
Can a Remote Sensing Approach with Hyperspectral Data Provide Early Detection and Mapping of Spatial Patterns of Black Bear Bark Stripping in Coast Redwoods?

Abstract
:1. Introduction
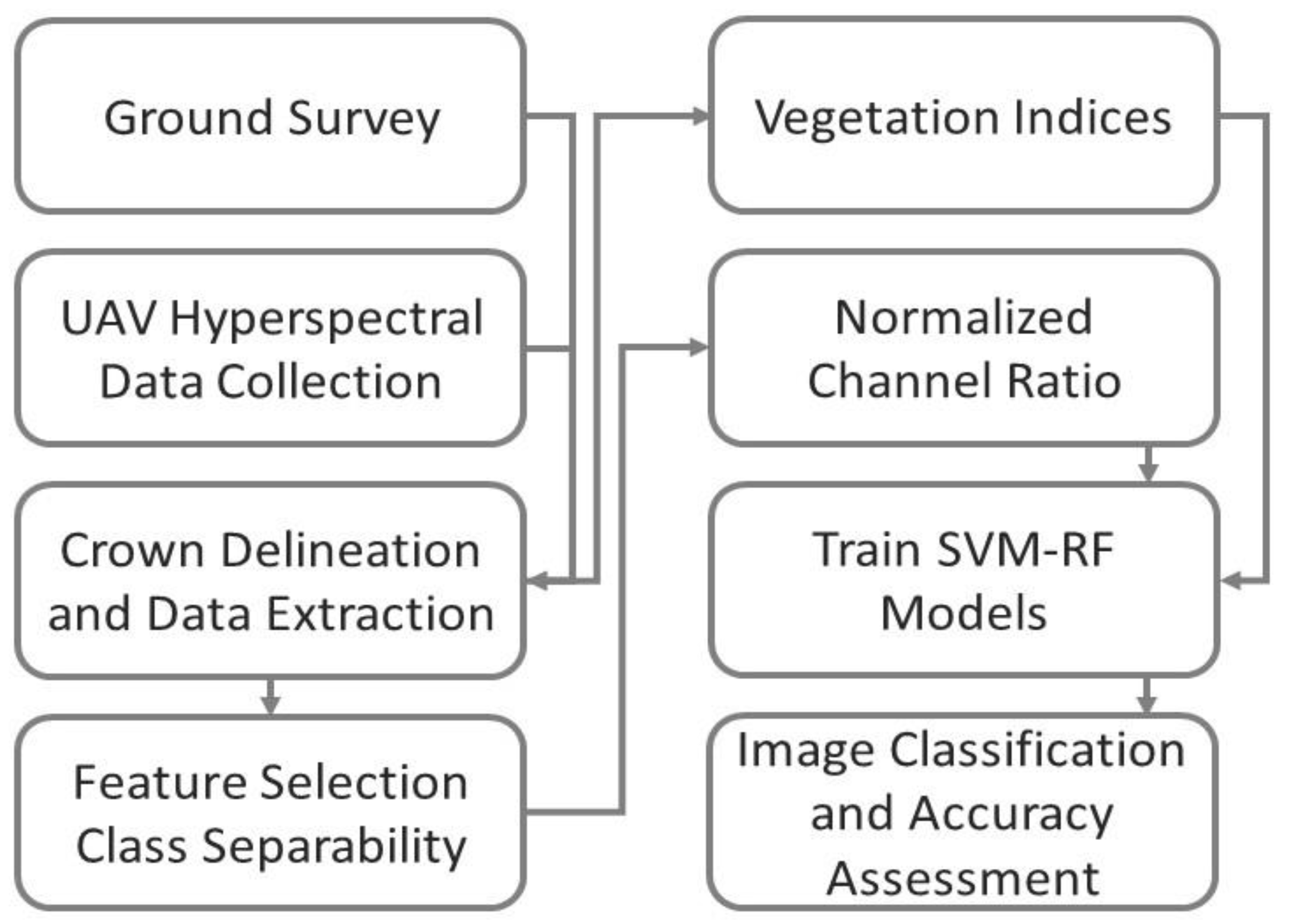
2. Materials and Methods
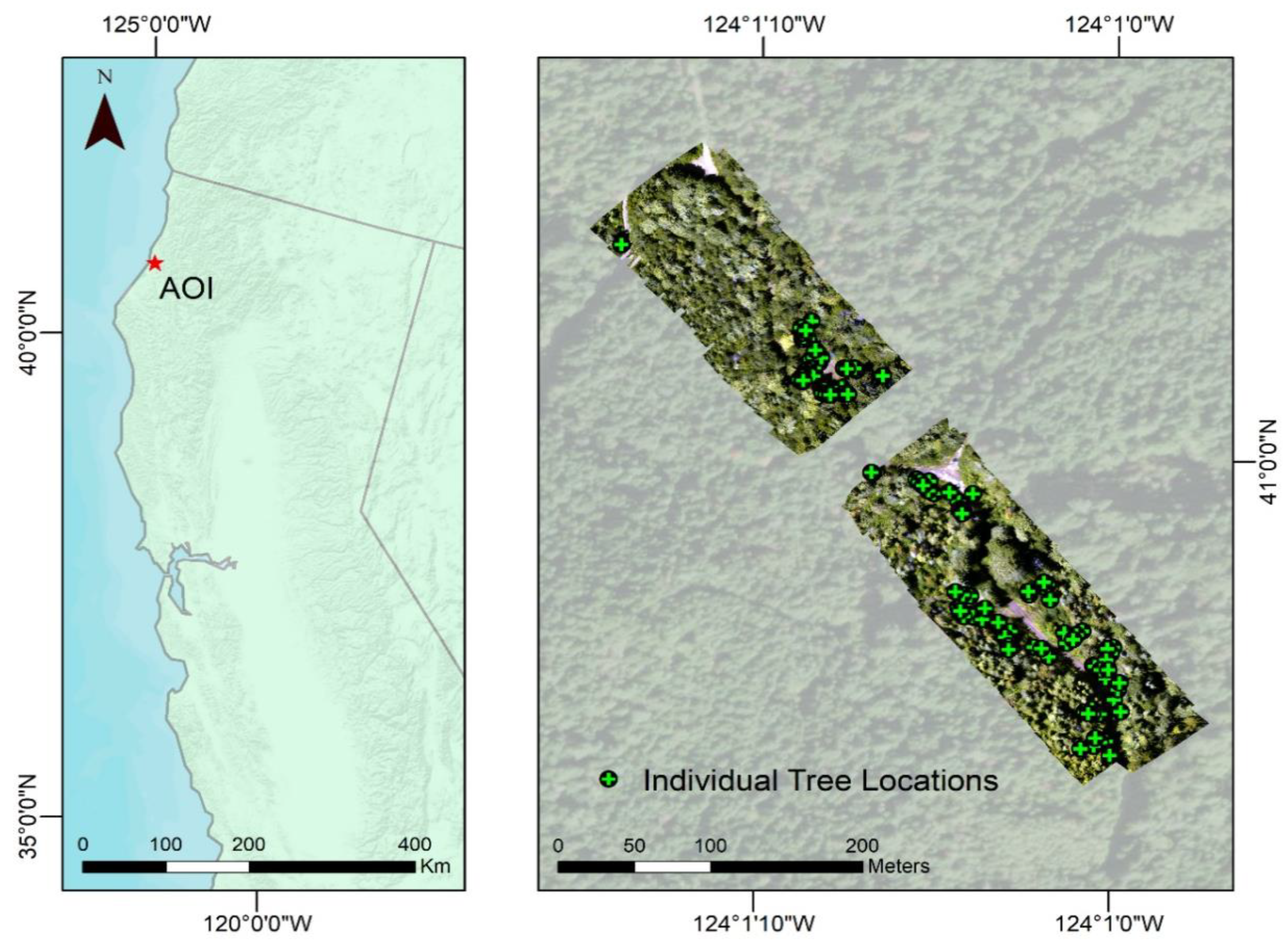
2.1. Site Location
2.2. Experimental Design/Treatment Details
2.3. Field Survey
2.4. UAV Data Collection and Processing
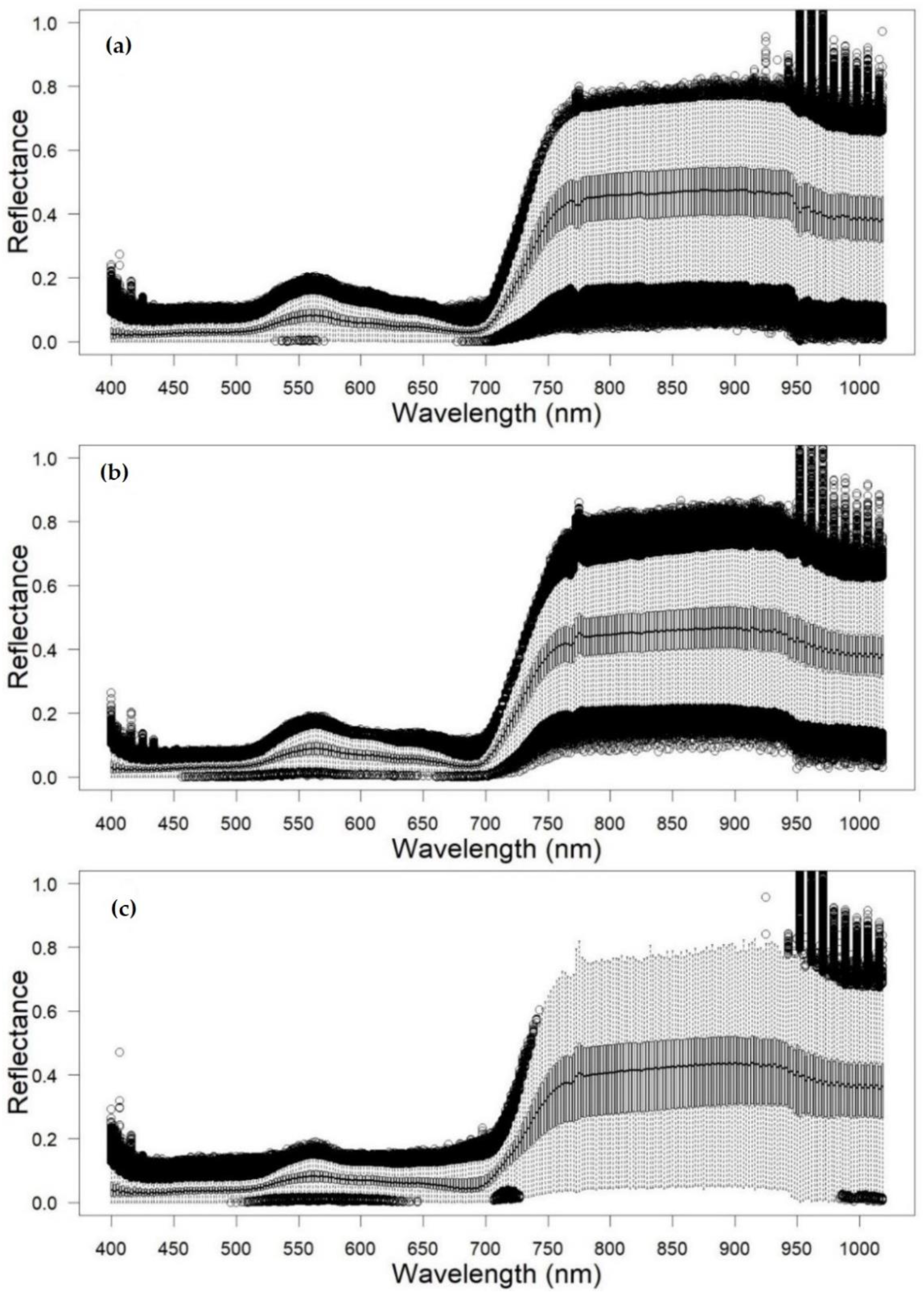
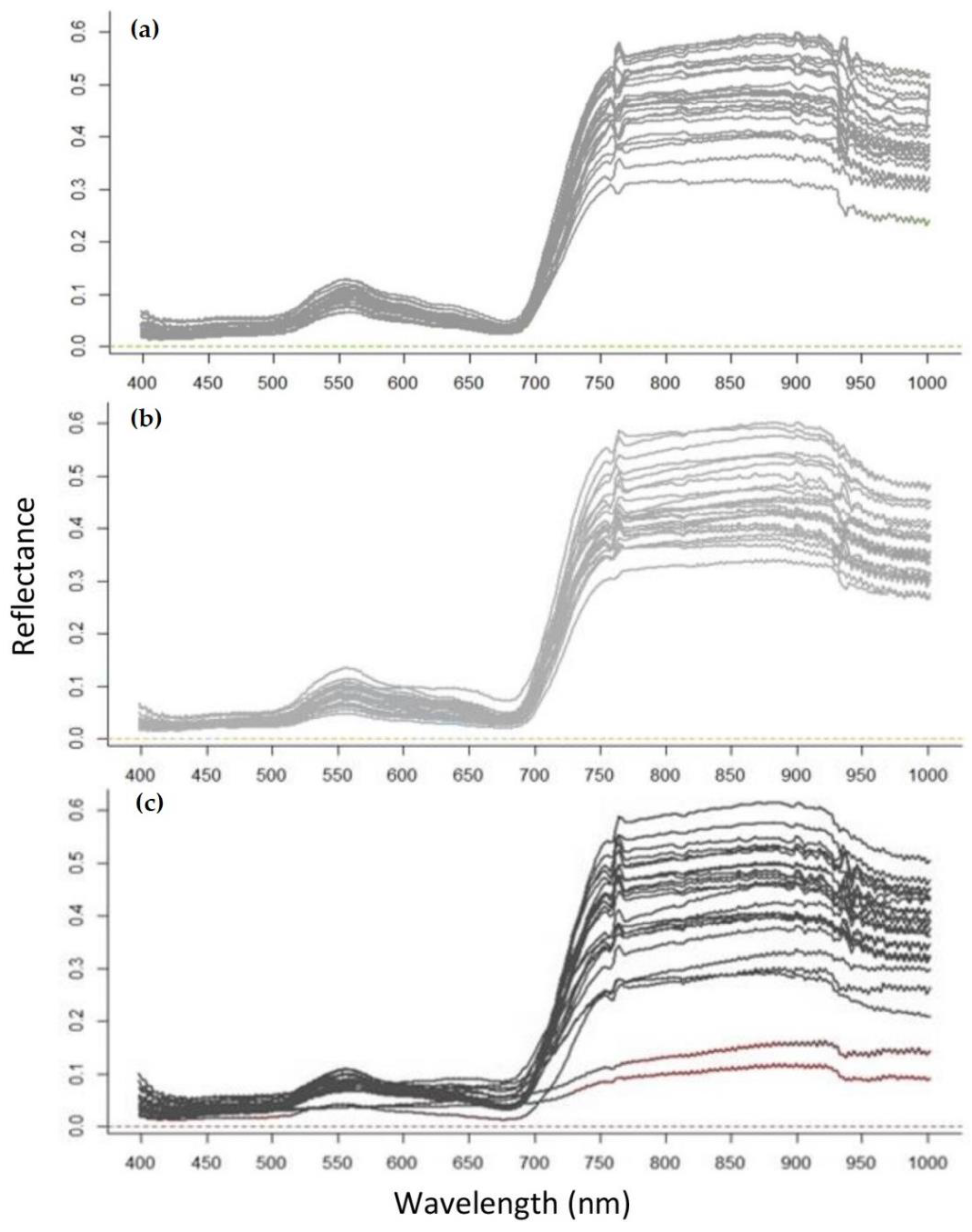
2.5. Crown Delineation and Spectral Signature Extraction
2.6. Feature Selection and Vegetation Indices Based on Class Separability
2.7. Classification
2.8. Accuracy Assessment
3. Results
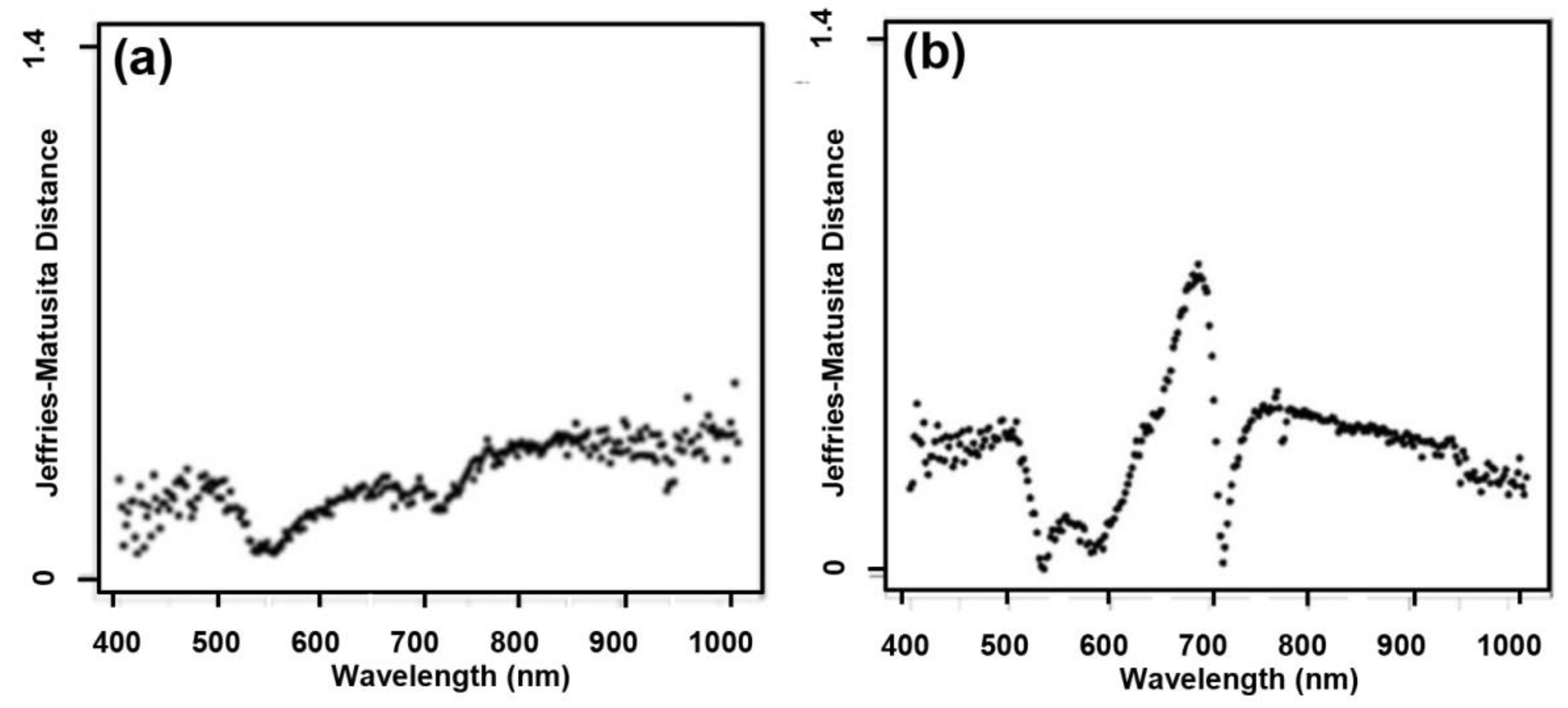
3.1. Feature Selection
3.2. Classification Accuracy
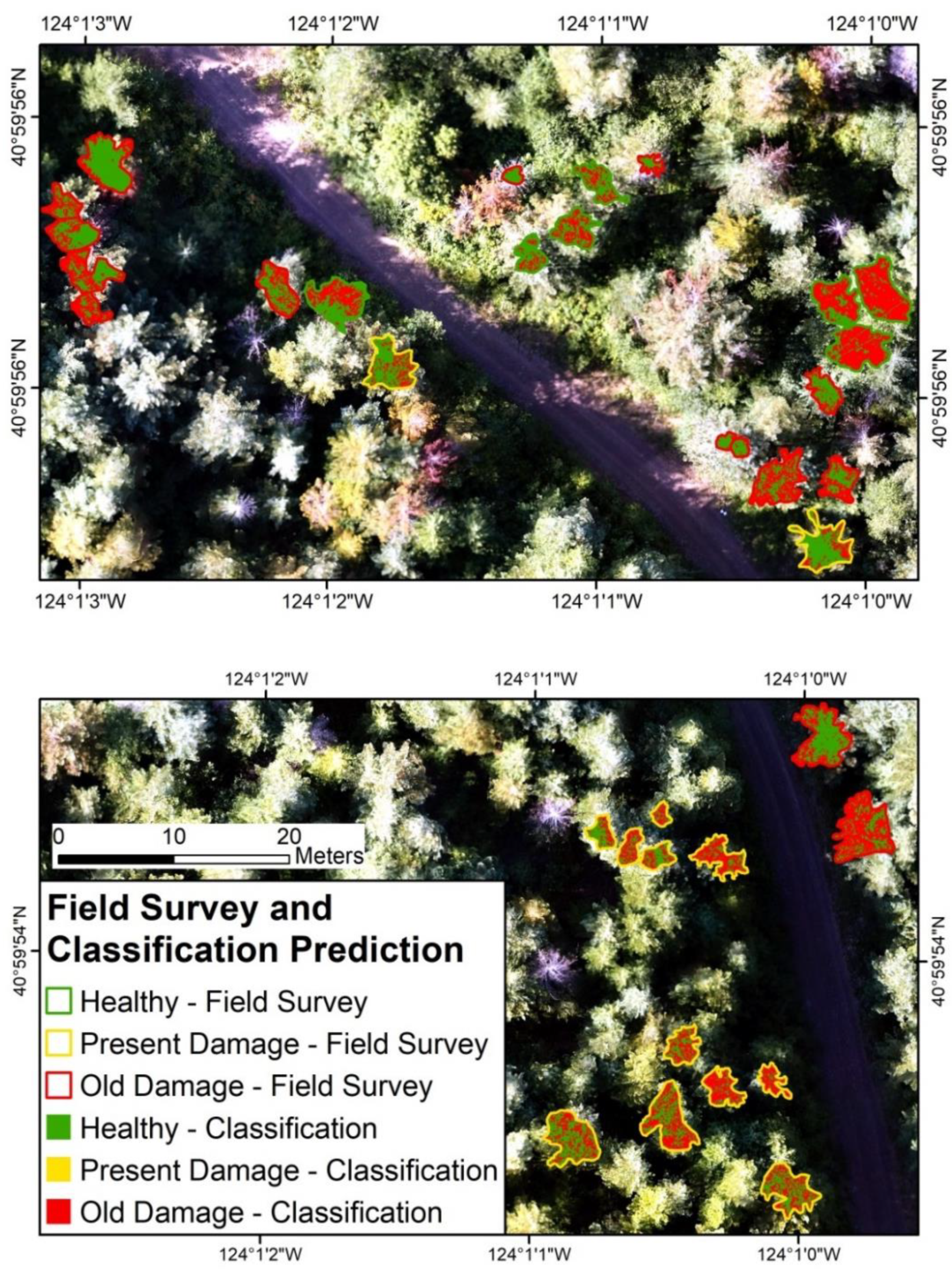
3.3. Model Prediction
4. Discussion
4.1. Redwood Tree Characteristics
4.2. Feature Selection, Variable Importance and Early Detection of Damage
4.3. UAV-Based Image Acquisition in Forest Health Monitoring
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sillett, S.C.; Van Pelt, R.; Carroll, A.L.; Kramer, R.D.; Ambrose, A.R.; Trask, D. How do tree structure and old age affect growth potential of California redwoods? Ecol. Monogr. 2015, 85, 181–212. [Google Scholar] [CrossRef]
- Brown, C. Habitat Use and Movement Patterns of Two Redwood Forest Salamanders, Aneides Vagrans and Ensatina Eschscholtzii, with an Examination of the Efficacy of Pit Tags for Marking Small Plethodontids; Humboldt State University: Arcata, CA, USA, 2017. [Google Scholar]
- Fish, U.S.; Service, W. Endangered and Threatened Wildlife and Plants: Determination of Threatened Status for the Northern Spotted Owl. Fed. Regist. 1990, 55, 26114–26194. [Google Scholar]
- Lorimer, C.G.; Porter, D.J.; Madej, M.A.; Stuart, J.D.; Veirs, S.D., Jr.; Norman, S.P.; O’Hara, K.L.; Libby, W.J. Presettlement and modern disturbance regimes in coast redwood forests: Implications for the conservation of old-growth stands. For. Ecol. Manag. 2009, 258, 1038–1054. [Google Scholar] [CrossRef]
- Ziegltrum, G. Cost-Effectiveness of the Black Bear Supplemental Feeding Program in Western Washington. Wildife Soc. Bull. 2006, 34, 375–379. [Google Scholar] [CrossRef]
- Giusti, G. Recognizing Damage by Black Bear Damage to Second Growth Redwoods. In Proceedings of the 13th Vertebrate Pest Conference, Monterey, CA, USA, 1–3 March 1988. [Google Scholar]
- Giusti, G. UC Agriculture & Natural Resources Proceedings of the Vertebrate Pest Conference. In Proceedings of the Vertebrate Pest Conference, Sacramento, CA, USA, 6–8 March 1990. [Google Scholar]
- Glover, F. Glover Source. J. Wildl. Manag. 1955, 19, 437–443. [Google Scholar] [CrossRef]
- Matthews, S.M.; Golightly, R.T.; Higley, J.M. Mark–resight density estimation for American black bears in Hoopa, California. Ursus 2008, 19, 13–21. [Google Scholar] [CrossRef]
- Ziegltrum, G.J. Efficacy of black bear supplemental feeding to reduce conifer damage in western washington. J. Wildl. Manag. 2004, 68, 470–474. [Google Scholar] [CrossRef] [Green Version]
- Kanaskie, A.; Chetock, J.; Irwin, G.; Overhusler, D. Black Bear Damage to Forest Trees in Northwest Oregon 1988–1989. Pest Manag. Rep. 1990, 90, 34p. [Google Scholar]
- Taylor, J.D.; Kline, K.N.; Morzillo, A.T. Estimating economic impact of black bear damage to western conifers at a landscape scale. For. Ecol. Manag. 2019, 432, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Nolte, D.L.; Dykzeul, M. Wildlife Impacts on Forest Resources. Hum. Confl. Wildl. Econ. Consid. 2000, 20, 163–168. [Google Scholar]
- Panigrahy, S.; Kumar, T.; Manjunath, K.R. Hyperspectral leaf signature as an added dimension for species discrimination: Case study of four tropical mangroves. Wetl. Ecol. Manag. 2012, 20, 101–110. [Google Scholar] [CrossRef]
- Foster, A.C.; Walter, J.A.; Shugart, H.H.; Sibold, J.; Negron, J. Spectral evidence of early-stage spruce beetle infestation in Engelmann spruce. For. Ecol. Manag. 2017, 384, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Hall, R.; Castilla, G.; White, J.; Cooke, B.; Skakun, R. Remote sensing of forest pest damage: A review and lessons learned from a Canadian perspective. Can. Èntomol. 2016, 148, S296–S356. [Google Scholar] [CrossRef]
- Näsi, R.; Honkavaara, E.; Lyytikäinen-Saarenmaa, P.; Blomqvist, M.; Litkey, P.; Hakala, T.; Viljanen, N.; Kantola, T.; Tanhuanpää, T.; Holopainen, M. Using UAV-Based Photogrammetry and Hyperspectral Imaging for Mapping Bark Beetle Damage at Tree-Level. Remote Sens. 2015, 7, 15467–15493. [Google Scholar] [CrossRef] [Green Version]
- Näsi, R.; Honkavaara, E.; Blomqvist, M.; Lyytikäinen-Saarenmaa, P.; Hakala, T.; Viljanen, N.; Kantola, T.; Holopainen, M. Remote sensing of bark beetle damage in urban forests at individual tree level using a novel hyperspectral camera from UAV and aircraft. Urban For. Urban Green. 2018, 30, 72–83. [Google Scholar] [CrossRef]
- Carter, G.A. Carter Source. Am. J. Bot. 1993, 80, 239–243. [Google Scholar] [CrossRef]
- Vogelmann, J.E.; Rock, B.N.; Moss, D.M. Red edge spectral measurements from sugar maple leaves. Int. J. Remote Sens. 1993, 14, 1563–1575. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Rakitin, V.Y. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Dash, J.P.; Watt, M.S.; Pearse, G.D.; Heaphy, M.; Dungey, H.S. Assessing very high resolution UAV imagery for monitoring forest health during a simulated disease outbreak. ISPRS J. Photogramm. Remote Sens. 2017, 131, 1–14. [Google Scholar] [CrossRef]
- Adam, E.; Mutanga, O.; Rugege, D. Multispectral and hyperspectral remote sensing for identification and mapping of wetland vegetation: A review. Wetl. Ecol. Manag. 2010, 18, 281–296. [Google Scholar] [CrossRef]
- Ortiz, S.; Breidenbach, J.; Kändler, G. Early Detection of Bark Beetle Green Attack Using TerraSAR-X and RapidEye Data. Remote Sens. 2013, 5, 1912–1931. [Google Scholar] [CrossRef] [Green Version]
- Sothe, C.; Dalponte, M.; De Almeida, C.M.; Schimalski, M.B.; Lima, C.L.; Liesenberg, V.; Miyoshi, G.T.; Tommaselli, A.M.G. Tree Species Classification in a Highly Diverse Subtropical Forest Integrating UAV-Based Photogrammetric Point Cloud and Hyperspectral Data. Remote Sens. 2019, 11, 1338. [Google Scholar] [CrossRef] [Green Version]
- Coops, N.C.; Waring, R.H.; Wulder, M.A.; White, J.C. Prediction and assessment of bark beetle-induced mortality of lodgepole pine using estimates of stand vigor derived from remotely sensed data. Remote Sens. Environ. 2009, 113, 1058–1066. [Google Scholar] [CrossRef]
- Turner, D.; Lucieer, A.; Watson, C. Development of an Unmanned Aerial Vehicle (UAV) for Hyper Resolution Vineyard Mapping Based on Visible, Multispectral, and Thermal Imagery. In Proceedings of the 34th International Symposium on Remote Sensing of Environment, Sydney, Australia, 10–15 April 2011. [Google Scholar]
- Maschler, J.; Atzberger, C.; Immitzer, M. Individual Tree Crown Segmentation and Classification of 13 Tree Species Using Airborne Hyperspectral Data. Remote Sens. 2018, 10, 1218. [Google Scholar] [CrossRef] [Green Version]
- Alonzo, M.; Roth, K.; Roberts, D. Identifying Santa Barbara’s urban tree species from AVIRIS imagery using canonical discriminant analysis. Remote Sens. Lett. 2013, 4, 513–521. [Google Scholar] [CrossRef]
- Alonzo, M.; Bookhagen, B.; Roberts, D.A. Urban tree species mapping using hyperspectral and lidar data fusion. Remote Sens. Environ. 2014, 148, 70–83. [Google Scholar] [CrossRef]
- Bunting, P.; Lucas, R. The delineation of tree crowns in Australian mixed species forests using hyperspectral Compact Airborne Spectrographic Imager (CASI) data. Remote Sens. Environ. 2006, 101, 230–248. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.; Hornero, A.; Hernández-Clemente, R.; Beck, P. Understanding the temporal dimension of the red-edge spectral region for forest decline detection using high-resolution hyperspectral and Sentinel-2a imagery. ISPRS J. Photogramm. Remote Sens. 2018, 137, 134–148. [Google Scholar] [CrossRef]
- Degerickx, J.; Roberts, D.; McFadden, J.; Hermy, M.; Somers, B. Urban tree health assessment using airborne hyperspectral and LiDAR imagery. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 26–38. [Google Scholar] [CrossRef] [Green Version]
- Lowe, A.; Harrison, N.; French, A.P. Hyperspectral image analysis techniques for the detection and classification of the early onset of plant disease and stress. Plant Methods 2017, 13, 1–12. [Google Scholar] [CrossRef]
- Esri, A. ArcGIS 10.1; ESRI Inc.: Redlands, CA, USA, 2012. [Google Scholar]
- Bruzzone, L.; Roli, F.; Serpico, S. An extension of the Jeffreys-Matusita distance to multiclass cases for feature selection. IEEE Trans. Geosci. Remote Sens. 1995, 33, 1318–1321. [Google Scholar] [CrossRef] [Green Version]
- Belgiu, M.; Drăguţ, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Medjahed, S.; Saadi, T.A.; Benyettou, A.; Ouali, M. Gray Wolf Optimizer for hyperspectral band selection. Appl. Soft Comput. 2016, 40, 178–186. [Google Scholar] [CrossRef]
- Rouse, J.W., Jr.; Haas, R.H.; Schell, J.A.; Deering, D.W. Paper A 20. In Third Earth Resources Technology Satellite-1 Symposium: The Proceedings of a Symposium Held by Goddard Space Flight Center at Washington, DC on 10–14 December 1973; Goddard Space Flight Center: Greenbelt, MD, USA, 1974; Volume 351, p. 309. [Google Scholar]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; De Colstoun, E.B.; McMurtrey, J.E., III. Estimating Corn Leaf Chlorophyll Concentration from Leaf and Canopy Reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Vogelmann, J.E. Comparison between two vegetation indices for measuring different types of forest damage in the north-eastern United States. Int. J. Remote Sens. 1990, 11, 2281–2297. [Google Scholar] [CrossRef]
- Kantola, T.; Vastaranta, M.; Lyytikäinen-Saarenmaa, P.; Holopainen, M.; Kankare, V.; Talvitie, M.; Hyyppa, J. Classification of Needle Loss of Individual Scots Pine Trees by Means of Airborne Laser Scanning. Forest 2013, 4, 386–403. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-C.; Lin, C.-J. LIBSVM: A Library for Support Vector Machines. ACM Trans. Intell. Syst. Technol. 2011, 2, 27. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. J. Electrochem. Soc. 1999, 129, 2865. [Google Scholar]
- Cortes, C.; Vapnik, V. Support-Vector Networks. Mach. Learn. 1995, 20, 273–297. [Google Scholar] [CrossRef]
- Vapnik, V. Pattern Recognition Using Generalized Portrait Method. Autom. Remote Control 1963, 24, 774–780. [Google Scholar]
- Ebrahimi, M.; Khoshtaghaza, M.; Minaei, S.; Jamshidi, B. Vision-based pest detection based on SVM classification method. Comput. Electron. Agric. 2017, 137, 52–58. [Google Scholar] [CrossRef]
- Gualtieri, J.A. The Support Vector Machine (SVM) Algorithm for Supervised Classification of Hyperspectral Remote Sensing Data. Kernel Methods Remote Sens. Data Anal. 2009, 3, 49–83. [Google Scholar] [CrossRef]
- Hsu, C.-W.; Chang, C.-C.; Lin, C.-J. A Practical Guide to SVM Classification. 2003. Available online: http//www.csie.ntu.edu.tw/~cjlin/papers/guide/guide.pdf (accessed on 6 December 2019).
- Tzotsos, A.; Argialas, D.P. Support Vector Machine Classification for Object-Based Image Analysis. In Lecture Notes in Geoinformation and Cartography; Springer: Berlin/Heidelberg, Germany, 2008; pp. 663–677. [Google Scholar]
- Melgani, F.; Bruzzone, L. Classification of hyperspectral remote sensing images with support vector machines. IEEE Trans. Geosci. Remote Sens. 2004, 42, 1778–1790. [Google Scholar] [CrossRef] [Green Version]
- Joachims, T. Optimizing Search Engines Using Clickthrough Data. In Proceedings of the Eighth ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, Edmonton, AB, USA, 23–26 July 2002; pp. 133–142. [Google Scholar]
- Gall, J.; Yao, A.; Razavi, N.; Van Gool, L.; Lempitsky, V. Hough Forests for Object Detection, Tracking, and Action Recognition. IEEE Trans. Pattern Anal. Mach. Intell. 2011, 33, 2188–2202. [Google Scholar] [CrossRef]
- Rodriguez-Galiano, V.; Ghimire, B.; Rogan, J.; Chica-Olmo, M.; Rigol-Sanchez, J. An assessment of the effectiveness of a random forest classifier for land-cover classification. ISPRS J. Photogramm. Remote Sens. 2012, 67, 93–104. [Google Scholar] [CrossRef]
- Chan, J.C.-W.; Paelinckx, D. Evaluation of Random Forest and Adaboost tree-based ensemble classification and spectral band selection for ecotope mapping using airborne hyperspectral imagery. Remote Sens. Environ. 2008, 112, 2999–3011. [Google Scholar] [CrossRef]
- Gislason, P.O.; Benediktsson, J.A.; Sveinsson, J.R. Random Forests for land cover classification. Pattern Recognit. Lett. 2006, 27, 294–300. [Google Scholar] [CrossRef]
- Metropolis, N.; Ulam, S. The Monte Carlo Method. J. Am. Stat. Assoc. 1949, 44, 335–341. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Kuhn, V.I.; Ranson, K.J.; Fedotova, E.V. Building Predictive Models in R Using the Caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Gates, D.M.; Keegan, H.J.; Schleter, J.C.; Weidner, V.R. Spectral Properties of Plants. Appl. Opt. 1965, 4, 11–20. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral Vegetation Indices and Novel Algorithms for Predicting Green LAI of Crop Canopies: Modeling and Validation in the Context of Precision Agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Knipling, E.B. Physical and physiological basis for the reflectance of visible and near-infrared radiation from vegetation. Remote Sens. Environ. 1970, 1, 155–159. [Google Scholar] [CrossRef]
- Mackinney, G. ABSORPTION OF LIGHT BY CHLOROPHYLL SOLUTIONS. J. Biol. Chem. 1941, 140, 315–322. [Google Scholar] [CrossRef]
- Carter, G.A.; Cibula, W.G.; Miller, R.L. Narrow-band Reflectance Imagery Compared with ThermalImagery for Early Detection of Plant Stress. J. Plant Physiol. 1996, 148, 515–522. [Google Scholar] [CrossRef]
- Rullan-Silva, C.; Olthoff, A.; De La Mata, J.D.; Pajares-Alonso, J. Remote Monitoring of Forest Insect Defoliation -A Review-. For. Syst. 2013, 22, 377–391. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.R.K.; Nieberding, F.; Prinz, T.; Knoth, C. Analysis of Unmanned Aerial System-Based CIR Images in Forestry—A New Perspective to Monitor Pest Infestation Levels. Forest 2015, 6, 594–612. [Google Scholar] [CrossRef] [Green Version]
- Kanaskie, A.; Mcwilliams, M.; Overhulser, D.; Christian, R. Black Bear Damage to Forest Trees in Northwest Oregon: Aerial and Ground Surveys, 2000. Oregon Dep. For. Pest Manag. Rep 2001, 28, 1–29. [Google Scholar]
- Fassnacht, F.E.; Latifi, H.; Stereńczak, K.; Modzelewska, A.; Lefsky, M.; Waser, L.T.; Straub, C.; Ghosh, A. Review of studies on tree species classification from remotely sensed data. Remote Sens. Environ. 2016, 186, 64–87. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Ranson, K.J.; Fedotova, E.V. Spatial pattern of Siberian silkmoth outbreak and taiga mortality. Scand. J. For. Res. 2007, 22, 531–536. [Google Scholar] [CrossRef]
- Wulder, M.; White, J.; Bentz, B.; Alvarez, M.; Coops, N. Estimating the probability of mountain pine beetle red-attack damage. Remote Sens. Environ. 2006, 101, 150–166. [Google Scholar] [CrossRef]
- Zhang, C.; Kovacs, J.M. The application of small unmanned aerial systems for precision agriculture: A review. Precis. Agric. 2012, 13, 693–712. [Google Scholar] [CrossRef]
- Aasen, H.; Honkavaara, E.; Lucieer, A.; Zarco-Tejada, P.J. Quantitative Remote Sensing at Ultra-High Resolution with UAV Spectroscopy: A Review of Sensor Technology, Measurement Procedures, and Data Correction Workflows. Remote Sens. 2018, 10, 1091. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Imaging Sensor | Headwall Nano-Hyperspec |
|---|---|
| Spectral bands | 273 spectral bands from 398 to 1001 nm |
| Focal Length | 4.8 mm |
| FWHM | 6 nm |
| Bit depth | 12-bit |
| Spatial bands | 640 |
| Ground sampling distance (GSD) | 2.5 cm |
| Flight height | 90 m |
| Flight speed | 4 m/s |
| ID | Damage Class | ITCs | Pixels | Pixels/ITC |
|---|---|---|---|---|
| 1 | No stress | 45 | 188,752 | 4194 |
| 2 | Fresh Damage 1 | 25 | 68,695 | 2747 |
| 3 | Old Damage | 38 | 111,607 | 2937 |
| Vegetation Indices | Equation | Reference |
|---|---|---|
| Normalized Difference Vegetation Index (NDVI) | Rouse et al. [39] | |
| Modified Chlorophyll Absorption Ratio Index (MCARI) | Daughtry et al. [40] | |
| Red-edge Normalized Difference Vegetation Index (RENDVI) | Gitelson and Merzlyak [21] | |
| Plant Senescing Reflectance Index (PSRI) | Merzlyak et al. [21] | |
| Vogelmann “red edge” Index (VREI1)Normalized Channel Ratio (NCR) | Vogelmann et al. [41] | |
| Coops et al. [26] |
| Features | Accuracy (%) SVM | Kappa SVM | Accuracy (%) RF | Kappa RF |
|---|---|---|---|---|
| VNIR | 83.8 | 0.75 | 73.4 | 0.60 |
| VIs | 57.6 | 0.36 | 54.8 | 0.32 |
| λ685; λ750 | 49.6 | 0.24 | 43.1 | 0.15 |
| NDVI | 45 | 0.17 | 38.8 | 0.08 |
| MCARI | 33.9 | 0.09 | 36.5 | 0.04 |
| RENDVI | 47.4 | 0.21 | 42.3 | 0.13 |
| PSRI | 45.5 | 0.18 | 38.4 | 0.07 |
| VREI 1 | 45.8 | 0.18 | 37.8 | 0.06 |
| NCR | 45.1 | 0.26 | 38.1 | 0.09 |
| full | 78.1 | 0.67 | 77.9 | 0.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magstadt, S.; Gwenzi, D.; Madurapperuma, B. Can a Remote Sensing Approach with Hyperspectral Data Provide Early Detection and Mapping of Spatial Patterns of Black Bear Bark Stripping in Coast Redwoods? Forests 2021, 12, 378. https://doi.org/10.3390/f12030378
Magstadt S, Gwenzi D, Madurapperuma B. Can a Remote Sensing Approach with Hyperspectral Data Provide Early Detection and Mapping of Spatial Patterns of Black Bear Bark Stripping in Coast Redwoods? Forests. 2021; 12(3):378. https://doi.org/10.3390/f12030378
Chicago/Turabian StyleMagstadt, Shayne, David Gwenzi, and Buddhika Madurapperuma. 2021. "Can a Remote Sensing Approach with Hyperspectral Data Provide Early Detection and Mapping of Spatial Patterns of Black Bear Bark Stripping in Coast Redwoods?" Forests 12, no. 3: 378. https://doi.org/10.3390/f12030378