
Dynamics of Soil Nutrients, Microbial Community Structure, Enzymatic Activity, and Their Relationships along a Chronosequence of Pinus massoniana Plantations

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
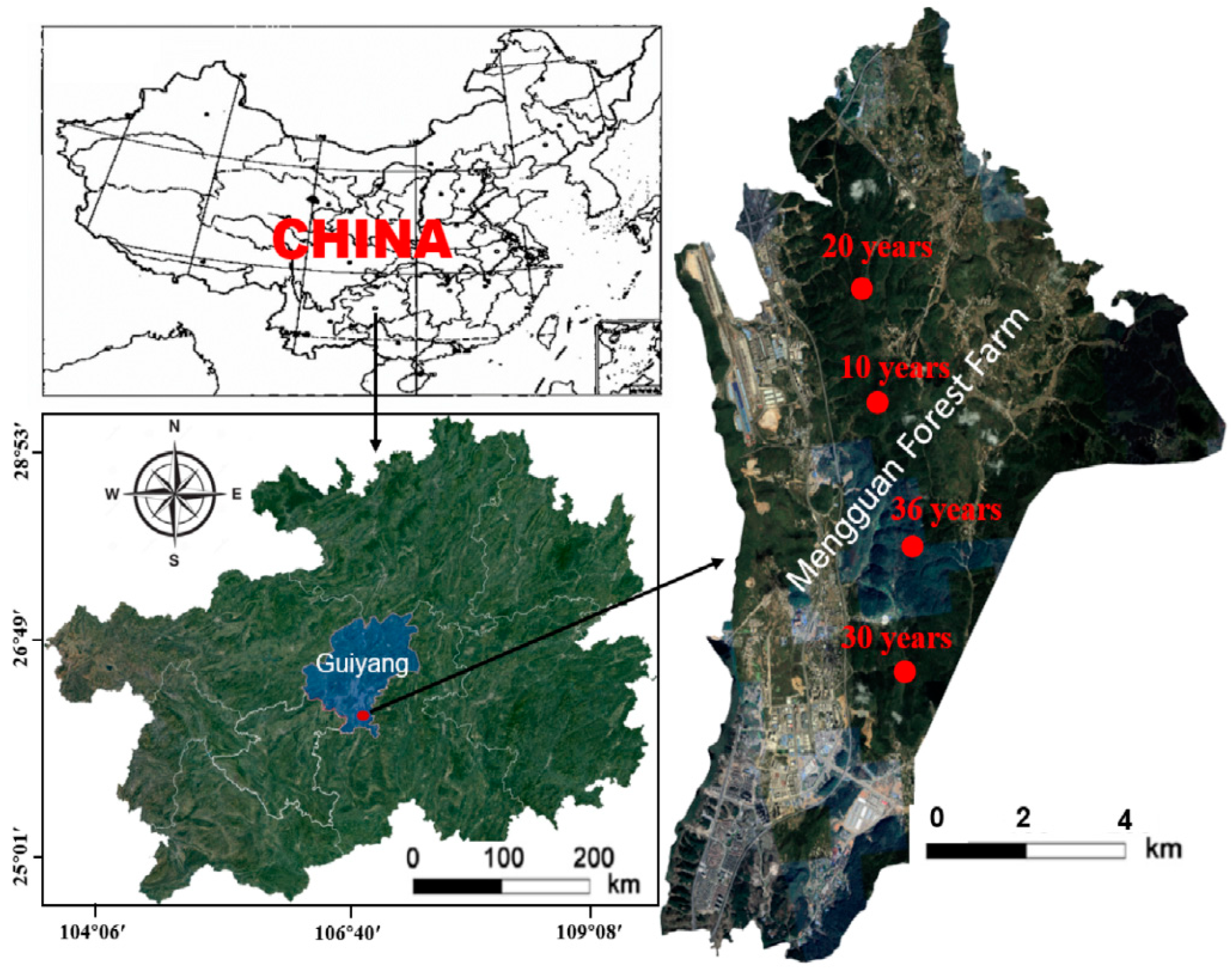
2.1. Study Region
2.2. Experimental Design
2.3. Soil Samples Collection
2.4. Soil Physicochemical Properties and Enzyme Activity Measurements
2.5. Determination of Soil Bacteria and Fungi
2.5.1. DNA Extraction, PCR Amplification, and Illumina Sequencing
2.5.2. Sequencing Data Processing
2.5.3. Operational Taxonomic Unit (OTU) Clustering and Notes
2.6. Statistical Analysis
3. Results
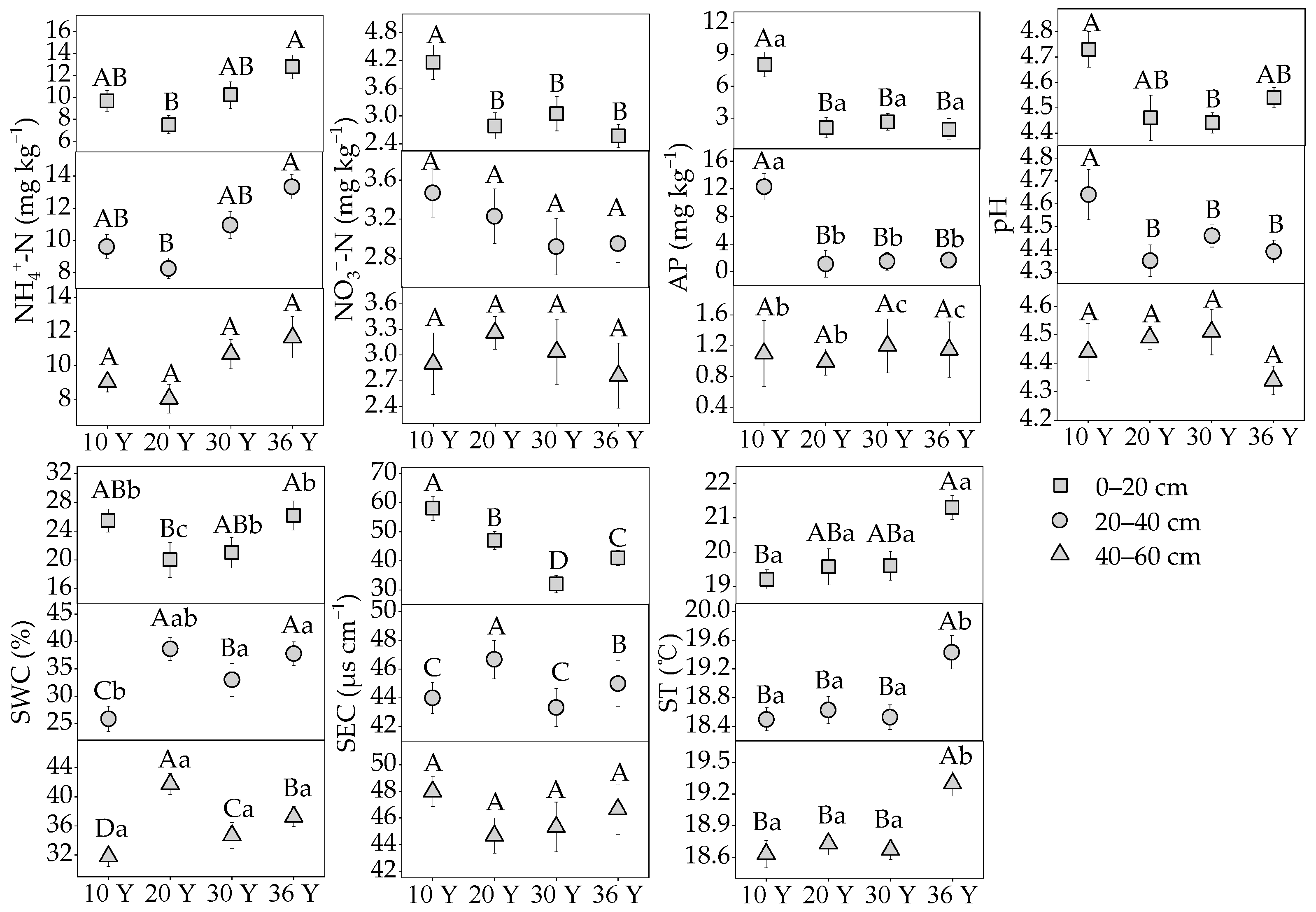
3.1. The Variation of Soil C: N: P Stoichiometry and Physicochemical Properties with Stand Age and Soil Depth
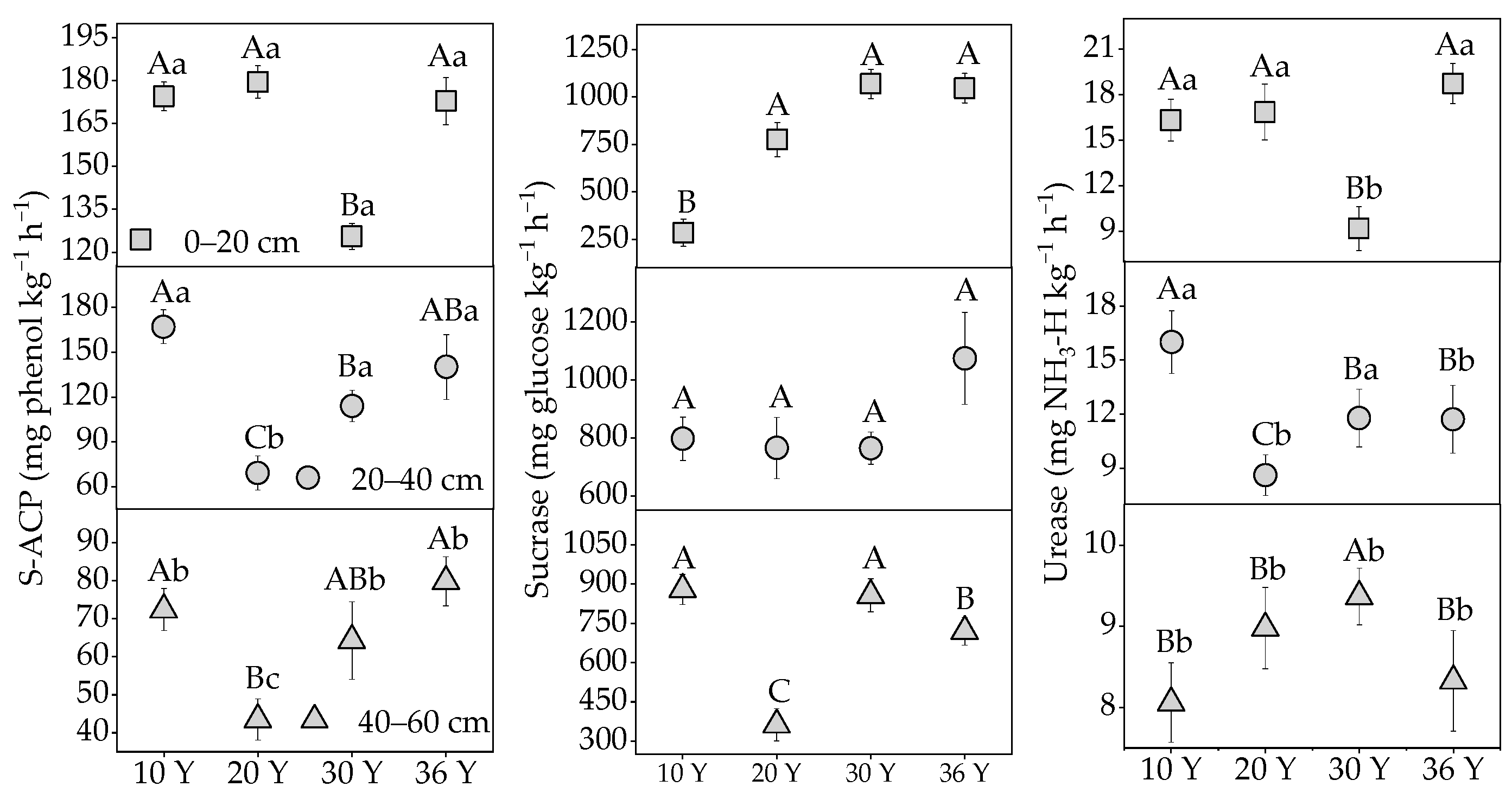
3.2. The Variation of Soil Enzyme Activities with Stand Age and Soil Depth
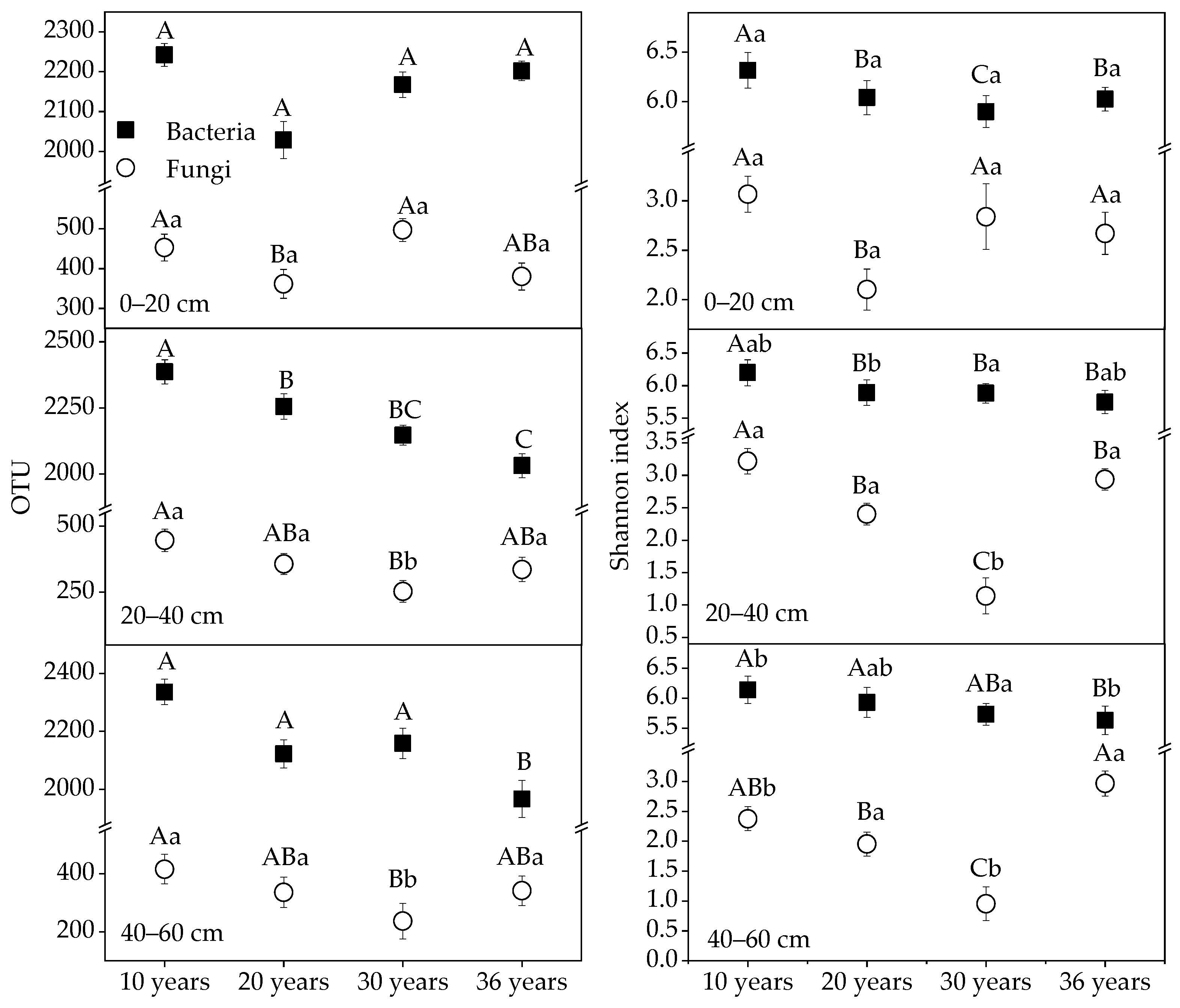
3.3. Alpha Diversity of Soil Bacteria and Fungi
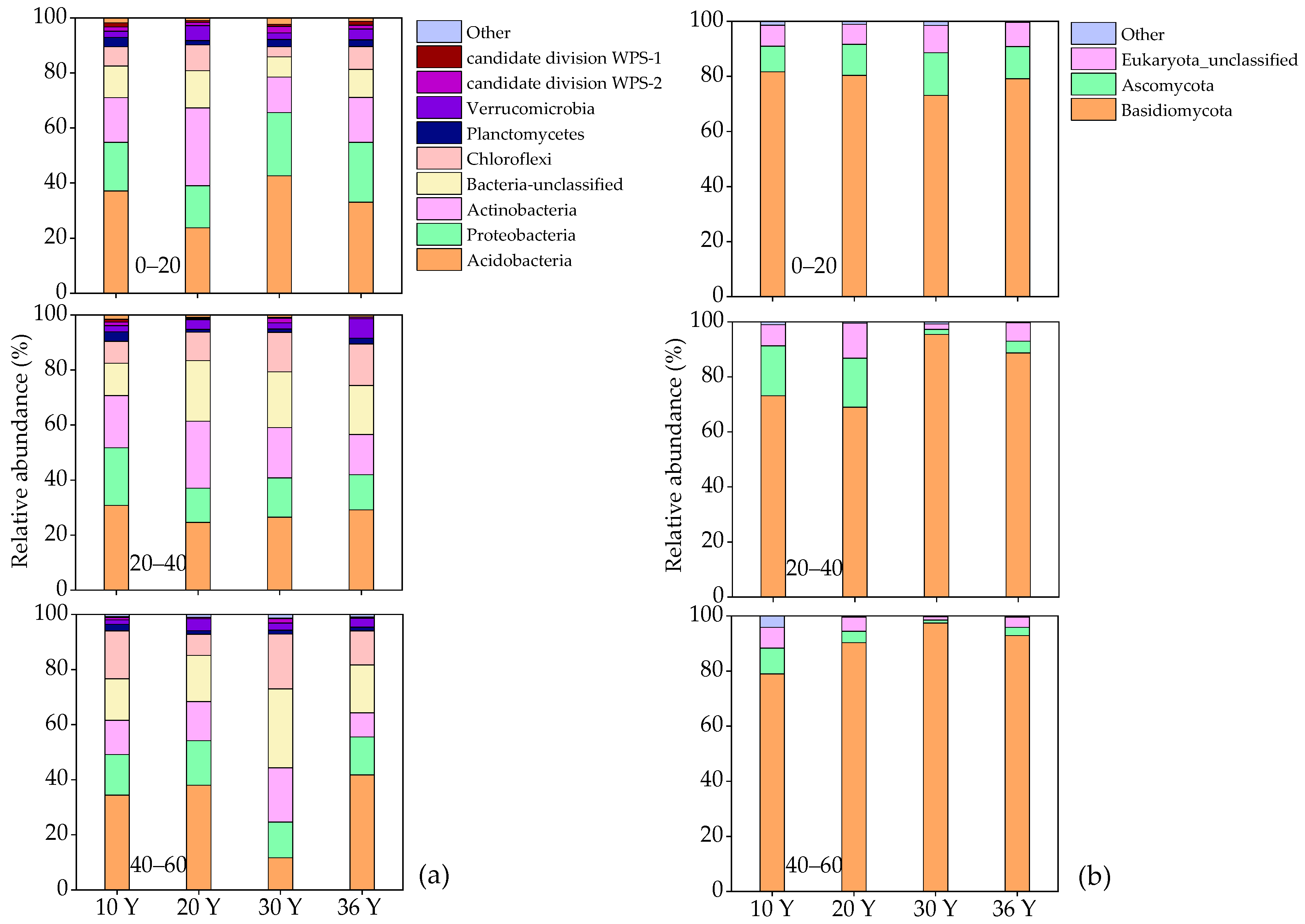
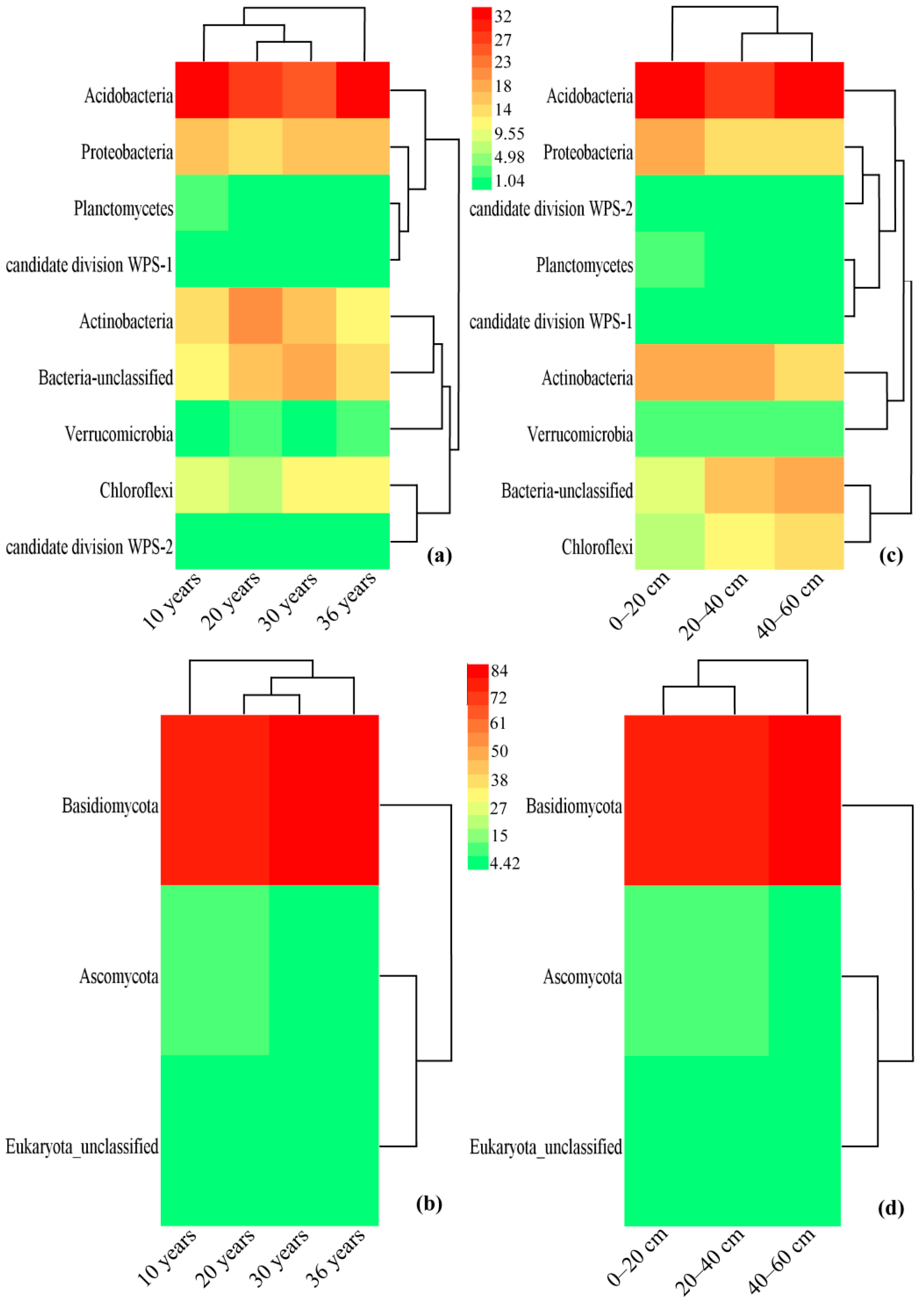
3.4. Community Composition of Bacteria and Fungi
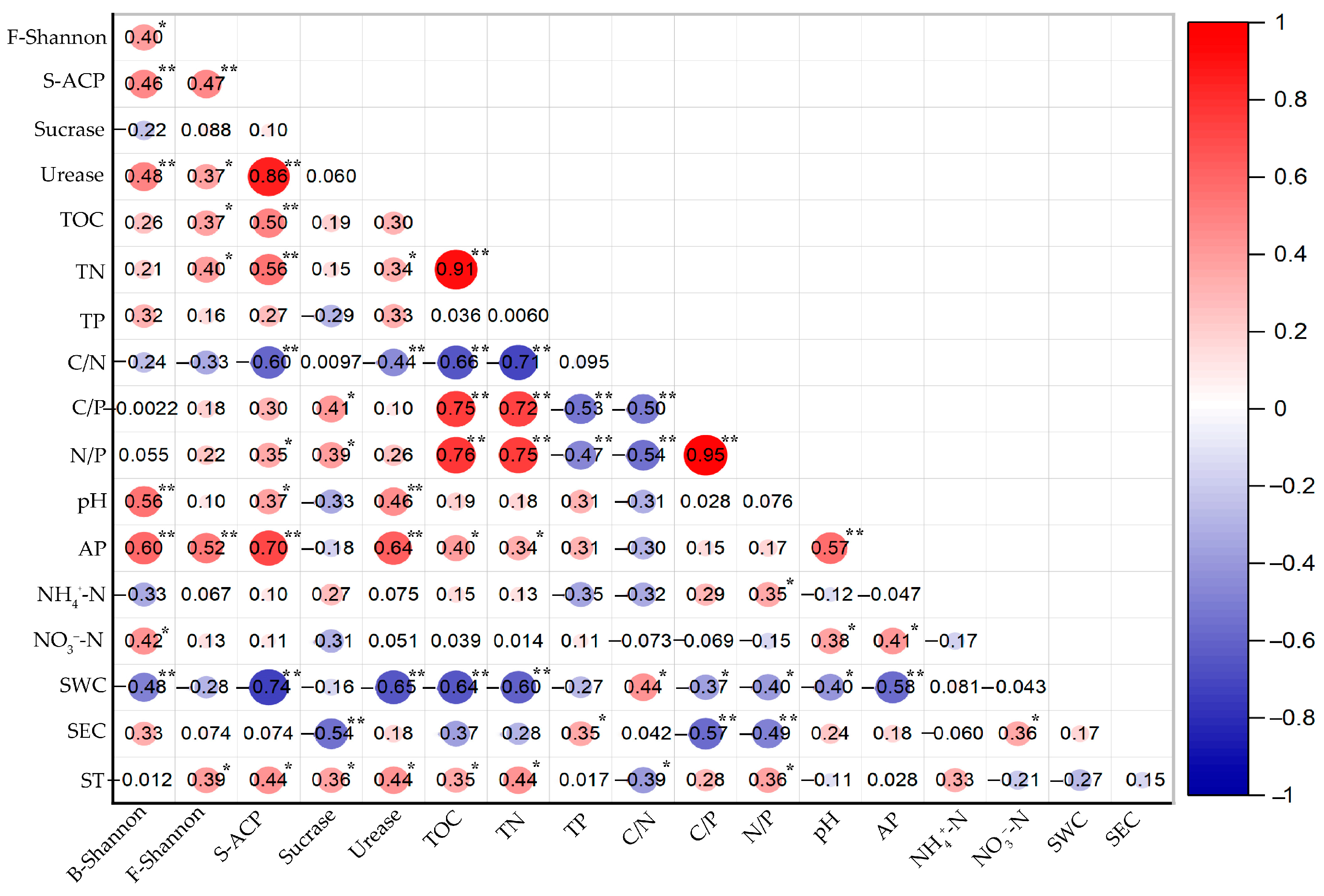
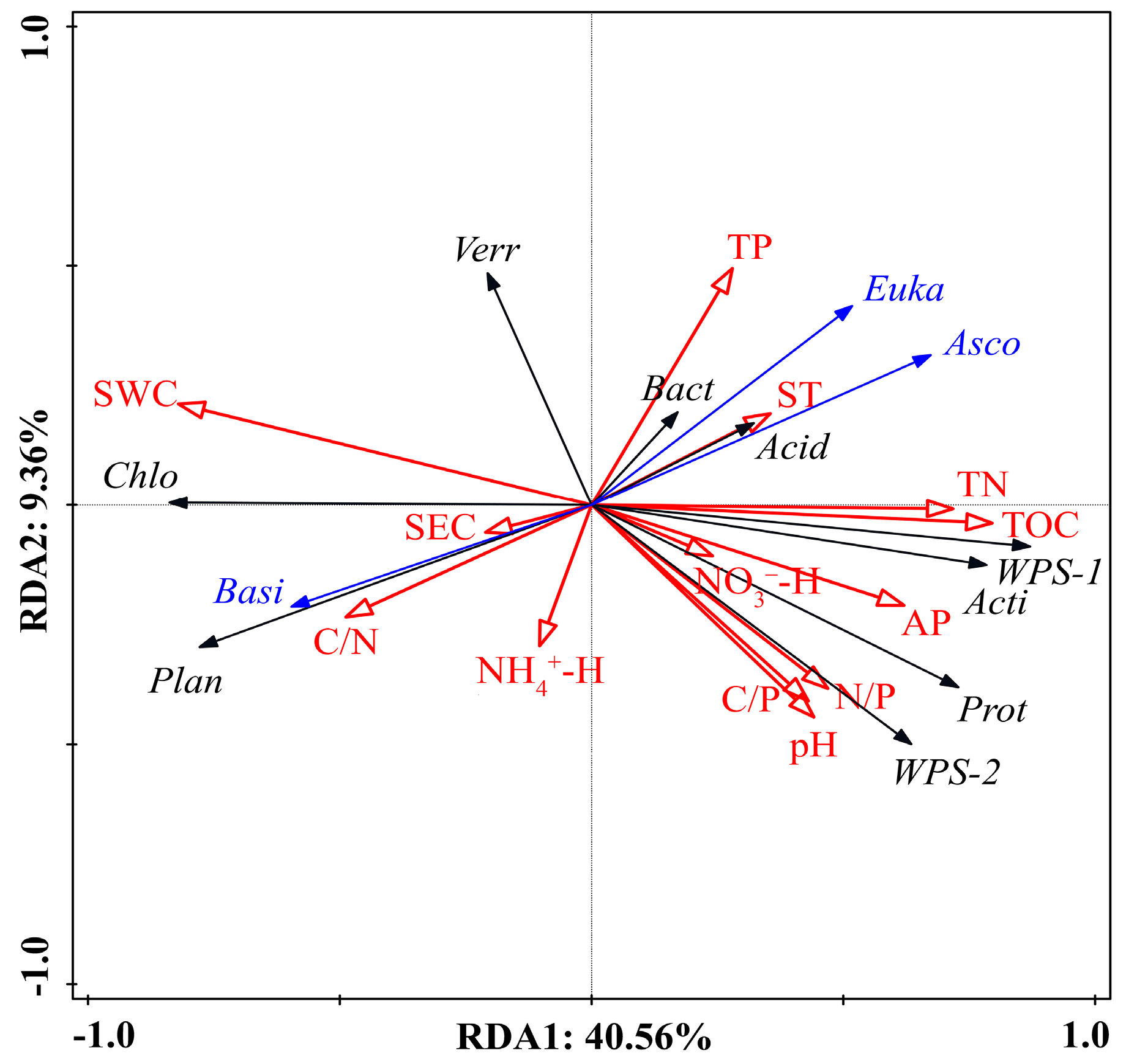
3.5. Relationship between Microbes, Enzyme Activity, and Soil Physicochemical Properties
4. Discussion
4.1. Effects of Soil Physicochemical Properties on Microbial Alpha Diversity
4.2. Effects of Soil Physicochemical Properties on Microbial Composition
4.3. Relationship among Soil Physicochemical Properties, Microbes, and Enzyme Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yang, H.; Wang, S.; Zhang, J.; Fan, B.; Zhang, W. Biomass and nutrients of Pinus massoniana plantations in southern China: Simulations for different management practices. J. Food Agric. Environ. 2011, 9, 689–693. [Google Scholar]
- SFA (State Forestry Administration P.R. China). National Forest Resources Statistics (2009–2014). NFI 8th; State Forestry Administration P.R: Bejing, China, 2014.
- Wen, L.; Lei, P.; Xiang, W.; Yan, W.; Liu, S. Soil microbial biomass carbon and nitrogen in pure and mixed stands of Pinus massoniana and Cinnamomum camphora differing in stand age. For. Ecol. Manag. 2014, 328, 150–158. [Google Scholar] [CrossRef]
- Chen, F.; Zheng, H.; Zhang, K.; Ouyang, Z.; Lan, J.; Li, H.; Shi, Q. Changes in soil microbial community structure and metabolic activity following conversion from native Pinus massoniana plantations to exotic Eucalyptus plantations. For. Ecol. Manag. 2013, 291, 65–72. [Google Scholar] [CrossRef]
- Wang, W.; Su, D.; Qiu, L.; Wang, H.; An, J.; Zheng, G.; Zu, Y. Concurrent changes in soil inorganic and organic carbon during the development of larch, Larix gmelinii, plantations and their effects on soil physicochemical properties. Environ. Earth Sci. 2013, 69, 1559–1570. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, X.; Chen, F.; Li, C.; Wu, L. Effects of the successive planting of Eucalyptus urophylla on soil bacterial and fungal community structure, diversity, microbial biomass, and enzyme activity. Land Degrad Dev. 2019, 1–11. [Google Scholar] [CrossRef]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N: P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Qiao, W.; Gao, D.; Dai, Y.; Deng, J.; Yang, G.; Han, X.; Ren, G. Relationship between soil nutrient properties and biological activities along a restoration chronosequence of Pinus tabulaeformis plantation forests in the Ziwuling Mountains, China. Catena 2018, 161, 85–95. [Google Scholar] [CrossRef]
- Ren, C.; Zhao, F.; Kang, D.; Yang, G.; Han, X.; Tong, X.; Feng, Y.; Ren, G. Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmland. For. Ecol. Manag. 2016, 376, 59–66. [Google Scholar] [CrossRef]
- Fageria, N.K. Soil quality vs. environmentally-based agricultural management practices. Commun. Soil Sci. Plant Anal. 2002, 33, 2301–2329. [Google Scholar] [CrossRef]
- Minasny, B.; Hong, S.Y.; Hartemink, A.E. Soil pH increase under paddy in South Korea between 2000 and 2012. Agric. Ecosyst. Environ. 2016, 221, 205–213. [Google Scholar] [CrossRef]
- Mayakaduwage, S.; Mosley, L.M.; Marschner, P. Phosphorus pools in acid sulfate soil are influenced by soil water content and form in which P is added. Geoderma 2021, 381, 114692. [Google Scholar] [CrossRef]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant–microbial–soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef] [Green Version]
- Powlson, D.S.; Jenkinson, D.S. A comparison of the organic matter, biomass, adenosine triphosphate and mineralizable nitrogen contents of ploughed and direct-drilled soils. J. Agric. Sci. 1981, 97, 713–721. [Google Scholar] [CrossRef]
- Van, D.H.M.G.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar]
- Zhang, C.; Liu, G.B.; Xue, S.; Wang, G.L. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of bandoned farmland in the Loess Plateay. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Tian, J.; He, N.; Hale, L.; Niu, S.; Yu, G.; Liu, Y.; Blagodatskaya, E.; Kuzyakov, Y.; Gao, Q.; Zhou, J. Soil organic matter availability and climate drive latitudinal patterns in bacterial diversity from tropical to cold-temperate forests. Funct. Ecol. 2018, 32, 61–70. [Google Scholar] [CrossRef]
- Lauber, C.L.; Ramirez, K.S.; Aanderud, Z.; Lennon, J.; Fierer, N. Temporal variability in soil microbial communities across land-use types. ISME J. 2013, 7, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Huang, Z.; He, Z.; Yu, Z.; Wang, M.; Davis, M.R.; Yang, Y. Soil C:N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations. Plant Soil. 2015, 387, 103–116. [Google Scholar] [CrossRef]
- Ren, C.; Chen, J.; Deng, J.; Zhao, F.; Han, X.; Yang, G.; Tong, X.; Feng, Y.; Shelton, S.; Ren, G. Response of microbial diversity to C:N:P stoichiometry in fine root and microbial biomass following afforestation. Biol. Fertil. Soils 2017, 53, 457–468. [Google Scholar] [CrossRef]
- Fang, F.; Wu, C.Z.; Hong, W.; Fan, H.L.; Song, P. Study on the ralationship between rhizospheric or non-rhizospheric soil enzyme and microbe in different plants. Subtrop. Agric. Reasearch. 2007, 3, 209–215. [Google Scholar]
- Adamczyk, B.; Kilpeläinen, P.; Kitunen, V.H.; Smolander, A. Potential activities of enzymes involved in N, C, P and S cycling in boreal forest soil under different tree species. Pedobiologia 2014, 57, 97–102. [Google Scholar] [CrossRef]
- Xu, Z.; Yu, G.; Zhang, X.; He, N.; Wang, Q.; Wang, S.; Wang, R.; Zhao, N.; Jia, Y.; Wang, C. Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China (NSTEC). Soil Biol. Biochem. 2017, 104, 152–163. [Google Scholar] [CrossRef]
- Stock, S.C.; Kster, M.; Dippold, M.A.; Nájera, F.; Kuzyakov, Y. Environmental drivers and stoichiometric constraints on enzyme activities in soils from rhizosphere to continental scale. Geoderma 2019, 337, 973–982. [Google Scholar] [CrossRef]
- Zhang, C.B.; Jin, Z.X.; Ke, S.X. Study on the relationship among enzymes activities, microorganism, respiration rate and physico-chemical properties of soil under different forests of Tiantai mountain. J. Plant Nutr. Fertil. 2004, 10, 51–56. [Google Scholar]
- Burns, R.G.; Deforest, J.L.; Marxsen, J.R.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Hedo, J.; Cerda, A.; Candel-Perez, D.; Vinegla, B. Unravelling the importance of forest age stand and forest structure driving microbiological soil properties, enzymatic activities and soil nutrients content in Mediterranean Spanish black pine(Pinus nigra Ar. ssp. salzmannii) Forest. Sci. Total Environ. 2016, 562, 145–154. [Google Scholar] [CrossRef]
- Shen, F.; Wu, J.; Fan, H.; Liu, W.; Guo, X.; Duan, H.; Hu, L.; Lei, X.; Wei, X. Soil N/P and C/P ratio regulate the responses of soil microbial community composition and enzyme activities in a long-term nitrogen loaded Chinese fir forest. Plant Soil. 2019, 436, 91–107. [Google Scholar] [CrossRef]
- Li, B.; Ge, T.; Xiao, H.; Zhu, Z.; Li, Y.; Shibistova, O.; Liu, S.; Wu, J.; Inubushi, K.; Guggenberger, G. Phosphorus content as a function of soil aggregate size and paddy cultivation in highly weathered soils. Environ. Sci. Pollut. Res. Int. 2016, 23, 7494–7503. [Google Scholar] [CrossRef]
- NFA (National Forestry Administration). Regulations for Age-Class and Age-Group Division of Main Tree-Species; Wang, H.C., Li, G.C., Luan, G.J., Wang, R.N., Wang, H.Z., Bian, F., Shi, J., Zhou, J., Wang, Q., Gao, Z.F., et al., Eds.; National Forestry Administration: Beijing, China, 2017. Available online: http://www.doc88.com/p-5671706807327.html (accessed on 27 October 2017).
- Bao, S. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Shi, Y.; Shen, Q.R.; Mao, Z.S.; Xu, G.H. Time and Horizontal Spatial Variations of NH4+-N and NO3−-N of Rhizospheric Soil with RiceCultivation on Upland Condition Mulched with Half-decomposed Rice Straw. Sci. Agric. Sin. 2002, 5, 520–524. [Google Scholar]
- Norman, R.J.; Stucki, J.W. The Determination of Nitrate and Nitrite in Soil Extracts by Ultraviolet Spectrophotometry. Soil Sci. Soc. Am. J. 1981, 45, 347–353. [Google Scholar] [CrossRef]
- Pfahler, V.; Bielnicka, A.; Smith, A.C.; Granger, S.J.; Turner, B.L. A rapid ammonium fluoride method to determine the oxygen isotope ratio of available phosphorus in tropical soils. Rapid Commun. Mass Spectrom. 2020, 34, e8647. [Google Scholar] [CrossRef] [Green Version]
- Guan, S.Y. Soil Enzyme and Its Research Methods; China Agricultural press: Beijing, China, 1986. [Google Scholar]
- Hugerth, L.W.; Wefer, H.A.; Lundin, S.; Jakobsson, H.E.; Lindberg, M.; Rodin, S.; Engstrand, L.; Andersson, A.F. DegePrime, a Program for Degenerate Primer Design for Broad-Taxonomic-Range PCR in Microbial Ecology Studies. Appl. Environ. Microbiol. 2014, 80, 5116–5123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, E.; Leeflang, P.; Glandorf, B.; Van Elsas, J.D.; Wernars, K. Analyis of fungal diversity in the wheat rhizosphere by sequencing of cloned PCR-amplified genes encoding 18S rRNA and temperature gradient gel electrophoresis. Appl. Environ. Microbiol. 1999, 65, 2614–2621. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Liu, N.; Zhang, Y. Soil aggregates regulate the impact of soil bacterial and fungal communities on soil respiration. Geoderma 2019, 337, 444–452. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids. Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Elizabeth, H.S.; Wenxiong, L.; Bailian, L.; Linkun, W.; Changxun, F.; Zhixing, Z.; Zhen, J. Soil Microbial Community Structure and Metabolic Activity of Pinus elliottii Plantations across Different Stand Ages in a Subtropical Area. PLoS ONE. 2015, 10, e0135354. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zang, H.; Tida, G.; Bai, J.; Guggenberger, G. Intensive fertilization (N, P, K, Ca, and S) decreases organic matter decomposition in paddy soil. Appl. Soil Ecol. 2018, 127, 51–57. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, P.B.; Khachane, A.N.; Campbell, C.D.; Thomas, N.; Freitag, T.E.; Al-Soud, W.A.; Sørensen, S.; Bardgett, R.D.; Singh, B.K. It is elemental: Soil nutrient stoichiometry drives bacterial diversity. Environ. Microbiol. 2017, 19, 1176–1188. [Google Scholar] [CrossRef] [PubMed]
- Högberg, M.N.; Högberg, P.; Myrold, D.D. Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three? Oecologia 2007, 150, 590–601. [Google Scholar] [CrossRef]
- Shi, P.L.; Zhang, X.Z. Some advance in the main factors controlling soil respiration. Adv. Earth Sci. 2005, 20, 778–785. [Google Scholar]
- Kjaller, A.; Struwe, S. Microfungi in ecosystems: Fungal occurrence and activity in litter and soil. Oikos 1982, 39, 389–442. [Google Scholar] [CrossRef]
- Guo, P.; Jiang, H.; Yu, S.; Ma, Y.; Dou, R.; Song, X. Comparison of Litter Decomposition of Six Species of Coniferous and Broad-leaved Trees in Subtropical China. Chin. J. Appl. Environ. Biol. 2009, 15, 655–659. [Google Scholar]
- Kubartová, A.; Ranger, J.; Berthelin, J.; Beguiristain, T. Diversity and Decomposing Ability of Saprophytic Fungi from Temperate Forest Litter. Microb. Ecol. 2009, 58, 98–107. [Google Scholar] [CrossRef]
- Du, C.; Geng, Z.; Wang, Q.; Zhang, T.; Wang, Y. Variations in bacterial and fungal communities through soil depth profiles in a Betula albosinensis forest. J. Microbiol. 2017, 55, 684–693. [Google Scholar] [CrossRef]
- Izumi, H.; Cairney, J.W.G.; Killham, K.; Moore, E.; Alexander, I.J.; Anderson, I.C. Bacteria associated with ectomycorrhizas of slash pine (Pinus elliotti) in south-eastern Queensland, Australia. FEMS Microbiol. Lett. 2008, 282, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and Associated Fungi Drive Long-Term Carbon Sequestration in Boreal Forest. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef]
- Zhang, Y.; Cong, J.; Lu, H.; Li, G.; Qu, Y.; Su, X.; Zhou, J.; Li, D. Community structure and elevational diversity patterns of soil Acidobacteria. J. Environ. Sci. 2014, 26, 1717–1724. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J. Growth rate–stoichiometry couplings in diverse biota. Ecol. Lett. 2010, 6, 936–943. [Google Scholar] [CrossRef] [Green Version]
- Kopecky, J.; Kyselkova, M.; Omelka, M.; Cermak, L.; Novotna, J.; Grundmann, G.L.; Moënne-Loccoz, Y.; Sagova-Mareckova, M. Actinobacterial community dominated by a distinct clade in acidic soil of a waterlogged deciduous forest. FEMS Microbiol. Ecol. 2011, 78, 386–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santalahti, M.; Sun, H.; Jumpponen, A.; Pennanen, T.; Heinonsalo, J. Vertical and seasonal dynamics of fungal communities in boreal Scots pine forest soil. FEMS Microbiol. Ecol. 2016, 92, fiw170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, R.M.; Wang, R.L.; Xiao, W.F.; Feng, X.H.; Liu, Z.B.; Ge, X.G.; Wang, X.R.; Zhang, W.Y. Spatial distribution of root biomass of Pinus massoniana plantation in Three Gorges Reservoir area, China. Acta Ecol. Sin. 2012, 32, 823–832. [Google Scholar] [CrossRef] [Green Version]
- He, Y.H.; Zhou, G.Y.; Wang, S.J.; Li, H. Fungal diversity in Cunninghamia lanceolata plantation soil. Acta Ecol. Sin. 2014, 34, 2725–2736. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter List | 10 Years | 20 Years | 30 Years | 36 Years |
|---|---|---|---|---|
| Altitude (m) | 1194 | 1175 | 1206 | 1214 |
| Slope aspect | N/EN | WN | N | EN |
| Slope (°) | 5 | 6 | 5 | 8 |
| Slope position | Downhill | Downhill | Downhill/Middle slope | Middle slope |
| Soil texture | Sandy loam | Sandy loam | Sandy loam | Sandy loam |
| Soil parent rock | Sandy shale | Sandy shale | Sandy shale | Sandy shale |
| Forest stand canopy density | 0.90 ± 0.07 a | 0.85 ± 0.10 ab | 0.75 ± 0.09 b | 0.80 ± 0.08 ab |
| Forest stand density (trees ha−1) | 4675 ± 256 a | 2812 ± 135 b | 1356 ± 103 c | 1083 ± 94 d |
| Average DBH (cm) | 8.53 ± 1.56 d | 12.66 ± 2.55 c | 18.46 ± 3.01 b | 21.26 ± 5.26 a |
| Average tree height (m) | 7.43 ± 1.83 d | 15.64 ± 2.74 c | 17.94 ± 2.43 b | 20.87 ± 3.26 a |
| Variables | F (p) Value | ||
|---|---|---|---|
| A | D | A × D | |
| TOC | 22.857 (<0.01) | 158.197 (<0.01) | 14.171 (<0.01) |
| TN | 8.122 (<0.01) | 68.933 (<0.01) | 5.745 (<0.01) |
| TP | 10.625 (<0.01) | 4.283 (0.026) | 1.520 (0.214) |
| C/N | 3.916 (0.021) | 34.720 (<0.01) | 1.525 (0.213) |
| C/P | 81.256 (<0.01) | 46.031 (<0.01) | 17.106 (<0.01) |
| N/P | 83.850 (<0.01) | 74.682 (<0.01) | 18.641 (<0.01) |
| pH | 5.162 (<0.01) | 2.950 (0.072) | 2.548 (0.047) |
| AP | 481.016 (<0.01) | 223.540 (<0.01) | 154.060 (<0.01) |
| NH4+-N | 7.864 (<0.01) | 0.333 (0.720) | 0.148 (0.988) |
| NO3−-N | 3.852 (0.022) | 0.220 (0.804) | 2.237 (0.074) |
| SWC | 18.574 (<0.01) | 138.262 (<0.01) | 12.449 (<0.01) |
| SEC | 38.255 (<0.01) | 2.493 (0.104) | 28.426 (<0.01) |
| ST | 7.196 (<0.01) | 12.471 (<0.01) | 0.843 (0.550) |
| S-ACP | 14.595 (<0.01) | 111.858 (<0.01) | 7.782 (<0.01) |
| Sucrase | 5.564 (<0.01) | 1.523 (0.238) | 4.751 (<0.01) |
| Urease | 18.137 (<0.01) | 116.335 (<0.01) | 22.932 (<0.01) |
| B-OTU | 5.960 (<0.01) | 0.661 (0.526) | 1.667 (0.173) |
| B-Shannon | 27.456 (<0.01) | 11.752 (<0.01) | 1.448 (0.238) |
| F-OTU | 5.745 (<0.01) | 7.696 (<0.01) | 3.575 (0.011) |
| F-Shannon | 28.070 (<0.01) | 9.711 (<0.01) | 8.495 (<0.01) |
| Variables | Explains (%) | F-Value | p-Value |
|---|---|---|---|
| SWC | 28.2 | 13.4 | 0.002 ** |
| TOC | 7 | 3.6 | 0.012 * |
| AP | 6.2 | 3.4 | 0.016 * |
| C/P | 5.7 | 3.4 | 0.014 * |
| ST | 3.1 | 1.9 | 0.082 |
| SEC | 3 | 1.8 | 0.11 |
| TN | 2 | 1.2 | 0.288 |
| NH4+-N | 1.9 | 1.2 | 0.338 |
| C/N | 1.7 | 1.1 | 0.378 |
| pH | 1.8 | 1.1 | 0.344 |
| TP | 1.2 | 0.8 | 0.518 |
| NO3−-N | 0.7 | 0.5 | 0.798 |
| N/P | 0.7 | 0.4 | 0.834 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, J.; Guo, Q.; Li, H.; Luo, S.; Zhang, Y.; Yao, S.; Fan, X.; Sun, X.; Qi, Y. Dynamics of Soil Nutrients, Microbial Community Structure, Enzymatic Activity, and Their Relationships along a Chronosequence of Pinus massoniana Plantations. Forests 2021, 12, 376. https://doi.org/10.3390/f12030376
Pan J, Guo Q, Li H, Luo S, Zhang Y, Yao S, Fan X, Sun X, Qi Y. Dynamics of Soil Nutrients, Microbial Community Structure, Enzymatic Activity, and Their Relationships along a Chronosequence of Pinus massoniana Plantations. Forests. 2021; 12(3):376. https://doi.org/10.3390/f12030376
Chicago/Turabian StylePan, Jinwen, Qiqiang Guo, Huie Li, Siqiong Luo, Yaqin Zhang, Shan Yao, Xin Fan, Xueguang Sun, and Yujiao Qi. 2021. "Dynamics of Soil Nutrients, Microbial Community Structure, Enzymatic Activity, and Their Relationships along a Chronosequence of Pinus massoniana Plantations" Forests 12, no. 3: 376. https://doi.org/10.3390/f12030376