
Influence of Slope Direction on the Soil Seed Bank and Seedling Regeneration of Castanopsis hystrix Seed Rain

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Research Area
2.2. Setting of Sample Plots
2.3. Investigation of Castanopsis Hystrix Seed Rain
2.4. Investigation of Castanopsis Hystrix Soil Seed Bank
2.5. Investigation of 1-Year-Old Castanopsis Hystrix Seedlings
2.6. Data Processing
3. Results
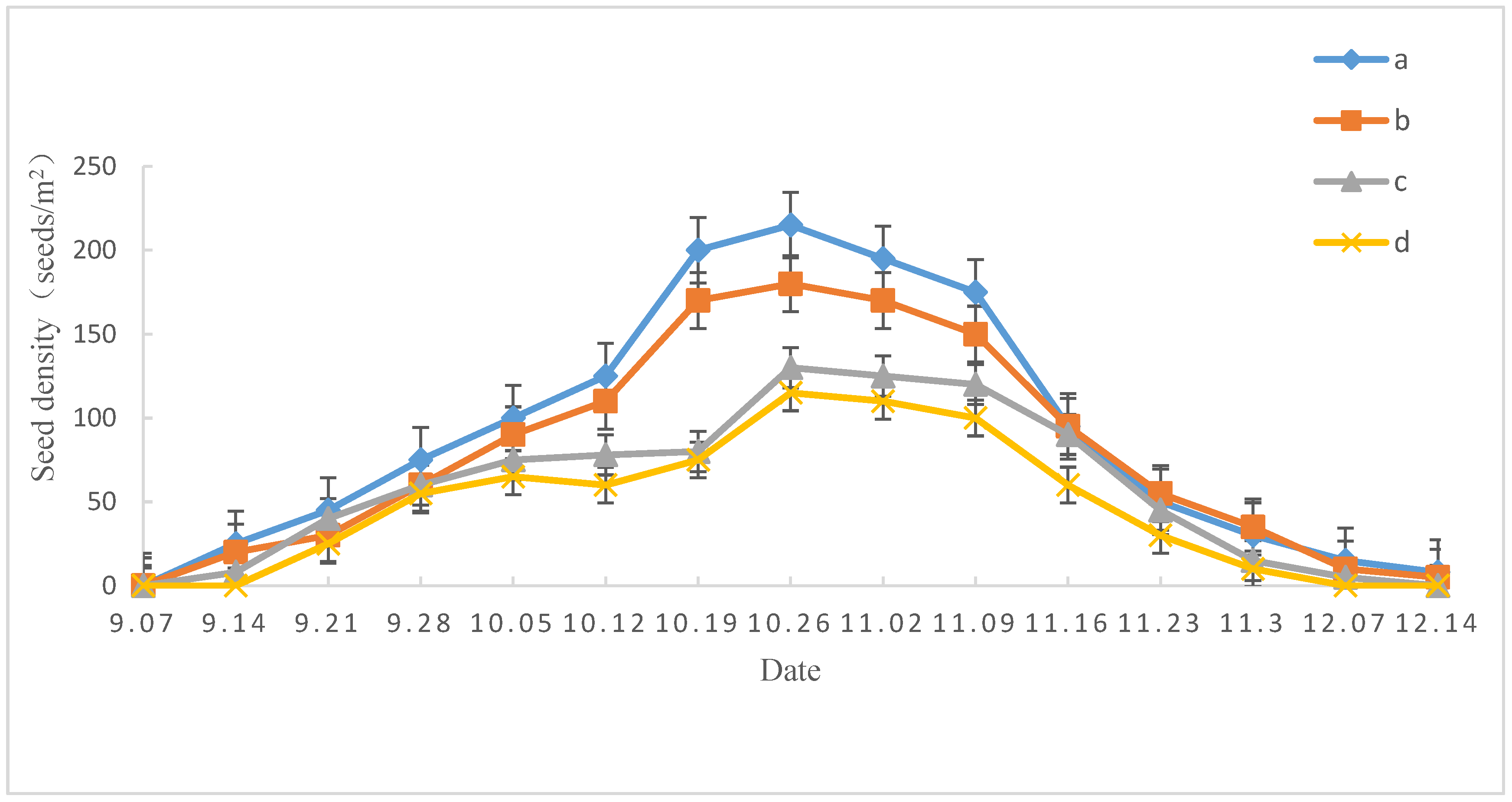
3.1. Characteristics of the Castanopsis Hystrix Seed Rain
3.1.1. Temporal Dynamics of Seed Rain Diffusion
3.1.2. Composition and Quality of Seed Rain
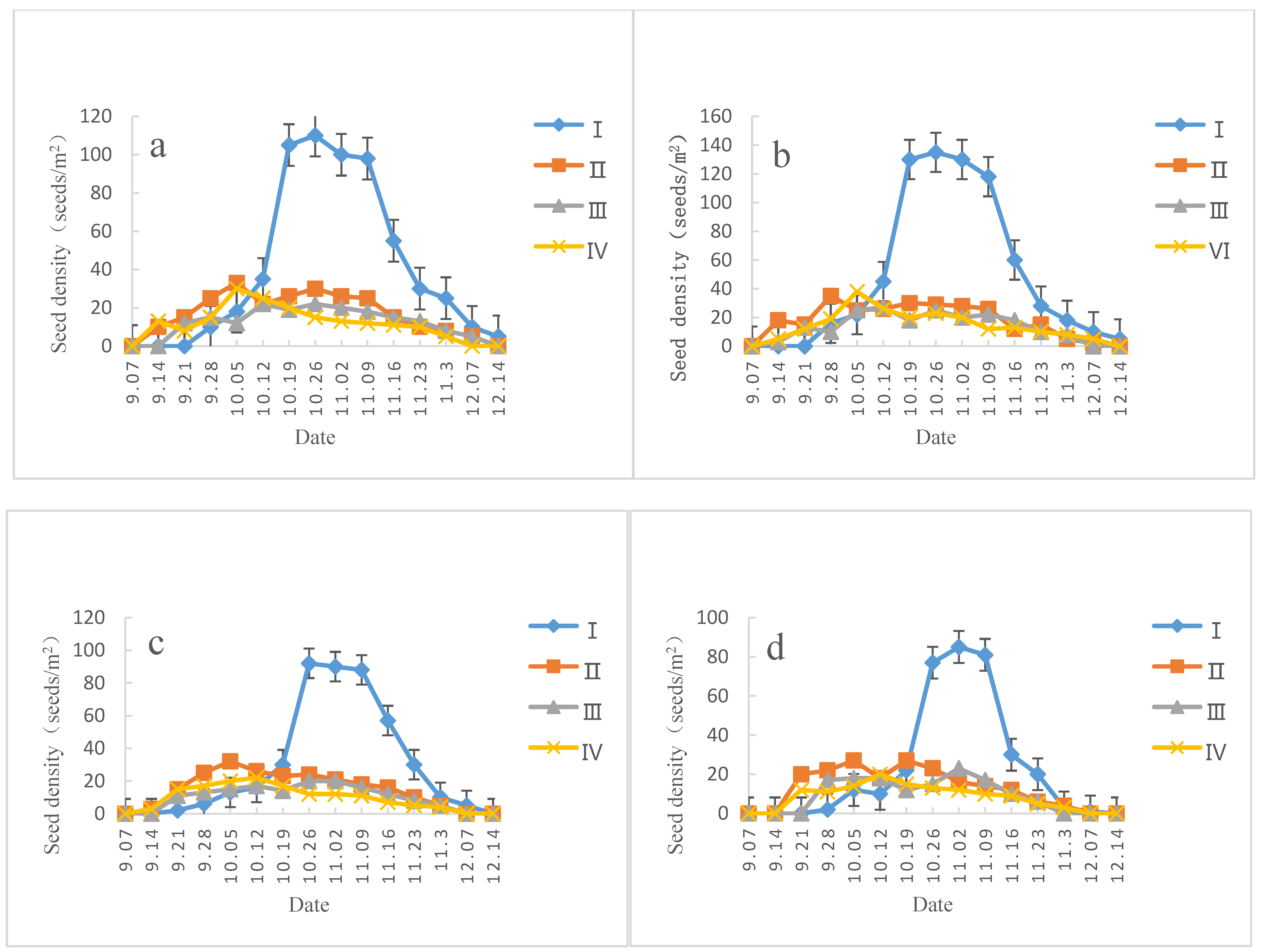
3.2. Characteristics of the Castanopsis Hystrix Soil Seed Bank
3.2.1. Reserves and Dynamics of the Soil Seed Bank
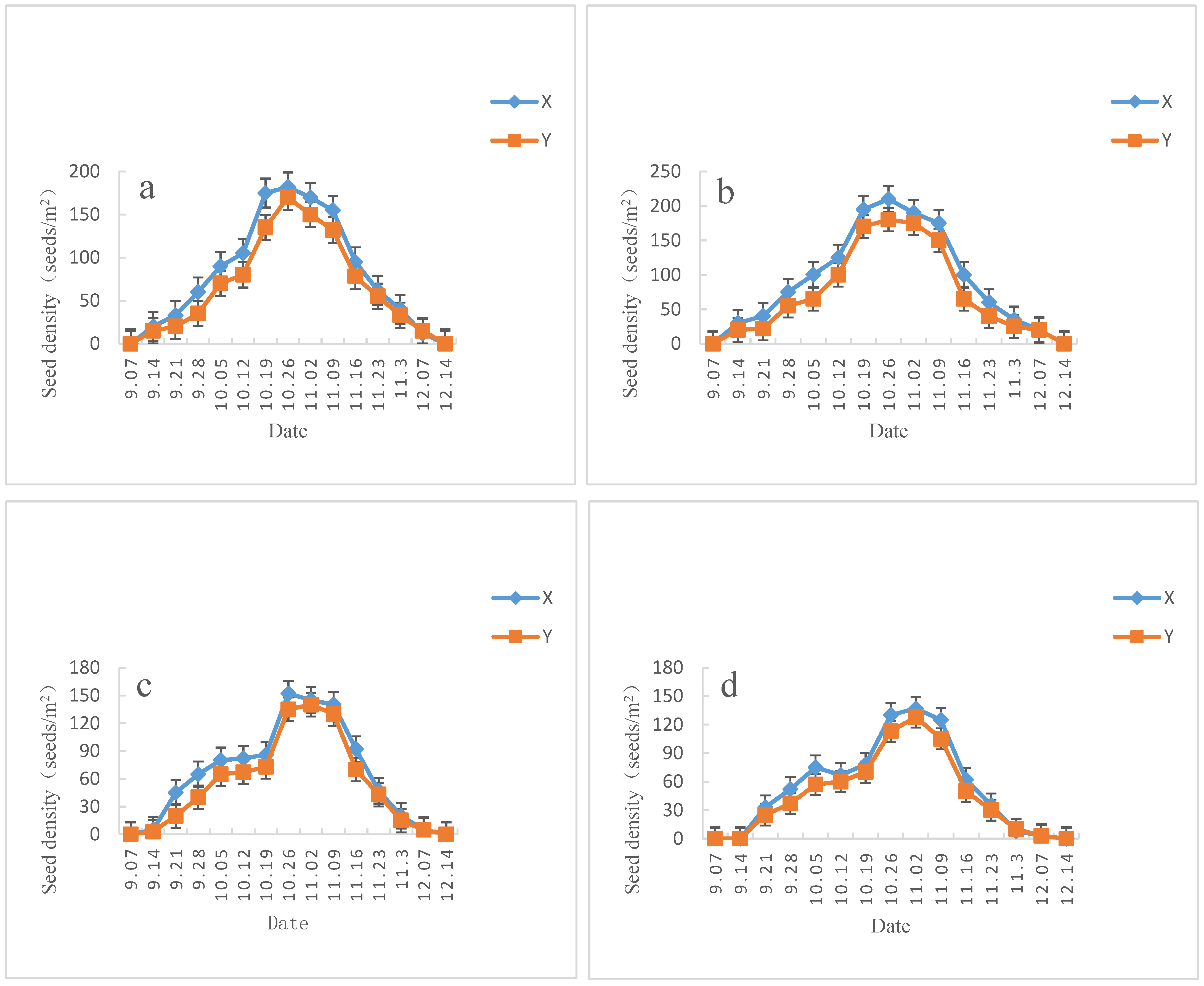
3.2.2. Vertical Distribution of Soil Seed Bank
3.3. Investigation of the Seedlings of Castanopsis Hystrix
4. Discussions
4.1. Characteristics of Castanopsis Hystrix Seed Rain
4.2. Dynamics of the Castanopsis Hystrix Soil Seed Bank
4.3. Regeneration of Castanopsis Hystrix Seedlings
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nathan, R.; Helene, C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 2000, 15, 278–285. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977; pp. 256–263. [Google Scholar]
- Li, X.S.; Peng, M.C.; Dang, C.L. Research progress on natural regeneration of plants. Chin. J. Ecol. 2007, 26, 2081–2088. [Google Scholar]
- Heredia, U.L.D.; Nanos, N.; Garcíadelrey, E.; Guzmán, P.; Gil, L. High seed dispersal ability of Pinus canariensis in stands of contrasting density inferred from genotypic data. For. Syst. 2015, 24, e015. [Google Scholar]
- Benigno, G.R.; Mulualem, T.; Guillermo, C.M. Soil seed bank assembly following secondary succession on abandoned agricultural fields in Nicaragua. J. For. Res. 2009, 20, 349–354. [Google Scholar]
- Simpson, R.L. Ecology of Soil Seed Bank; Academic Press: San Diego, CA, USA, 1989; pp. 149–209. [Google Scholar]
- Lu, Y.L.; Zhang, W.H.; Yang, B.; Wang, Y.N. Seed rain, soil seed bank and seedling regeneration of Quercus aliena var. acureserrata in different slope directions on the middle Qinling Mountains, China. Chin. J. Appl. Ecol. 2019, 30, 1–11. [Google Scholar]
- Liu, Z.G.; Zhu, J.J.; Yuan, X.L.; Wang, H.X.; Tan, H. On seed rain soil seed bank of Larix olgensis in montane regions of eastern Liaoning Province, China. Acta Ecol. Sin. 2007, 27, 579–587. [Google Scholar]
- Eriksson, O. Evolution of seed dispersal and recruitment in clonal plants. Oikos 1992, 63, 439–448. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, Y.; Tian, Z.W.; Jia, H.Y.; Zhao, R.R.; An, N. Dynamics of seed rain, soil seed bank and seedling regeneration of Castanopsis hystrix. Sci. Silvae Sin. 2020, 56, 37–49. [Google Scholar]
- Chang, J.G.; Liu, S.R.; Shi, Z.M.; Kang, B.; Chen, B.Y. Response of soil respiration to soil temperature and moisture regimes in the natural forest of Quercus aliena var. acuteserrata. Sci. Silvae Sin. 2006, 42, 21–27. [Google Scholar]
- Liu, G.Q.; Ni, W.J.; Liu, H.F.; Gravenhorst, G. Eco-environment and nutrient accumulation of sharptooth oak stands in southern slope of Mt. Qinling. Chin. J. Appl. Ecol. 2002, 13, 513–518. [Google Scholar]
- Li, Z.; Dong, N.N.; Hou, L.; Zhang, F.; Ma, W.G. Characteristics of soil and litter organic carbon and total nitrogen in different age groups of Quercus aliena var. acuteserrata forests in the Qinling Mountains. J. Cent. South Univ. For. Technol. 2017, 37, 127–132. [Google Scholar]
- Liu, C.; Sun, P.S.; Liu, S.R.; Lu, H.B.; Chen, Z.C.; Liu, X.J. Leaf photosynthetic pigment seasonal dynamic of Quercus aliena var. acuteserrata and its spectral reflectance response under throughfall elimination. Chin. J. Appl. Ecol. 2017, 28, 1077–1086. [Google Scholar]
- Chen, Z.C.; Liu, C.; Liu, X.J.; Wan, X.C. Effects of irradiation and tree size on the carbon-water balances of Quercus aliena var. acuteserrata. Sci. Silvae Sin. 2017, 53, 18–25. [Google Scholar]
- Zhang, W.H.; Lu, Y.C.; Zhou, J.Y.; Zhang, X.H.; Shi, X.H. Population structure and dynamics of Quercus variabilis in different habitats on northern slope of Bashan Mountain. Sci. Silvae Sin. 2008, 44, 12–15. [Google Scholar]
- Yin, H.J.; Cheng, X.Y.; Lai, T.; Lin, B.; Liu, Q. Seed rain, soil seed bank and seedling regeneration in a 65-year Picea asperata plantation in subalpine coniferous, western Sichuan, China. Chin. J. Plant Ecol. 2011, 35, 35–44. [Google Scholar] [CrossRef]
- Tanouchi, H.; Sato, T.; Takeshita, K. Comparative studies on acorn and seedling dynamics of four Quercus species in evergreen broad-leaved forest. J. Plant Res. 1994, 107, 153–159. [Google Scholar] [CrossRef]
- Wang, L.Y.; Liu, G.Z.; Yue, J.W.; Long, W.; Yang, H.; Jin, H.P. Seed rain and soil seed bank of typical evergreen broad-leaved forests in Jiangxi Province. China For. Sci. Technol. 2015, 29, 37–43. [Google Scholar]
- Willson, M.F. Dispersal mode, seed shadows, and colonization patterns. Plant Ecol. 1993, 107, 261–280. [Google Scholar]
- Liu, T.; Zhou, Z.Q. Seed rain and soil seed bank of Quercus mongolica populations. J. Northeast. For. Univ. 2007, 35, 22–23. [Google Scholar]
- Wang, W.; Ma, K.P.; Liu, C.R. Seed shadow of Quercus liaotungensis in a broad leaved forest in Dongling Mountain. Acta Bot. Sin. 2000, 42, 195–202. [Google Scholar]
- Horton, J.S.; Wright, J.T. The wood rat as an ecological factor in southern California watersheds. Ecology 1944, 25, 341–351. [Google Scholar] [CrossRef]
- Tian, R.S. The influence of site condition on the growth of Larix principis-rupprechtii. Mod. Hortic. 2018, 19, 76–77. [Google Scholar]
- Watt, A.S. On the causes of failure of natural regeneration in British oak woods. J. Ecol. 1919, 7, 173–203. [Google Scholar] [CrossRef]
- Liu, G.X.; Mao, P.S.; Wang, Y.W.; Han, J.G. Effects of adult neighbour and gap size on seedling emergence and earth growth of Bromus inermis Leyss. Ecol. Res. 2008, 23, 197–205. [Google Scholar] [CrossRef]
- Koutecká, E.; Lep, J. Effect of light and moisture conditions and seed age on germination of three closely related Myosotis species. Folia Geobot. 2009, 44, 109–130. [Google Scholar] [CrossRef]
- Zhan, X.; Li, L.; Cheng, W. Restoration of Stipa krylovii steppes in Inner Mongolia of China: Assessment of seed banks and vegetation composition. J. Arid Environ. 2007, 68, 298–307. [Google Scholar] [CrossRef]
- Zhou, Z.Q.; Bao, W.K.; Wu, F.Z.; Wu, N. Capability and limitation of regeneration of Rosa hugonis and Rosa soulieana in the dry valley of the upper Minjiang River. Acta Ecol. Sin. 2009, 29, 1931–1939. [Google Scholar]
- Garcia, D.; Obseo, J.R.; Martinez, I. Spatial concordance between seed rain and seedling establishment in bird-dispersed trees: Does scale matter. J. Ecol. 2005, 93, 693–704. [Google Scholar] [CrossRef]
- Zhang, X.B.; Wang, R.J.; Shangguan, Z.P. Dynamics of seed rain and soil seed bank in Pinus tabuliformis Carr. Forests in eroded hilly loess regions of the Loess Plateau in China. Acta Ecol. Sin. 2009, 29, 1877–1884. [Google Scholar]
- Crawley, M.J.; Long, C.R. Alternate bearing, predator satiation and seedling recruitment in Quercus robur L. J. Ecol. 1995, 83, 683–696. [Google Scholar] [CrossRef]
- Du, X.J.; Guo, Q.F.; Gao, X.M.; Ma, K.P. Seed rain, soil seed bank, seed loss and regeneration of Castanopsis fargesii(Fagaceae) in a subtropical evergreen broad-leaved forest. For. Ecol. Manag. 2007, 238, 212–219. [Google Scholar] [CrossRef]
- Zhang, X.L.; Xu, J.W.; Shen, H.L.; Huang, J.; Zhang, P. Animal predation and dispersal of Sorbus pohuashanensis fruits and seeds. Chin. J. Appl. Ecol. 2010, 21, 2677–2683. [Google Scholar]
- Cai, L.J.; Liu, F.G.; Xiao, W.M.; Zhao, A.J. Spatial distribution characteristics of vegetation coverage in Renshou County. Sichuan For. Explor. Des. 2017, 2, 50–56. [Google Scholar]
- Wang, Z.L.; Gao, X.M. The predating behavior of rodents and its effects to acorns of Quercus aliena var. acuteserrata. Acta Ecol. Sin. 2006, 26, 3533–3541. [Google Scholar] [CrossRef]
- Csontos, P. Seed banks: Ecological definitions and sampling considerations. Community Ecol. 2007, 8, 75–85. [Google Scholar] [CrossRef]
- Gao, R.M.; Shi, X.D.; Guo, Y.D.; Fan, L.Y. Seed rain, soil seed bank and regeneration of Larix principis-rupprechtii stands in the Upper Reaches of Wenyuhe. Acta Ecol. Sin. 2015, 35, 3588–3597. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Index | Soil Depth/cm | Sunny Slope | Semi-Sunny Slope | Semi-Shady Slope | Shady Slope |
|---|---|---|---|---|---|
| 0–20 | 66.91 ± 13.30 Aa | 70.48 ± 11.37 Aa | 45.95 ± 4.08 Ab | 47.23 ± 5.13 Ab | |
| Organic content/g∙kg−1 | 20–40 | 42.32 ± 2.75 Ba | 45.38 ± 7.36 Ba | 38.20 ± 8.82 Aa | 37.12 ± 6.14 Aa |
| 40–60 | 32.35 ± 5.87 Ba | 33.73 ± 4.72 Ba | 24.22 ± 5.68 Bb | 24.84 ± 4.17 Bb | |
| 0–20 | 1.32 ± 0.11 Aa | 1.96 ± 0.33 Ab | 1.15 ± 0.46 Aa | 1.18 ± 0.77 Aa | |
| Total nitrogen content/g∙kg−1 | 20–40 | 1.02 ± 0.13 Aa | 0.91 ± 0.07 Ba | 0.70 ± 0.03 Bb | 0.66 ± 0.09 Bb |
| 40–60 | 0.76 ± 0.24 Aa | 0.84 ± 0.12 Ba | 0.69 ± 0.02 Ba | 0.65 ± 0.01 Ba | |
| 0–20 | 0.37 ± 0.03 Aab | 0.42 ± 0.08 Ab | 0.27 ± 0.01 Aa | 0.24 ± 0.05 Aa | |
| Total phosphorus content/g∙kg−1 | 20–40 | 0.26 ± 0.03 Ba | 0.38 ± 0.06 Ab | 0.20 ± 0.02 Ba | 0.19 ± 0.01 Ba |
| 40–60 | 0.20 ± 0.04 Ca | 0.32 ± 0.03 Ab | 0.15 ± 0.02 Ba | 0.17 ± 0.03 Ba | |
| 0–20 | 5.68 ± 0.26 Aa | 5.87 ± 0.19 Aa | 3.92 ± 0.32 Ab | 3.80 ± 0.45 Ab | |
| Total potassium content/g∙kg−1 | 20–40 | 5.01 ± 0.14 Ba | 5.06 ± 0.25 Bc | 3.46 ± 0.13 ABb | 3.40 ± 0.26 Bb |
| 40–60 | 4.45 ± 0.07 Ca | 4.52 ± 0.63 Bb | 3.03 ± 0.30 Bb | 3.00 ± 0.51 Bb | |
| 0–20 | 37.90 ± 11.39 Aa | 25.39 ± 5.90 Ab | 19.16 ± 2.56 Ab | 17.25 ± 3.17 Ab | |
| C/N | 20–40 | 25.81 ± 4.36 Aa | 28.46 ± 6.37 Aa | 29.80 ± 7.81 Aa | 26.91 ± 4.66 Aa |
| 40–60 | 27.62 ± 5.75 ABa | 23.22 ± 3.25 Aa | 21.07 ± 6.20 Aa | 20.20 ± 4.18 Aa | |
| 0–20 | 4.43 ± 0.09 Aa | 4.98 ± 0.86 Aa | 4.10 ± 0.08 Ab | 4.06 ± 0.09 Ab | |
| pH | 20–40 | 4.65 ± 0.08 Aa | 4.50 ± 0.07 Aa | 4.52 ± 0.12 Ba | 4.46 ± 0.08 Ba |
| 40–60 | 4.77 ± 0.18 Aa | 4.70 ± 0.15 Aa | 4.59 ± 0.05 Ba | 4.50 ± 0.03 Ba |
| Gradient/° | Slope Position | Slope Direction | Average Height of Trees/m | Average DBH/cm | Depth of Litter Layer/cm | Soil Moisture/% | Canopy Density/% |
|---|---|---|---|---|---|---|---|
| 28–33 | Middle | Sunny slope | 28.0 | 40.6 | 3.8 | 42.2 | 80 |
| 31–35 | Middle | Semi-sunny slope | 27.3 | 38.7 | 4.3 | 45.3 | 83 |
| 29–36 | Middle | Semi-shady slope | 26.8 | 37.0 | 4.6 | 47.5 | 75 |
| 15–20 | Down | Shady slope | 25.1 | 36.2 | 5.0 | 49.7 | 71 |
| Slope Direction | Total Amount of Seed Rain (Seeds∙m−2) | Amount of Mature Seeds (Seeds∙m−2) | Amount of Immature Seeds (Seeds∙m−2) | Amount of Insect-Damaged Seeds (Seeds∙m−2) | Amount of Gnawed Seeds (Seeds∙m−2) | Seed Vigor (%) |
|---|---|---|---|---|---|---|
| Sunny slope | 457.80 ± 10.05 b | 238.04 ± 5.48 b | 95.80 ± 3.98 b | 63.40 ± 2.12 a | 60.56 ± 1.84 a | 49.6 ± 3.8 ab |
| Semi-sunny slope | 516.52 ± 10.46 a | 284.32 ± 6.40 a | 103.16 ± 4.20 a | 64.64 ± 2.04 a | 64.40 ± 2.26 a | 51.0 ± 4.2 a |
| Semi-shady slope | 359.00 ± 8.09 c | 172.52 ± 3.72 c | 84.72 ± 3.66 c | 51.92 ± 1.04 b | 49.84 ± 1.72 b | 43.5 ± 3.8 b |
| Shady slope | 310.44 ± 6.31 d | 129.24 ± 2.60 d | 74.44 ± 2.72 d | 55.64 ± 1.50 b | 51.12 ± 1.62 b | 34.1 ± 3.2 c |
| Slope Direction | Date | Seed Bank Density (Seeds∙m−2) | Amount of Mature Seeds (Seeds∙m−2) | Vigor (%) | Amount of Immature Seeds (Seeds∙m−2) | Amount of Insect-Damaged Seeds (Seeds∙m−2) | Amount of Gnawed Seeds (Seeds∙m−2) | Amount of Moldy Seeds (Seeds∙m−2) |
|---|---|---|---|---|---|---|---|---|
| Sunny slope | 2017-12 | 272.80 ± 8.34 a | 136.88 ± 4.62 a | 48.4 ± 2.6 a | 71.16 ± 4.14 a | 28.68 ± 1.36 a | 27.40 ± 1.34 a | 8.68 ± 0.64 d |
| 2018-04 | 143.64 ± 4.84 b | 62.56 ± 2.50 b | 17.2 ± 1.2 b | 21.28 ± 1.12 b | 18.76 ± 1.14 b | 21.52 ± 1.02 a | 19.52 ± 1.00 c | |
| 2018-06 | 86.28 ± 3.36 c | 15.64 ± 0.88 c | 3.9 ± 0.3 c | 19.24 ± 1.02 b | 16.28 ± 1.04 b | 9.20 ± 0.56 b | 25.92 ± 1.32 b | |
| 2018-08 | 64.20 ± 1.96 c | 5.76 ± 0.34 c | 0.0 ± 0.0 c | 10.56 ± 1.32 c | 11.36 ± 0.70 b | 4.88 ± 0.34 b | 31.64 ± 1.64 a | |
| Semi-sunny slope | 2017-12 | 308.68 ± 8.46 a | 163.40 ± 4.40 a | 52.6 ± 2.6 a | 75.04 ± 4.02 a | 29.88 ± 1.48 a | 29.32 ± 1.44 a | 11.04 ± 0.70 d |
| 2018-04 | 151.24 ± 5.00 b | 73.20 ± 2.72 b | 18.8 ± 1.4 b | 22.52 ± 1.36 b | 21.04 ± 1.10 b | 17.80 ± 1.18 b | 16.68 ± 1.02 c | |
| 2018-06 | 93.68 ± 4.04 c | 18.32 ± 0.98 c | 4.3 ± 0.4 c | 14.88 ± 0.78 c | 18.92 ± 0.96 b | 14.00 ± 0.72 c | 27.56 ± 1.38 b | |
| 2018-08 | 65.76 ± 2.82 d | 7.48 ± 0.20 c | 0.0 ± 0.0 c | 8.44 ± 0.56 d | 8.84 ± 0.48 c | 7.24 ± 0.36 d | 33.76 ± 1.92 a | |
| Semi-shady slope | 2017-12 | 239.36 ± 7.18 a | 111.60 ± 4.72 a | 39.3 ± 2.4 a | 72.08 ± 4.10 a | 25.40 ± 1.16 a | 22.52 ± 1.04 a | 7.76 ± 0.54 d |
| 2018-04 | 87.20 ± 3.78 b | 27.08 ± 1.74 b | 12.7 ± 1.1 b | 15.40 ± 1.00 b | 22.80 ± 1.06 a | 8.88 ± 0.48 b | 13.04 ± 0.68 c | |
| 2018-06 | 61.20 ± 2.66 c | 8.64 ± 0.44 c | 3.3 ± 0.3 c | 6.64 ± 0.54 c | 16.92 ± 1.08 b | 6.32 ± 0.32 b | 22.68 ± 0.98 b | |
| 2018-08 | 49.40 ± 2.08 c | 2.56 ± 0.14 c | 0.0 ± 0.0 c | 5.60 ± 0.46 c | 7.20 ± 0.40 c | 5.08 ± 0.24 b | 28.96 ± 1.56 a | |
| Shady slope | 2017-12 | 171.16 ± 5.62 a | 69.28 ± 3.52 a | 31.2 ± 2.2 a | 48.44 ± 1.62 a | 18.20 ± 1.12 a | 26.28 ± 1.24 a | 8.96 ± 0.56 d |
| 2018-04 | 83.16 ± 3.44 b | 24.56 ± 1.08 b | 10.5 ± 1.0 b | 15.88 ± 0.98 b | 7.72 ± 0.40 b | 15.24 ± 0.66 b | 19.76 ± 0.90 c | |
| 2018-06 | 57.56 ± 2.16 c | 8.92 ± 0.56 c | 2.5 ± 0.2 c | 6.76 ± 0.56 c | 5.92 ± 0.32 b | 6.28 ± 0.52 c | 29.68 ± 1.14 b | |
| 2018-08 | 45.44 ± 2.20 c | 0.64 ± 0.04 d | 0.0 ± 0.0 c | 2.16 ± 0.16 c | 3.48 ± 0.26 b | 4.84 ± 0.34 c | 34.32 ± 1.46 a |
| Slope Direction | Date | Litter Layer | 0–2 cm Humus Layer | 2–5 cm Soil Layer |
|---|---|---|---|---|
| Sunny slope | 2017-12 | 133.86 ± 7.60 a | 5.08 ± 0.66 b | 0.0 ± 0.0 c |
| 2018-04 | 65.38 ± 4.47 b | 12.80 ± 1.26 a | 0.0 ± 0.0 c | |
| 2018-06 | 35.25 ± 3.16 c | 15.18 ± 1.45 a | 0.21 ± 0.02 b | |
| 2018-08 | 23.64 ± 1.73 c | 16.93 ± 1.83 a | 0.88 ± 0.12 a | |
| Semi-sunny slope | 2017-12 | 143.60 ± 8.06 a | 9.38 ± 1.12 b | 0.0 ± 0.0 c |
| 2018-04 | 67.20 ± 4.22 b | 16.80 ± 1.68 a | 0.0 ± 0.0 c | |
| 2018-06 | 37.50 ± 3.70 c | 17.56 ± 1.71 a | 1.25 ± 0.09 b | |
| 2018-08 | 22.18 ± 2.11 c | 19.92 ± 1.92 a | 1.60 ± 0.18 a | |
| Semi-shady slope | 2017-12 | 116.93 ± 6.60 a | 7.48 ± 0.42 b | 0.0 ± 0.0 c |
| 2018-04 | 36.78 ± 3.70 b | 13.88 ± 1.36 a | 0.0 ± 0.0 c | |
| 2018-06 | 22.86 ± 2.46 c | 15.20 ± 1.65 a | 1.05 ± 0.12 b | |
| 2018-08 | 15.66 ± 1.33 c | 16.52 ± 1.84 a | 1.48 ± 0.16 a | |
| Shady slope | 2017-12 | 85.36 ± 5.38 a | 6.16 ± 0.68 b | 0.0 ± 0.0 c |
| 2018-04 | 35.46 ± 3.30 b | 12.49 ± 1.20 a | 0.0 ± 0.0 c | |
| 2018-06 | 21.30 ± 1.83 c | 13.21 ± 1.35 a | 1.80 ± 0.16 b | |
| 2018-08 | 13.82 ± 1.28 c | 15.48 ± 1.57 a | 2.25 ± 0.28 a |
| Slope Direction | Density of Regenerated Seedlings (Plants∙m−2) | Density of Surviving Seedlings (Plants∙m−2) | Survival Rate (%) |
|---|---|---|---|
| Sunny slope | 29.73 ± 3.31 a | 13.60 ± 1.74 b | 45.75 ± 3.15 b |
| Semi-sunny slope | 37.10 ± 4.55 a | 19.65 ± 2.62 a | 52.96 ± 3.06 a |
| Semi-shady slope | 16.52 ± 2.40 b | 6.80 ± 0.90 bc | 41.16 ± 4.37 b |
| Shady slope | 8.33 ± 1.42 b | 2.37 ± 0.57 c | 28.45 ± 4.58 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Liu, Y.; Jia, H.; Sun, W.; Ming, A.; Pang, S.; An, N.; Zhang, J.; Tang, C.; Dong, S. Influence of Slope Direction on the Soil Seed Bank and Seedling Regeneration of Castanopsis hystrix Seed Rain. Forests 2021, 12, 500. https://doi.org/10.3390/f12040500
Zhao Z, Liu Y, Jia H, Sun W, Ming A, Pang S, An N, Zhang J, Tang C, Dong S. Influence of Slope Direction on the Soil Seed Bank and Seedling Regeneration of Castanopsis hystrix Seed Rain. Forests. 2021; 12(4):500. https://doi.org/10.3390/f12040500
Chicago/Turabian StyleZhao, Zong, Yong Liu, Hongyan Jia, Wensheng Sun, Angang Ming, Shengjiang Pang, Ning An, Jihui Zhang, Chuang Tang, and Shitao Dong. 2021. "Influence of Slope Direction on the Soil Seed Bank and Seedling Regeneration of Castanopsis hystrix Seed Rain" Forests 12, no. 4: 500. https://doi.org/10.3390/f12040500