
Rapid Changes in Ground Vegetation of Mature Boreal Forests—An Analysis of Swedish National Forest Inventory Data

 , , and
, , and
Abstract
:1. Introduction
- Identify if directional changes in percentage cover of species and species groups have occurred in old/mature forests that have not been subject to forest management during a 10-year period.
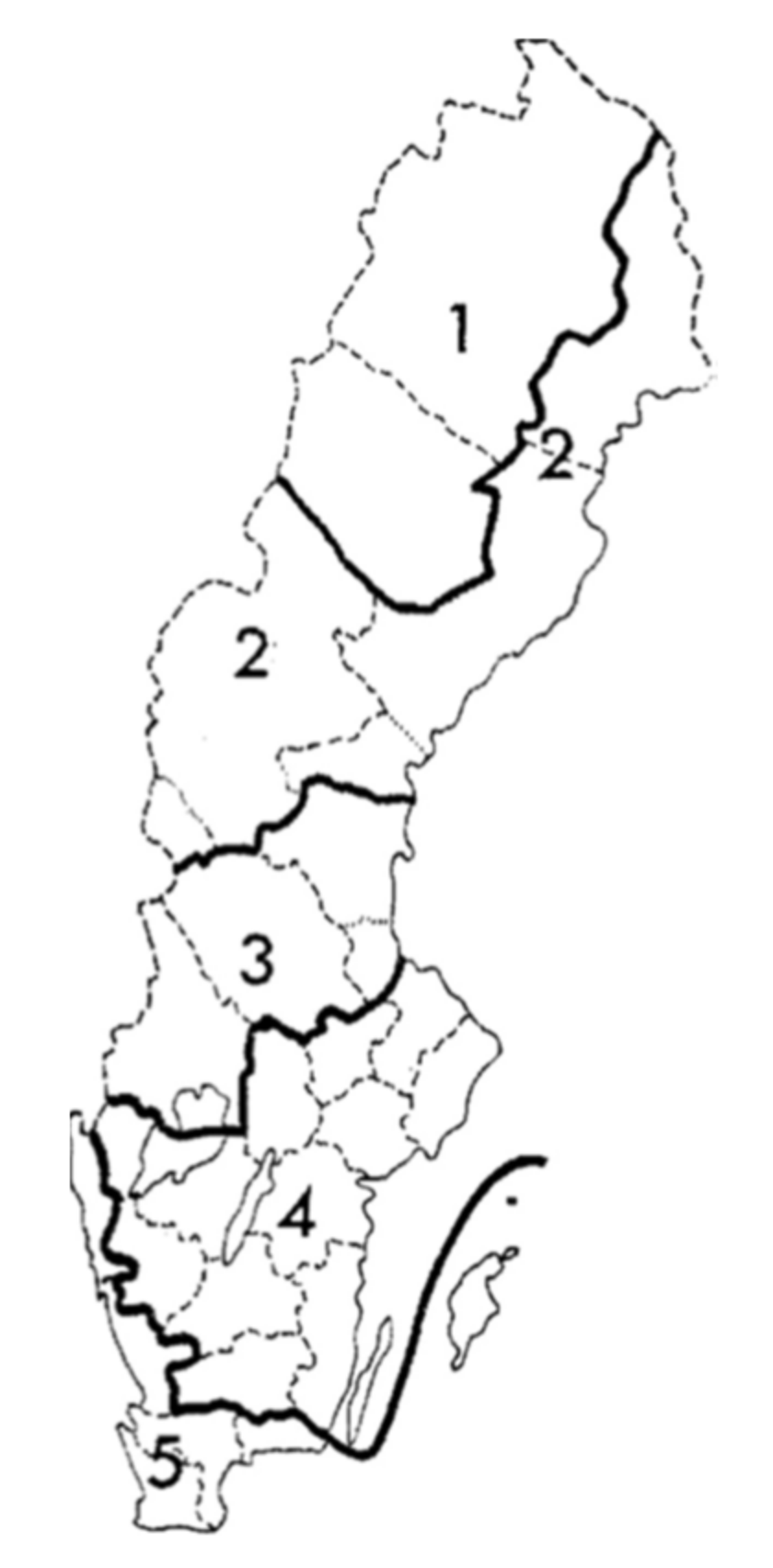
- If such changes are present, to analyse if the direction of change varies among four regions in Sweden, stretching from hemiboreal to northern boreal regions.
- To explore factors that might have contributed to the identified changes.
2. Materials and Methods
2.1. Study Area
2.2. NFI Data
2.3. Statistical Analysis
3. Results
3.1. Stand Structure
3.2. Change in Species Abundance
3.3. Factors Correlating with Change in Cover
3.4. Change in Species Groups
4. Discussion
4.1. Succession
4.2. Climate
4.3. Nitrogen Deposition
4.4. Time Lags in Responses
4.5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Nomenclature
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Species | Stand Productivity | Basal Area Change | Thinning Prior First Inventory | Maturity Class | N |
|---|---|---|---|---|---|
| Graminoids | |||||
| Broad leaved grasses | 0.044 | <0.001 | 0.066 | 0.160 | 482 |
| Narrow leaved grasses | 0.613 | 0.431 | 0.397 | 0.204 | 953 |
| Carex globularis | 0.606 | 0.686 | 0.014 | 0.527 | 240 |
| Herbs | |||||
| Anemone nemorosa | 0.429 | 0.036 | 0.538 | 0.048 | 195 |
| Maianthemum bifolium | 0.111 | 0.963 | 0.299 | 0.067 | 470 |
| Melampyrum spp. | 0.718 | 0.099 | 0.010 | 0.368 | 762 |
| Oxalis acetosella | 0.325 | 0.844 | 0.287 | 0.020 | 259 |
| Dwarf shrubs | |||||
| Calluna vulgaris | 0.081 | 0.482 | <0.001 | 0.320 | 442 |
| Empetrum nigrum | 0.698 | 0.346 | 0.854 | 0.866 | 486 |
| Rubus chamaemorus | 0.634 | 0.291 | 0.447 | 0.334 | 172 |
| Rhododendron tomentosum | 0.857 | 0.570 | 0.066 | 0.595 | 133 |
| Vaccinium myrtillus | 0.459 | 0.318 | 0.004 | 0.018 | 1230 |
| Vaccinium uliginosum | 0.374 | 0.233 | 0.467 | 0.434 | 325 |
| Vaccinium vitis-idae | 0.777 | 0.633 | 0.300 | 0.232 | 1143 |
| Spore plants | |||||
| Equisetum sylvaticum | 0.482 | 0.258 | 0.347 | 0.503 | 236 |
| Gymnocarpium dryopteris | 0.435 | 0.351 | 0.828 | 0.234 | 222 |
| Lycopodiaceae | 0.818 | 0.552 | 0.271 | 0.037 | 278 |
| Tall ferns | 0.096 | 0.528 | 0.611 | 0.474 | 124 |
| Lichens | |||||
| Cladonia spp. excl. Cladina | 0.151 | 0.46 | 0.704 | 0.570 | 799 |
| Cladonia grp. Cladina | <0.001 | 0.608 | 0.401 | <0.001 | 607 |
| Other ground lichens | 0.003 | 0.623 | 0.572 | 0.443 | 626 |
| Mosses | |||||
| Hylocomium splendens | 0.148 | 0.083 | 0.086 | 0.046 | 1194 |
| Pleurozium schreberi | 0.718 | 0.070 | 0.007 | 0.268 | 1262 |
| Polytrichum spp. | 0.825 | 0.997 | 0.186 | 0.517 | 465 |
| Sphagnum spp. | 0.275 | 0.513 | 0.686 | 0.257 | 629 |
Appendix B
- Heat indicator—range from 1 (high alpine) to 13 (climate cultivation zone 1 in southernmost Sweden)
- Continentality indicator—range from 1 (hyperoceanic) to 9 (hypercontinental)
- Light indicator—range from 1 (deep shade) to 7 (always in full sun)
- Moisture—range from 1 (very dry) to 12 (deep permanent water)
- Soil pH—range from 1 (strongly acidic, pH < 4.5) to 8 (alkaline, pH > 7.5)
- Nitrogen—range from 1 (very N-poor) to 9 (mostly on artificially N-enriched soils)
| Species | No. of Regions | Heat | Continentality | Light | Moisture | Soil (pH) | Nitrogen |
|---|---|---|---|---|---|---|---|
| Species included with decreasing cover | |||||||
| Maianthemum bifolium | 3 | 4 | 4 | 3 | 4 | 3 | 3 |
| Narrow leaved grasses * | 3 | 2 | 5 | 4 | 4 | 3 | 5 |
| Melampyrum spp.** | 2 | 3.5 | 5.5 | 4.5 | 3.5 | 3 | 3.5 |
| Equisetum sylvaticum | 2 | 3 | 5 | 4 | 5 | 2 | 3 |
| Carex globularis | 2 | 4 | 8 | 4 | 6 | 2 | 2 |
| Rubus chamaemorus | 2 | 3 | 5 | 4 | 6 | 1 | 2 |
| Vaccinium myrtillus | 2 | 3 | 5 | 4 | 5 | 2 | 2 |
| Empetrum nigrum | 2 | 3 | 6 | 5 | 6 | 2 | 3 |
| Gymnocarpium dryopteris | 1 | 3 | 5 | 2 | 4 | 4 | 6 |
| Oxalis acetosella | 1 | 4 | 4 | 2 | 5 | 4 | 5 |
| Anemone nemorosa | 1 | 5 | 4 | 4 | 5 | 4 | 6 |
| Calluna vulgaris | 1 | 5 | 5 | 5 | 4 | 2 | 2 |
| Vaccinium uliginosum | 1 | 2 | 6 | 5 | 6 | 2 | 2 |
| Mean value | 3.5 | 5.2 | 3.8 | 4.9 | 2.9 | 3.4 | |
| NFI species (N=36) not included in the study or not showing decreased cover | |||||||
| Mean value | 4.8 | 4.9 | 4.0 | 5.4 | 4.7 | 5.4 | |
Appendix C
| r2 | p-Value | |
|---|---|---|
| Region | 0.0304 | <0.0001 |
| Period | 0.0021 | =0.0040 |
| NMDS1 | NMDS2 | NMDS3 | NMDS4 | |
|---|---|---|---|---|
| Graminoids | −0.48293 | 0.178719 | 0.513926 | 0.278248 |
| Herbs | −0.658058 | 0.256932 | −0.1777114 | −0.4974966 |
| Spore plants | −1.053622 | −0.587013 | −0.7484537 | 0.465134 |
| Dwarf shrubs | 0.307069 | −0.163221 | −0.0674924 | 0.0319021 |
| Lichen | 0.647159 | 0.894044 | −0.339901 | 0.2470946 |
| Mosses | 0.197559 | −0.157064 | 0.1328394 | −0.1176820 |
| Region | Time | NMDS1 | NMDS2 | NMDS3 | NMDS4 |
|---|---|---|---|---|---|
| 1 | 1 | 0.10220234 | −0.06057463 | −0.0929060 | 0.06475263 |
| 2 | 1 | −0.01220946 | 0.00021519 | −0.0994352 | 0.02586666 |
| 3 | 1 | −0.06833441 | 0.07131811 | 0.01481396 | −0.01272042 |
| 4 | 1 | −0.07908161 | 0.03532577 | 0.14189990 | −0.03512510 |
| 1 | 2 | 0.12696383 | −0.06684115 | −0.06356854 | 0.06253225 |
| 2 | 2 | 0.02208145 | −0.02946240 | −0.08038184 | 0.00224422 |
| 3 | 2 | −0.00856692 | 0.01606524 | 0.0243088 | −0.03468139 |
| 4 | 2 | −0.04081097 | −0.00274272 | 0.12981631 | −0.04795374 |

References
- Nilsson, M.-C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y.H. Understory Vegetation Dynamics of North American boreal forests. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Bergeron, Y. Species and stand dynamics in the mixed woods of Quebec’s southern boreal forest. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Bergstedt, J.; Milberg, P. The impact of logging intensity on field-later vegetation in Swedish boreal forests. For. Ecol. Manag. 2001, 154, 105–115. [Google Scholar] [CrossRef]
- Hedwall, P.O.; Brunet, J.; Nordin, A.; Bergh, J. Changes in the abundance of keystone forest floor species in response to changes of forest structure. J. Veg. Sci. 2013, 24, 296–306. [Google Scholar] [CrossRef]
- Hedwall, P.O.; Gustafsson, L.; Brunet, J.; Lindbladh, M.; Axelsson, A.-L.; Strengbom, J. Half a century of multiple anthropogenic stressors has altered northern forest understory plant communities. Ecol. Appl. 2019, 29, e01874. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phoenix, G.K.; Emmett, B.A.; Britton, A.J.; Caporn, S.J.M.; Dise, N.B.; Helliwell, R.; Jones, L.; Leake, J.R.; Leith, I.D.; Sheppard, L.J.; et al. Impacts of atmospheric nitrogen deposition: Responses of multiple plant and soil parameters across contrasting ecosystems in long-term field experiments. Glob. Chang. Biol. 2012, 18, 1197–1215. [Google Scholar] [CrossRef]
- Villén-Perez, S.; Heikkinen, J.; Salemaa, M.; Mäkipää, R. Global warming will affect the maximum potential abundance of boreal plant species. Ecography 2020, 43, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Forest Europe 2015. In Proceedings of the State of Europe’s Forests 2015, Ministerial Conference on the Protection of Forests in Europe, Madrid, Spain, 20–21 October 2015.
- Wulder, M.A.; Campbell, C.; White, J.C.; Flannigan, M.; Campbell, I.D. National circumstances in the international circumboreal community. For. Chron. 2007, 83, 539–556. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2014: Summary for Policymakers, Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Tamm, C.O. Nitrogen in terrestrial ecosystems. Questions of productivity, vegetational change and ecosystem stability. Ecol. Stud. 1991, 81, 1–116. [Google Scholar]
- Strengbom, J.; Nordin, A. Commercial forest fertilization causes long-term residual effects in ground vegetation of boreal forests. For. Ecol. Manag. 2008, 256, 2175–2181. [Google Scholar] [CrossRef]
- SLU 2019. Forest statistics 2019–Official Statistics of Sweden; Swedish University of Agricultural Sciences: Umeå, Sweden, 2019. [Google Scholar]
- Perring, M.P.; Bernhardt-Römermann, M.; Baeten, L.; Midolo, G.; Blondeel, H.; Depauw, L.; Landuyt, D.; Maes, S.L.; De Lombaerde, E.; Carón, M.M.; et al. Global environmental change effects on plant community composition trajectories depend upon management legacies. Glob. Chang. Biol. 2018, 24, 1722–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridman, J.; Holm, S.; Nilsson, M.; Nilsson, P.; Ringvall, A.H.; Ståhl, G. Adapting National Forest Inventories to changing requirements–the case of the Swedish National Forest Inventory at the turn of the 20th century. Silva Fenn. 2014, 48, 1095. [Google Scholar] [CrossRef] [Green Version]
- FAO. Global Forest Resources Assessment. Terms and Definitions; Food and Agriculture Organization of the United Nations (FAO), Forestry Department: Rome, Italy, 2010. [Google Scholar]
- Östlund, L.; Zackrisson, O.; Axelsson, A.-L. The history and transformation of a Scandinavian boreal forest landscape since the 19th century. Can. J. For. Res. 1997, 27, 1198–1206. [Google Scholar] [CrossRef]
- Milberg, P.; Bergstedt, J.; Fridman, J.; Odell, G.; Westerberg, L. Observer bias and random variation in vegetation monitoring data. J. Veg. Sci. 2008, 19, 633–644. [Google Scholar] [CrossRef] [Green Version]
- Odell, G. (Ed.) Fältinstruktion 2018: Riksinventering av Skog; Swedish University of Agricultural Sciences: Umeå, Swedish, 2018. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 20 December 2018).
- Tyler, T.; Herbertsson, L.; Olofsson, J.; Olsson, P.A. Ecological indicator and traits values for Swedish vascular plants. Ecol. Indic. 2021, 120, 106923. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, M.; et al. Vegan, Community Ecology Package, Version 2.5-2; Available online: http://www.R-project.org/ (accessed on 20 December 2018).
- Hedwall, P.O.; Brunet, J. Trait variations of ground flora species disentangle the effects of global change and altered land-use in Swedish forests during 20 years. Glob. Chang. Biol. 2016, 22, 4038–4047. [Google Scholar] [CrossRef]
- Tonteri, T.; Salemaa, M.; Rautio, P.; Hallikainen, V.; Korpela, L.; Merilä, P. Forest management regulates temporal change in the cover of boreal plant species. For. Ecol. Manag. 2016, 381, 115–124. [Google Scholar] [CrossRef]
- Uotila, A.; Kouki, J. Understorey vegetation in spruce-dominated forests in eastern Finland and Russian Karelia: Successional patterns after anthropogenic and natural disturbances. For. Ecol. Manag. 2005, 215, 113–137. [Google Scholar] [CrossRef]
- Kardell, L. Occurrence and production of bilberry, lingonberry and raspberry in Sweden’s forests. For. Ecol. Manag. 1980, 2, 285–298. [Google Scholar] [CrossRef]
- Palmqvist, K. Carbon economy in lichens. New Phytol. 2000, 148, 11–36. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Palmqvist, K.; Solhaug, K.A.; Holien, H.; Hilmo, O.; Nybakken, L.; Myhre, L.C.; Ohlson, M. Growth of epiphytic old forest lichens across climatic and successional gradients. Can. J. For. Res. 2007, 37, 1832–1845. [Google Scholar] [CrossRef]
- Cabrajic, A.V.J.; Moen, J.; Palmqvist, K. Predicting growth of mat-forming lichens on a landscape scale–Comparing models with different complexities. Ecography 2010, 33, 949–960. [Google Scholar] [CrossRef]
- SMHI 2018. Official Data from the Swedish Meteorological and Hydrological Institute. Available online: https://www.smhi.se/klimat/ (accessed on 15 January 2019).
- Walker, M.D.; Wahren, C.H.; Hollister, R.D.; Henry, G.H.R.; Ahlquist, L.E.; Alatalo, J.M.; Bret-Harte, M.S.; Calef, M.P.; Callaghan, T.V.; Carroll, A.M.; et al. Plant community responses to experimental warming across the tundra biome. Proc. Natl. Acad. Sci. USA 2006, 103, 1342–1346. [Google Scholar] [CrossRef] [Green Version]
- De Frenne, P.; Rodrıguez-Sanchez, F.; Coomes, D.A.; Baeten, L.; Verstraeten, G.; Vellend, M.; Bernhardt-Römermann, M.; Brown, C.D.; Brunet, J.; Cornelis, J.; et al. Microclimate moderates plant responses to macroclimate warming. Proc. Natl. Acad. Sci. USA 2013, 110, 18561–18565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedwall, P.O.; Skoglund, J.; Linder, S. Interactions with successional stage and nutrient status determines the life-form-specific effects of increased soil temperature on boreal forest floor vegetation. Ecol. Evol. 2015, 5, 948–960. [Google Scholar] [CrossRef]
- Jungqvist, G.; Oni, S.K.; Teutschbein, C.; Futter, M.N. Effect on soil temperature in Swedish boreal forests. PLoS ONE 2014, 9, e93957. [Google Scholar] [CrossRef] [Green Version]
- Kreyling, J.; Haei, M.; Laudon, H. Absence of snow cover reduces understory plant cover and alters plant community composition in boreal forest. Oecologia 2012, 168, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Binkley, D.; Högberg, P. Tamm Review: Revisiting the influence of nitrogen deposition on Swedish forests. For. Ecol. Manag. 2016, 368, 222–239. [Google Scholar] [CrossRef]
- Strengbom, J.; Walheim, M.; Näsholm, T.; Ericson, L. Regional Differences in the Occurrence of Understorey Species Reflect Nitrogen Deposition in Swedish Forests. Ambio 2003, 32, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Pihl Karlsson, G.; Hellsten, S.; Karlsson, P.E.; Akselsson, C.; Ferm, M. Kvävedepositionen till Sverige: Jämförelse av Depositionsdata Från Krondroppsnätet, Luft- och Nederbördskemiska Nätet Samt EMEP; IVL Swedish Environmental Research Institute Rapport B2030: Stockholm, Sweden, 2012. [Google Scholar]
- Engardt, M.; Simpson, D.; Schwikowski, M.; Granat, L. Deposition of sulphur and nitrogen in Europe 1900–2050. Model calculations and comparison to historical observations. Tellus B Chem. Phys. Meteorol. 2017, 69, 1328945. [Google Scholar] [CrossRef] [Green Version]
- Kellner, O.; Redbo-Torstensson, P. Effects of elevated nitrogen deposition on the field-layer vegetation in coniferous forests. Ecol. Bull. 1995, 44, 227–237. [Google Scholar]
- Wardle, D.A.; Jonsson, M.; Bansal, S.; Bardgett, R.D.; Gundale, M.J.; Metcalfe, D.B. Linking vegetation change, carbon sequestration and biodiversity: Insights from island ecosystems in a long-term natural experiment. J. Ecol. 2012, 100, 16–30. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, T. Förteckning över svenska kärlväxter. Sven. Bot. Tidskr. 1997, 91, 241–560. [Google Scholar]
- Stenroos, S.; Velmala, S.; Pykälä, J.; Ahti, T. Lichens of Finland; Botanical Museum, Finnish Museum of Natural History LUOMUS, University of Helsinki: Helsinki, Finland, 2016. [Google Scholar]
- Hallingbäck, T.; Hedenäs, L.; Weibull, H. Ny checklista för Sveriges mossor. Sven. Bot. Tidskr. 2006, 100, 96–148. [Google Scholar]


| Maturity Class * | ||||
|---|---|---|---|---|
| Region | 33 | 34 | 41 | 42 |
| 1 | 70 (17.5%) | 89 (5.9%) | 102 (32.0%) | 156 (44.6%) |
| 2 | 64 (20.0%) | 89 (8.6%) | 96 (25.9%) | 139 (45.5%) |
| 3 | 55 (26.5%) | 77 (7.0%) | 81 (20.3%) | 122 (46.3%) |
| 4 | 50 (23.1%) | 69 (6.8%) | 70 (26.1%) | 99 (44.1%) |
| First Inventory | Second Inventory | |||||
|---|---|---|---|---|---|---|
| Region | Mean | SD | Mean | SD | N | Difference |
| Basal area (m2 ha−1) | ||||||
| 1 | 20.1 | 8.02 | 22.1 | 8.33 | 294 | 2.0 |
| 2 | 24.8 | 9.86 | 27.1 | 10.48 | 312 | 2.2 |
| 3 | 26.30 | 10.53 | 29.2 | 10.99 | 398 | 2.9 |
| 4 | 28.0 | 9.69 | 30.7 | 10.75 | 296 | 2.8 |
| Total | 24.9 | 10.05 | 27.4 | 10.72 | 1300 | 2.5 |
| Canopy cover (%) | ||||||
| 1 | 55.6 | 16.77 | 60.9 | 15.21 | 46 | 5.2 |
| 2 | 56.1 | 18.42 | 57.0 | 13.38 | 44 | 1.0 |
| 3 | 50.9 | 17.86 | 63.9 | 11.73 | 55 | 13.0 |
| 4 | 58.5 | 12.72 | 63.2 | 12.19 | 34 | 4.7 |
| Total | 54.8 | 16.96 | 61.3 | 13.36 | 179 | 6.5 |
| Cover (%) | ||||||
|---|---|---|---|---|---|---|
| Species * | Region | N | Initial | Change Absolute (Relative) | LCL | UCL |
| Graminoids | ||||||
| Broad leaved grasses | 3 | 187 | 6.8 | −1.7 (−25%) | −3.04 | −0.36 |
| Broad leaved grasses | 4 | 135 | 7.6 | −2.3 (−30%) | −3.97 | −0.53 |
| Carex globularis | 2 | 69 | 1.7 | −0.6 (−34%) | −1.01 | −0.11 |
| Carex globularis | 3 | 80 | 2.0 | −0.8 (−42%) | −1.51 | −0.19 |
| Narrow leaved grasses | 2 | 240 | 3.4 | −1.2 (−35%) | −1.73 | −0.64 |
| Narrow leaved grasses | 3 | 274 | 5.0 | −2.0 (−40%) | −3.08 | −0.97 |
| Narrow leaved grasses | 4 | 208 | 5.8 | −2.1 (−36%) | −3.32 | −0.92 |
| Herbs | ||||||
| Anemone nemorosa | 3 | 107 | 2.1 | −0.8 (−38%) | −1.55 | −0.02 |
| Maianthemum bifolium | 1 | 62 | 3.7 | −1.6 (−43%) | −3.03 | −0.21 |
| Maianthemum bifolium | 3 | 160 | 1.7 | −0.4 (−26%) | −0.80 | −0.09 |
| Maianthemum bifolium | 4 | 111 | 1.9 | −0.9 (−49%) | −1.42 | −0.43 |
| Melampyrum spp. | 3 | 245 | 1.2 | −0.3 (−27%) | −0.62 | −0.01 |
| Melampyrum spp. | 4 | 155 | 1.0 | −0.3 (−31%) | −0.52 | −0.07 |
| Oxalis acetosella | 2 | 89 | 3.4 | −1.3 (−38%) | −2.17 | −0.43 |
| Dwarf shrubs | ||||||
| Calluna vulgaris | 3 | 164 | 5.3 | −0.9 (−16%) | −1.68 | −0.05 |
| Empetrum nigrum | 3 | 93 | 3.7 | −1.1 (−29%) | −1.82 | −0.31 |
| Empetrum nigrum | 4 | 27 | 1.2 | −0.8 (−68%) | −1.21 | −0.42 |
| Rubus chamaemorus | 1 | 52 | 4.3 | −2.1 (−50%) | −3.92 | −0.34 |
| Rubus chamaemorus | 3 | 49 | 4.2 | −1.5 (−36%) | −2.85 | −0.19 |
| Vaccinium myrtillus | 1 | 291 | 26.2 | −5.8 (−22%) | −7.51 | −4.07 |
| Vaccinium myrtillus | 2 | 307 | 22.6 | −4.0 (−17%) | −5.49 | −2.42 |
| Vaccinium uliginosum | 4 | 52 | 5.6 | −1.8 (−31%) | −3.43 | −0.08 |
| Spore plants | ||||||
| Equisetum sylvaticum | 1 | 80 | 3.5 | −2.2 (−64%) | −3.38 | −1.04 |
| Equisetum sylvaticum | 2 | 91 | 1.4 | −0.5 (−35%) | −0.82 | −0.14 |
| Gymnocarpium dryopteris | 1 | 52 | 6.8 | −3.0 (−44%) | −5.02 | −1.01 |
| Lichens | ||||||
| Cladonia spp. excl. Cladina | 2 | 196 | 0.4 | −0.1 (−32%) | −0.23 | −0.03 |
| Cladonia spp. excl. Cladina | 4 | 162 | 0.4 | −0.2 (−48%) | −0.28 | −0.07 |
| Cladonia grp. Cladina | 2 | 162 | 6.3 | −3.0 (−47%) | −4.15 | −1.78 |
| Cladonia grp. Cladina | 3 | 184 | 8.3 | −2.5 (−30%) | −3.56 | −1.47 |
| Cladonia grp. Cladina | 4 | 81 | 2.8 | −0.9 (−33%) | −1.85 | −0.02 |
| Other ground lichens | 2 | 151 | 0.9 | −0.2 (−25%) | −0.39 | −0.03 |
| Other ground lichens | 3 | 195 | 1.1 | −0.4 (−33%) | −0.68 | −0.05 |
| Mosses | ||||||
| Hylocomium splendens | 3 | 358 | 16.9 | 3.6 (21%) | 1.82 | 5.34 |
| Hylocomium splendens | 4 | 269 | 14.3 | 5.7 (40%) | 3.59 | 7.79 |
| Pleurozium schreberi | 1 | 293 | 36.2 | −5.9 (−16%) | −8.39 | −3.40 |
| Pleurozium schreberi | 3 | 383 | 22.7 | 2.2 (10%) | 0.46 | 3.94 |
| Pleurozium schreberi | 4 | 276 | 22.5 | −4.4 (−20%) | −6.64 | −2.18 |
| Polytrichum spp. | 2 | 127 | 4.1 | −1.2 (−28%) | −2.09 | −0.21 |
| Sphagnum spp. | 1 | 120 | 21.8 | −3.6 (−17%) | −6.31 | −0.90 |
| Sphagnum spp. | 2 | 157 | 20.2 | −3.0 (−15%) | −5.03 | −0.93 |
| Sphagnum spp. | 4 | 149 | 21.3 | 3.4 (16%) | 0.56 | 6.30 |
| Relative cover Change (%) in Un-Thinned and Thinned Stands | ||
|---|---|---|
| Species | Unthinned | Thinned |
| Melampyrum spp. | −33% (635) | 8% (127) |
| Vaccinium myrtillus | −15% (1033) | 6% (197) |
| Calluna vulgaris | −20% (358) | 59% (84) |
| Carex globularis | −32% (217) | −69% (23) |
| Pleurozium schreberi | −5% (1052) | −16% (210) |
| Relative Cover Change (%) in Maturity Classes | ||||
|---|---|---|---|---|
| Species | 33 | 34 | 41 | 42 |
| Oxalis acetosella | 10% (40) | −59% (14) | −46% (72) | −21% (133) |
| Anemone nemorosa | 8% (51) | −36% (16) | −25% (49) | −65% (79) |
| Vaccinium myrtillus | −1% (185) | −13% (71) | −14% (314) | −14% (660) |
| Lycopodiaceae | −39% (41) | 74% (15) | −2% (70) | −28% (153) |
| Hylocomium splendens | 34% (190) | 23% (73) | 9% (296) | 9% (635) |
| Cladonia grp. Cladina | −54% (77) | −29% (38) | −28% (165) | −22% (327) |
| Cover (%) | ||||||
|---|---|---|---|---|---|---|
| Species Group | N | Initial | Change Absolute (Relative) | LCL | UCL | Direction |
| Graminoids | ||||||
| Region 1 | 245 | 3.6 | −0.5 (−13%) | −0.99 | 0.06 | NS |
| Region 2 | 254 | 6.0 | −1.3 (−22%) | −2.15 | −0.54 | Decrease |
| Region 3 | 332 | 10.2 | −2.8 (−27%) | −4.16 | −1.43 | Decrease |
| Region 4 | 250 | 10.7 | −2.6 (−24%) | −4.12 | −1.09 | Decrease |
| Herbs | ||||||
| Region 1 | 179 | 3.0 | −1.0 (−33%) | −1.57 | −0.37 | Decrease |
| Region 2 | 230 | 4.6 | −0.9 (−19%) | −1.51 | −0.19 | Decrease |
| Region 3 | 309 | 3.8 | −0.8 (−20%) | −1.34 | −0.17 | Decrease |
| Region 4 | 225 | 4.1 | −1.1 (−28%) | −1.93 | −0.32 | Decrease |
| Dwarf shrubs | ||||||
| Region 1 | 269 | 56.4 | −8.0 (−14%) | −10.49 | −5.56 | Decrease |
| Region 2 | 286 | 49.7 | −6.7 (−14%) | −8.91 | −4.57 | Decrease |
| Region 3 | 354 | 33.7 | −1.7 (−5%) | −3.55 | 0.17 | NS |
| Region 4 | 277 | 24. 8 | −0.8 (−3%) | −2.70 | 1.12 | NS |
| Spore plants * | ||||||
| Region 1 | 159 | 5.0 | −2.2 (−44%) | −3.23 | −1.11 | Decrease |
| Region 2 | 175 | 5.1 | −1.3 (−25%) | −2.19 | −0.36 | Decrease |
| Region 3 | 124 | 2.9 | −0.3 (−9%) | −0.95 | 0.44 | NS |
| Region 4 | 80 | 2.5 | 0.4 (15%) | −0.40 | 1.18 | NS |
| Lichens | ||||||
| Region 1 | 221 | 5.7 | −0.6 (−11%) | −1.68 | 0.48 | NS |
| Region 2 | 241 | 7.3 | −3.0 (−42%) | −4.08 | −1.99 | Decrease |
| Region 3 | 308 | 6.9 | −2.0 (−28%) | −2.79 | −1.12 | Decrease |
| Region 4 | 222 | 1.6 | −0.4 (−26%) | −0.85 | −0.01 | Decrease |
| Mosses | ||||||
| Region 1 | 269 | 79.4 | −7.6 (−10%) | −10.29 | −4.95 | Decrease |
| Region 2 | 290 | 79.1 | −3.2 (−4%) | −5.91 | −0.42 | Decrease |
| Region 3 | 367 | 63.6 | 5.9 (9%) | 3.56 | 8.33 | Increase |
| Region 4 | 280 | 57.4 | 2.2 (4%) | −0.78 | 5.09 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jonsson, B.G.; Dahlgren, J.; Ekström, M.; Esseen, P.-A.; Grafström, A.; Ståhl, G.; Westerlund, B. Rapid Changes in Ground Vegetation of Mature Boreal Forests—An Analysis of Swedish National Forest Inventory Data. Forests 2021, 12, 475. https://doi.org/10.3390/f12040475
Jonsson BG, Dahlgren J, Ekström M, Esseen P-A, Grafström A, Ståhl G, Westerlund B. Rapid Changes in Ground Vegetation of Mature Boreal Forests—An Analysis of Swedish National Forest Inventory Data. Forests. 2021; 12(4):475. https://doi.org/10.3390/f12040475
Chicago/Turabian StyleJonsson, Bengt Gunnar, Jonas Dahlgren, Magnus Ekström, Per-Anders Esseen, Anton Grafström, Göran Ståhl, and Bertil Westerlund. 2021. "Rapid Changes in Ground Vegetation of Mature Boreal Forests—An Analysis of Swedish National Forest Inventory Data" Forests 12, no. 4: 475. https://doi.org/10.3390/f12040475