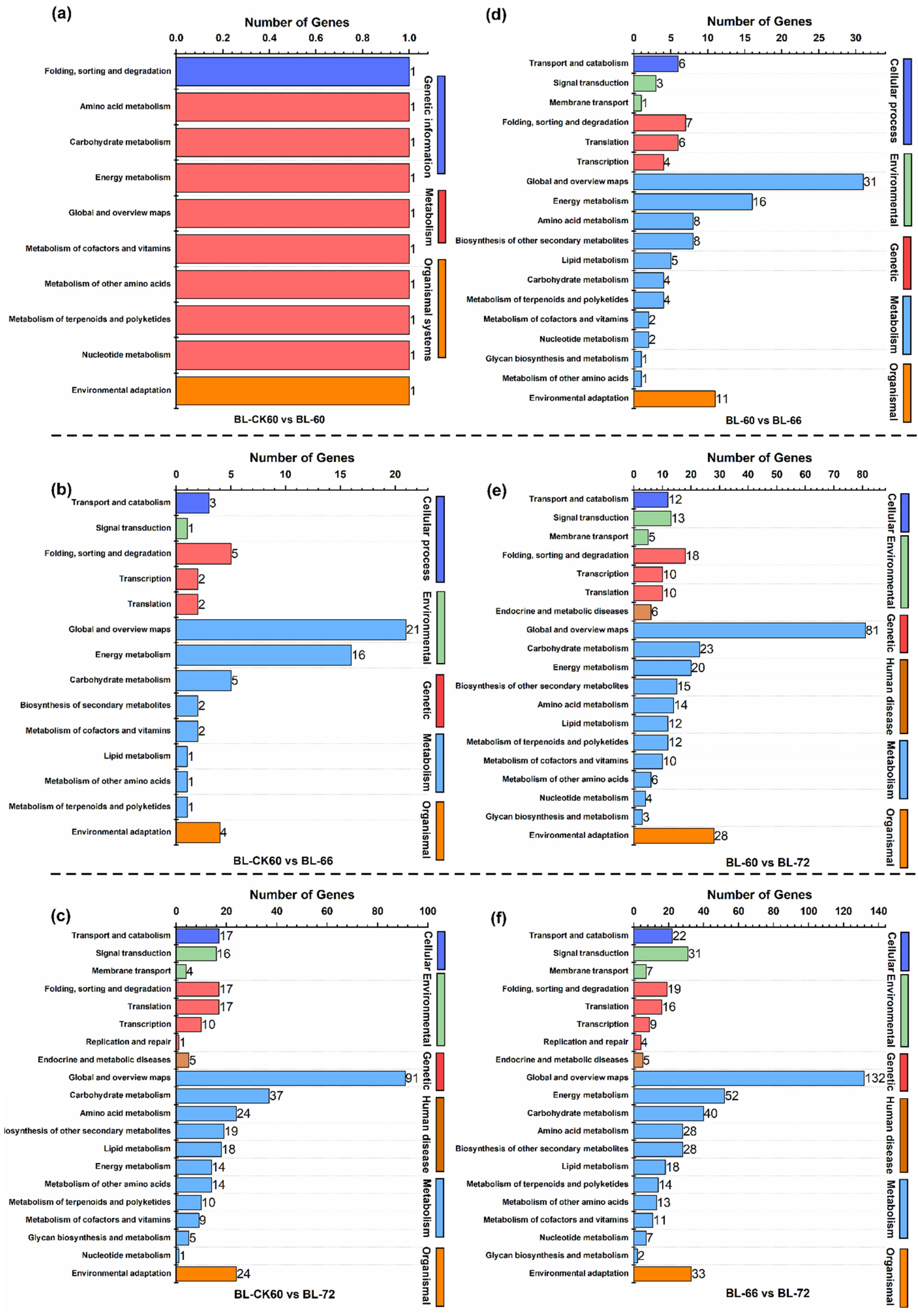
2.2. Deep Sequencing, Functional Annotation of DEGs, and qRT-PCR Analysis
2.3. Effect of Exogenous NAA on Plant Hormone Signal Transduction Pathway
2.3.1. Auxin
2.3.2. Cytokinin
2.3.3. Abscisic Acid
2.3.4. Gibberellic Acid
2.3.5. Jasmonic Acid, Brassinosteroids, and Salicylic Acid
2.3.6. Starch and Sucrose Metabolism Pathway
2.3.7. Phenylpropanoid Biosynthesis Pathway
2.3.8. Carbon Metabolism Pathway
3. Discussion
3.1. Histological Changes for the Process of Adventitious Root Formation in R. pseudoacacia
3.2. Functional Annotation of DEGs and Expression Analysis
3.3. Plant Hormone Signal Transduction Pathway
3.3.1. Auxin
3.3.2. Cytokinin
3.3.3. Abscisic Acid
3.3.4. Gibberellic Acid
3.3.5. Jasmonic Acid, Brassinosteroids and Salicylic Acid
3.3.6. Starch and Sucrose Metabolism Pathway
3.3.7. Phenylpropanoid Biosynthesis Pathway
3.3.8. Carbon Metabolism Pathway
4. Materials and Methods
4.1. Plant Material
4.2. Anatomical and Microscopic Analysis
4.3. RNA Extraction, Deep Sequencing, Functional Annotation, Pathway Enrichment, and qRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.; Sun, Y.; Han, C.; Dong, L.; Guo, Q.; Li, X.; Li, Y. Integrated Analysis of the Physiology, RNA, and microRNA Involved in Black Locust (Robinia pseudoacacia) Rejuvenation. 2019. Available online: http://doi.org/10.21203/rs.2.10375/v1 (accessed on 16 March 2021).
- Rédei, K. Black Locust (Robinia pseudoacacia L.) Growing in Hungary; Publications of the Hungarian Forest Research Institute: Budapest, Ungarn, 1998. [Google Scholar]
- Rédei, K.; Keserű, Z.; Csiha, I.; Rásó, J.; Bakti, B.; Takács, M. Improvement of black locust (Robinia pseudoacacia L.) growing under marginal site conditions in Hungary: Case studies. Acta Agrar. Debr. 2018, 74, 129–133. [Google Scholar] [CrossRef]
- Ragonezi, C.; Klimaszewska, K.; Castro, M.R.; Lima, M.; de Oliveira, P.; Zavattieri, M.A. Adventitious rooting of conifers: Influence of physical and chemical factors. Trees 2010, 24, 975–992. [Google Scholar] [CrossRef] [Green Version]
- Meng, B.N.; Peng, Z.D.; Zhang, Z.L.; Xu, H.M.; Li, Y. Research on cuttage of tetraploid black locust (Robinia pseudoacacia L.) hardwood treated by low temperature sand storage and plant growth regulator. Heilongjiang Agric. Sc. 2010, 8, 85–88. [Google Scholar]
- Da Costa, C.T.; De Almeida, M.R.; Ruedell, C.M.; Schwambach, J.; Maraschin, F.D.S.; Fett-Neto, A.G. When stress and development go hand in hand: Main hormonal controls of adventitious rooting in cuttings. Front. Plant Sci. 2013, 4, 133. [Google Scholar] [CrossRef] [Green Version]
- Bellini, C.; Pacurar, D.I.; Perrone, I. Adventitious roots and lateral roots: Similarities and differences. Annu. Rev. Plant Biol. 2014, 65, 639–666. [Google Scholar] [CrossRef]
- Lu, N.; Dai, L.; Luo, Z.; Wang, S.; Wen, Y.; Duan, H.; Li, Y. Characterization of the transcriptome and gene expression of tetraploid black locust cuttings in response to etiolation. Genes 2017, 8, 345. [Google Scholar] [CrossRef] [Green Version]
- Klopotek, Y.; Haensch, K.T.; Hause, B.; Hajirezaei, M.R.; Druege, U. Dark exposure of petunia cuttings strongly improves adventitious root formation and enhances carbohydrate availability during rooting in the light. J. Plant Physiol. 2010, 167, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Lakehal, A.; Bellini, C. Control of adventitious root formation: Insights into synergistic and antagonistic hormonal interactions. Physiol. Plant. 2019, 165, 90–100. [Google Scholar] [CrossRef] [Green Version]
- De Klerk, G.-J.; Guan, H.; Huisman, P.; Marinova, S. Effects of phenolic compounds on adventitious root formation and oxidative decarboxylation of applied indoleacetic acid in Malus ‘Jork 9’. Plant Growth Regul. 2011, 63, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Kevers, C.; Hausman, J.F.; Faivre-Rampant, O.; Dommes, J.; Gaspar, T. What We Have Learned about the Physiology of In Vitro Adventitious Rooting of Woody Plants and How It Relates to Improvements in the Practice; Research Signpost: Kerala, India, 2009; pp. 209–225. [Google Scholar]
- Zaman, M.; Kurepin, L.V.; Catto, W.; Pharis, R.P. Evaluating the use of plant hormones and biostimulators in forage pastures to enhance shoot dry biomass production by perennial ryegrass (Lolium perenne L.). J. Sci. Food Agric. 2016, 96, 715–726. [Google Scholar] [CrossRef]
- Richards, M.R.; Rupp, L.A. Etiolation improves rooting of bigtooth maple cuttings. HortTechnology 2012, 22, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Batten, D.; Mullins, M. Ethylene and adventitious root formation in hypocotyl segments of etiolated mung-bean (Vigna radiata (L.) Wilczek) seedlings. Planta 1978, 138, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Pacurar, D.I.; Perrone, I.; Bellini, C. Auxin is a central player in the hormone cross-talks that control adventitious rooting. Physiol. Plant. 2014, 151, 83–96. [Google Scholar] [CrossRef] [PubMed]
- De Klerk, G.-J.; Van Der Krieken, W.; de Jong, J.C. Review the formation of adventitious roots: New concepts, new possibilities. Vitr. Cell. Dev. Biol. Plant 1999, 35, 189–199. [Google Scholar] [CrossRef]
- Legué, V.; Rigal, A.; Bhalerao, R.P. Adventitious root formation in tree species: Involvement of transcription factors. Physiol. Plant. 2014, 151, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Olatunji, D.; Geelen, D.; Verstraeten, I. Control of endogenous auxin levels in plant root development. Int. J. Mol. Sci. 2017, 18, 2587. [Google Scholar] [CrossRef] [Green Version]
- Wiesman, Z.; Lavee, S. Enhancement of IBA stimulatory effect on rooting of olive cultivar stem cuttings. Sci. Hortic. 1995, 62, 189–198. [Google Scholar] [CrossRef]
- Li, S.-W.; Xue, L.; Xu, S.; Feng, H.; An, L. Hydrogen peroxide acts as a signal molecule in the adventitious root formation of mung bean seedlings. Environ. Exp. Bot. 2009, 65, 63–71. [Google Scholar] [CrossRef]
- Koyuncu, F.; Balta, F. Adventitious root formation in leaf-bud cuttings of tea (Camellia sinensis L.). Pak. J. Bot. 2004, 36, 763–768. [Google Scholar]
- Goldfarb, B.; Hackett, W.P.; Furnier, G.R.; Mohn, C.A.; Plietzsch, A. Adventitious root initiation in hypocotyl and epicotyl cuttings of eastern white pine (Pinus strobus) seedlings. Physiol. Plant. 1998, 102, 513–522. [Google Scholar] [CrossRef]
- Smith, N.; Wareing, P. The distribution of latent root primordia in stems of Populus× robusta, and factors affecting the emergence of preformed roots from cuttings. For. Int. J. For. Res. 1972, 45, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Carlson, M.C. Nodal adventitious roots in willow stems of different ages. Am. J. Bot. 1950, 37, 555–561. [Google Scholar] [CrossRef]
- Weijers, D.; Franke-van Dijk, M.; Vencken, R.J.; Quint, A.; Hooykaas, P.; Offringa, R. An Arabidopsis Minute-like phenotype caused by a semi-dominant mutation in a RIBOSOMAL PROTEIN S5 gene. Development 2001, 128, 4289–4299. [Google Scholar] [CrossRef] [PubMed]
- Garrido, G.; Ramón Guerrero, J.; Angel Cano, E.; Acosta, M.; Sánchez-Bravo, J. Origin and basipetal transport of the IAA responsible for rooting of carnation cuttings. Physiol. Plant. 2002, 114, 303–312. [Google Scholar] [CrossRef]
- Ren, H.; Hu, H.; Luo, X.; Zhang, C.; Li, X.; Li, P.; Shen, C. Dynamic changes of phytohormone signaling in the base of Taxus media stem cuttings during adventitious root formation. Sci. Hortic. 2019, 246, 338–346. [Google Scholar] [CrossRef]
- Dumas, E.; Monteuuis, O. In vitro rooting of micropropagated shoots from juvenile and mature Pinus pinaster explants: Influence of activated charcoal. Plant Cell Tissue Organ Cult. 1995, 40, 231–235. [Google Scholar] [CrossRef]
- Yang, G.; Chen, S.; Wang, S.; Liu, G.; Li, H.; Huang, H.; Jiang, J. BpGH3. 5, an early auxin-response gene, regulates root elongation in Betula platyphylla× Betula pendula. Plant Cell Tissue Organ Cult. 2015, 120, 239–250. [Google Scholar] [CrossRef]
- Keuskamp, D.H.; Pollmann, S.; Voesenek, L.A.; Peeters, A.J.; Pierik, R. Auxin transport through PIN-FORMED 3 (PIN3) controls shade avoidance and fitness during competition. Proc. Natl. Acad. Sci. USA 2010, 107, 22740–22744. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Carvajal, G.A.; Morse, A.M.; Dervinis, C.; Davis, J.M. The cytokinin type-B response regulator PtRR13 is a negative regulator of adventitious root development in Populus. Plant Physiol. 2009, 150, 759–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naseem, M.; Kaltdorf, M.; Dandekar, T. The nexus between growth and defence signalling: Auxin and cytokinin modulate plant immune response pathways. J. Exp. Bot. 2015, 66, 4885–4896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Rizzo, S.; Crespi, M.; Frugier, F. The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. Plant Cell 2006, 18, 2680–2693. [Google Scholar] [CrossRef] [Green Version]
- Bonke, M.; Thitamadee, S.; Mähönen, A.P.; Hauser, M.T.; Helariutta, Y. APL regulates vascular tissue identity in Arabidopsis. Nature 2003, 426, 181–186. [Google Scholar] [CrossRef]
- Prabu, G.; Prasad, D.T. Functional characterization of sugarcane MYB transcription factor gene promoter (PScMYBAS1) in response to abiotic stresses and hormones. Plant Cell Rep. 2012, 31, 661–669. [Google Scholar] [CrossRef]
- De Smet, I.; Zhang, H.; Inze, D.; Beeckman, T. A novel role for abscisic acid emerges from underground. Trends Plant Sci. 2006, 11, 434–439. [Google Scholar] [CrossRef]
- Dawood, T.; Yang, X.; Visser, E.J.; Te Beek, T.A.; Kensche, P.R.; Cristescu, S.M.; Rieu, I. A co-opted hormonal cascade activates dormant adventitious root primordia upon flooding in Solanum dulcamara. Plant Physiol. 2016, 170, 2351–2364. [Google Scholar] [CrossRef] [Green Version]
- Goessen, R. Exploring the Involvement of Gene Regulatory Variation in Wheat Improvement with Network Analysis. 2018. Available online: https://edepot.wur.nl/433739 (accessed on 16 March 2021).
- Shkolnik-Inbar, D.; Bar-Zvi, D. ABI4 mediates abscisic acid and cytokinin inhibition of lateral root formation by reducing polar auxin transport in Arabidopsis. Plant Cell 2010, 22, 3560–3573. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.-J.; Parkin, I.; Lydiate, D.; Hannoufa, A. An auxin-responsive SCARECROW-like transcriptional activator interacts with histone deacetylase. Plant Mol. Biol. 2004, 55, 417–431. [Google Scholar] [CrossRef]
- Vielba, J.M.; Díaz-Sala, C.; Ferro, E.; Rico, S.; Lamprecht, M.; Abarca, D.; Sánchez, C. CsSCL1 is differentially regulated upon maturation in chestnut microshoots and is specifically expressed in rooting-competent cells. Tree Physiol. 2011, 31, 1152–1160. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.-F.; Yang, S.Y.; Chen, K.T.; Hsing, Y.I.; Zeevaart, J.A.; Chen, L.J.; Yu, S.M. A novel class of gibberellin 2-oxidases control semidwarfism, tillering, and root development in rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffens, B.; Sauter, M. Epidermal cell death in rice is regulated by ethylene, gibberellin, and abscisic acid. Plant Physiol. 2005, 139, 713–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Liang, Y.; Xing, L.; Mao, J.; Liu, Z.; Dong, F.; Zhang, D. Transcriptome analysis reveals multiple hormones, wounding and sugar signaling pathways mediate adventitious root formation in apple rootstock. Int. J. Mol. Sci. 2018, 19, 2201. [Google Scholar] [CrossRef] [Green Version]
- Agulló-Antón, M.Á.; Ferrández-Ayela, A.; Fernández-García, N.; Nicolás, C.; Albacete, A.; Pérez-Alfocea, F.; Acosta, M. Early steps of adventitious rooting: Morphology, hormonal profiling and carbohydrate turnover in carnation stem cuttings. Physiol. Plant. 2014, 150, 446–462. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Mongelard, G.; Floková, K.; Păcurar, D.I.; Novák, O.; Staswick, P.; Bellini, C. Auxin controls Arabidopsis adventitious root initiation by regulating jasmonic acid homeostasis. Plant Cell 2012, 24, 2515–2527. [Google Scholar] [CrossRef] [Green Version]
- Lischweski, S.; Muchow, A.; Guthörl, D.; Hause, B. Jasmonates act positively in adventitious root formation in petunia cuttings. BMC Plant Biol. 2015, 15, 229. [Google Scholar] [CrossRef] [Green Version]
- Kazan, K.; Manners, J.M. MYC2: The master in action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.-T.; Yu, X.Y.; Zhu, Y.J.; Zhuang, M.; Zhang, Z.M.; Jin, L.; Jin, F.J. Research progress on the basic helix-loop-helix transcription factors of Aspergillus species. Adv. Appl. Microbiol. 2019, 109, 31–59. [Google Scholar] [PubMed]
- Xu, W.; Campbell, P.; Vargheese, A.K.; Braam, J. The Arabidopsis XET-related gene family: Environmental and hormonal regulation of expression. Plant J. 1996, 9, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Purugganan, M.M.; Polisensky, D.H.; Antosiewicz, D.M.; Fry, S.C.; Braam, J. Arabidopsis TCH4, regulated by hormones and the environment, encodes a xyloglucan endotransglycosylase. Plant Cell 1995, 7, 1555–1567. [Google Scholar] [PubMed] [Green Version]
- Cai, C.; Guo, W.; Zhang, B. Genome-wide identification and characterization of SPL transcription factor family and their evolution and expression profiling analysis in cotton. Sci. Rep. 2018, 8, 762. [Google Scholar]
- Yu, N.; Niu, Q.W.; Ng, K.H.; Chua, N.H. The role of miR156/SPL s modules in Arabidopsis lateral root development. Plant J. 2015, 83, 673–685. [Google Scholar] [CrossRef]
- OuYang, F.; Wang, J.; Li, Y. Effects of cutting size and exogenous hormone treatment on rooting of shoot cuttings in Norway spruce [Picea abies (L.) Karst.]. New For. 2015, 46, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Li, Z.; Yuan, H.; Fang, P.; Chen, X.; Li, W. Proper gibberellin localization in vascular tissue is required to regulate adventitious root development in tobacco. J. Exp. Bot. 2013, 64, 3411–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruedell, C.M.; de Almeida, M.R.; Fett-Neto, A.G. Concerted transcription of auxin and carbohydrate homeostasis-related genes underlies improved adventitious rooting of microcuttings derived from far-red treated Eucalyptus globulus Labill mother plants. Plant Physiol. Biochem. 2015, 97, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Neuteboom, L.W.; Veth-Tello, L.M.; Clijdesdale, O.R.; Hooykaas, P.J.; van der Zaal, B.J. A novel subtilisin-like protease gene from Arabidopsis thaliana is expressed at sites of lateral root emergence. DNA Res. 1999, 6, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.; Frugis, G.; Colgan, D.; Chua, N.H. Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development. Genes Dev. 2000, 14, 3024–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.-W.; Shi, R.F.; Leng, Y.; Zhou, Y. Transcriptomic analysis reveals the gene expression profile that specifically responds to IBA during adventitious rooting in mung bean seedlings. BMC Genom. 2016, 17, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakabayashi, K.; Sakurai, N.; Kuraishi, S. Effects of abscisic acid on the synthesis of cell-wall polysaccharides in segments of etiolated squash hypocotyl II. Levels of UDP-neutral sugars. Plant Cell Physiol. 1991, 32, 427–432. [Google Scholar] [CrossRef]
- Ciereszko, I.; Johansson, H.; Kleczkowski, L.A. Interactive effects of phosphate deficiency, sucrose and light/dark conditions on gene expression of UDP-glucose pyrophosphorylase in Arabidopsis. J. Plant Physiol. 2005, 162, 343–353. [Google Scholar] [CrossRef]
- Abu-Abied, M.; Szwerdszarf, D.; Mordehaev, I.; Levy, A.; Belausov, E.; Yaniv, Y.; Sadot, E. Microarray analysis revealed upregulation of nitrate reductase in juvenile cuttings of Eucalyptus grandis, which correlated with increased nitric oxide production and adventitious root formation. Plant J. 2012, 71, 787–799. [Google Scholar] [CrossRef]
- Wei, K.; Wang, L.Y.; Wu, L.Y.; Zhang, C.C.; Li, H.L.; Tan, L.Q.; Cheng, H. Transcriptome analysis of indole-3-butyric acid-induced adventitious root formation in nodal cuttings of Camellia sinensis (L.). PLoS ONE 2014, 9, e107201. [Google Scholar] [CrossRef] [Green Version]
- Boudet, A.; Lapierre, C.; Grima-Pettenati, J. Tansley review No. 80. Biochemistry and molecular biology of lignification. New Phytol. 1995, 129, 203–236. [Google Scholar] [CrossRef]
- Tamagnone, L.; Merida, A.; Stacey, N.; Plaskitt, K.; Parr, A.; Chang, C.F.; Martin, C. Inhibition of phenolic acid metabolism results in precocious cell death and altered cell morphology in leaves of transgenic tobacco plants. Plant Cell 1998, 10, 1801–1816. [Google Scholar] [CrossRef] [Green Version]
- Macedo, E.S.; Sircar, D.; Cardoso, H.G.; Peixe, A.; Arnholdt-Schmitt, B. Involvement of alternative oxidase (AOX) in adventitious rooting of Olea europaea L. microshoots is linked to adaptive phenylpropanoid and lignin metabolism. Plant Cell Rep. 2012, 31, 1581–1590. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano, C.; Pallero-Baena, M.; Casimiro, I.; De Rybel, B.; Orman-Ligeza, B.; Van Isterdael, G.; Del Pozo, J.C. The emerging role of reactive oxygen species signaling during lateral root development. Plant Physiol. 2014, 165, 1105–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheniany, M.; Ebrahimzadeh, H.; Masoudi-nejad, A.; Vahdati, K.; Leslie, C. Effect of endogenous phenols and some antioxidant enzyme activities on rooting of Persian walnut (Juglans regia L.). Afr. J. Plant Sci. 2010, 4, 479–487. [Google Scholar]
- Schopfer, P. Hydroxyl radical-induced cell-wall loosening in vitro and in vivo: Implications for the control of elongation growth. Plant J. 2001, 28, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, A.; Ando, F.; Toyofuku, K.; Kawashima, C. Sucrose metabolism for the development of seminal root in maize seedlings. Plant Prod. Sci. 2009, 12, 9–16. [Google Scholar] [CrossRef]
- Vié, N.; Copois, V.; Bascoul-Mollevi, C.; Denis, V.; Bec, N.; Robert, B.; Gongora, C. Overexpression of phosphoserine aminotransferase PSAT1 stimulates cell growth and increases chemoresistance of colon cancer cells. Mol. Cancer 2008, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, C.G.; Berkowitz, O.; Hell, R.; Noji, M.; Saito, K. Characterization and expression analysis of a serine acetyltransferase gene family involved in a key step of the sulfur assimilation pathway in Arabidopsis. Plant Physiol. 2005, 137, 220–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Mochida, K.; Kato, T.; Tabata, S.; Yoshimoto, N.; Noji, M.; Saito, K. Comparative genomics and reverse genetics analysis reveal indispensable functions of the serine acetyltransferase gene family in Arabidopsis. Plant Cell 2008, 20, 2484–2496. [Google Scholar] [CrossRef] [Green Version]
- Rutter, W.J. Evolution of Aldolase; In Fed. Proceedings; November–December 1964; Volume 23, pp. 1248–1257. Available online: https://pubmed.ncbi.nlm.nih.gov/14236133/ (accessed on 16 March 2021).
- Cho, Y.-H.; Yoo, S.-D. Signaling role of fructose mediated by FINS1/FBP in Arabidopsis thaliana. PLoS Genet 2011, 7, e1001263. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.-J.; Shin, D.J.; Kim, B.G.; Byun, M.O. Putative fructose-1, 6-bisphosphate aldolase 1 (AtFBA1) affects stress tolerance in yeast and Arabidopsis. J. Plant Biotechnol. 2012, 39, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Webb, B.A.; Dosey, A.M.; Wittmann, T.; Kollman, J.M.; Barber, D.L. The glycolytic enzyme phosphofructokinase-1 assembles into filaments. J. Cell Biol. 2017, 216, 2305–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curci, P.L.; Cigliano, R.A.; Zuluaga, D.L.; Janni, M.; Sanseverino, W.; Sonnante, G. Transcriptomic response of durum wheat to nitrogen starvation. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kamada, T.; Nito, K.; Hayashi, H.; Mano, S.; Hayashi, M.; Nishimura, M. Functional differentiation of peroxisomes revealed by expression profiles of peroxisomal genes in Arabidopsis thaliana. Plant Cell Physiol. 2003, 44, 1275–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reumann, S.; Ma, C.; Lemke, S.; Babujee, L. AraPerox. A database of putative Arabidopsis proteins from plant peroxisomes. Plant Physiol. 2004, 136, 2587–2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massoumi, M.; Krens, F.A.; Visser, R.G.; De Klerk, G.J.M. Etiolation and flooding of donor plants enhance the capability of Arabidopsis explants to root. Plant Cell Tissue Organ Cult. 2017, 130, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Suárez, E.; Alfayate, C.; Pérez-Francés, J.F.; Rodríguez-Pérez, J.A. Structural and ultrastructural variations in in vitro and ex vitro rooting of microcuttings from two micropropagated Leucospermum (Proteaceae). Sci. Hortic. 2018, 239, 300–307. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Clean Reads (M) | Clean Bases (G) | Q20 (%) | Q30 (%) | GC (%) | Read Length (bp) | Total Mapped (%) | Splice Mapped (%) | Unique Mapped (%) | Multiple Mapped (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| BL-CK60-R1 | 41.6759 | 6.2514 | 96.95% | 91.80% | 42.63% | 150 | 86.44% | 30.60% | 81.60% | 4.84% |
| BL-CK60-R2 | 40.7821 | 6.1275 | 97.54% | 93.07% | 42.20% | 150 | 87.42% | 30.56% | 82.88% | 4.54% |
| BL-CK60-R3 | 39.9473 | 5.9921 | 96.85% | 91.62% | 42.95% | 150 | 86.87% | 32.51% | 82.23% | 4.64% |
| BL-60-R1 | 46.7699 | 7.0155 | 96.82% | 91.56% | 42.38% | 150 | 85.98% | 30.32% | 81.73% | 4.24% |
| BL-60-R2 | 45.0371 | 6.7556 | 96.68% | 91.25% | 42.39% | 150 | 84.91% | 28.39% | 80.41% | 4.50% |
| BL-60-R3 | 59.2106 | 8.8816 | 97.78% | 93.59% | 42.35% | 150 | 84.75% | 26.66% | 80.59% | 4.16% |
| BL-66-R1 | 38.8345 | 5.8252 | 96.85% | 91.55% | 42.43% | 150 | 86.52% | 29.99% | 81.83% | 4.69% |
| BL-66-R2 | 41.0296 | 6.1544 | 96.97% | 91.81% | 42.11% | 150 | 86.20% | 28.55% | 81.68% | 4.51% |
| BL-66-R3 | 41.4342 | 6.2151 | 96.88% | 91.62% | 42.46% | 150 | 87.36% | 31.33% | 82.67% | 4.69% |
| BL-72-R1 | 57.8766 | 8.6815 | 97.34% | 92.74% | 42.23% | 150 | 82.91% | 26.06% | 78.76% | 4.15% |
| BL-72-R2 | 47.1213 | 7.0682 | 96.87% | 91.69% | 42.30% | 150 | 83.40% | 25.65% | 79.24% | 4.17% |
| BL-72-R3 | 47.6405 | 7.1461 | 96.79% | 91.47% | 42.43% | 150 | 84.22% | 27.69% | 79.91% | 4.31% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munir, M.Z.; Ud Din, S.; Imran, M.; Zhang, Z.; Pervaiz, T.; Han, C.; Un Nisa, Z.; Bakhsh, A.; Atif Muneer, M.; Sun, Y.; et al. Transcriptomic and Anatomic Profiling Reveal Etiolation Promotes Adventitious Rooting by Exogenous Application of 1-Naphthalene Acetic Acid in Robinia pseudoacacia L. Forests 2021, 12, 789. https://doi.org/10.3390/f12060789
Munir MZ, Ud Din S, Imran M, Zhang Z, Pervaiz T, Han C, Un Nisa Z, Bakhsh A, Atif Muneer M, Sun Y, et al. Transcriptomic and Anatomic Profiling Reveal Etiolation Promotes Adventitious Rooting by Exogenous Application of 1-Naphthalene Acetic Acid in Robinia pseudoacacia L. Forests. 2021; 12(6):789. https://doi.org/10.3390/f12060789
Chicago/Turabian StyleMunir, Muhammad Zeeshan, Saleem Ud Din, Muhammad Imran, Zijie Zhang, Tariq Pervaiz, Chao Han, Zaib Un Nisa, Ali Bakhsh, Muhammad Atif Muneer, Yuhan Sun, and et al. 2021. "Transcriptomic and Anatomic Profiling Reveal Etiolation Promotes Adventitious Rooting by Exogenous Application of 1-Naphthalene Acetic Acid in Robinia pseudoacacia L." Forests 12, no. 6: 789. https://doi.org/10.3390/f12060789