
Seasonal Stabilities of Soil Nematode Communities and Their Relationships with Environmental Factors in Different Temperate Forest Types on the Chinese Loess Plateau

Abstract
:1. Introduction
2. Materials and Methods
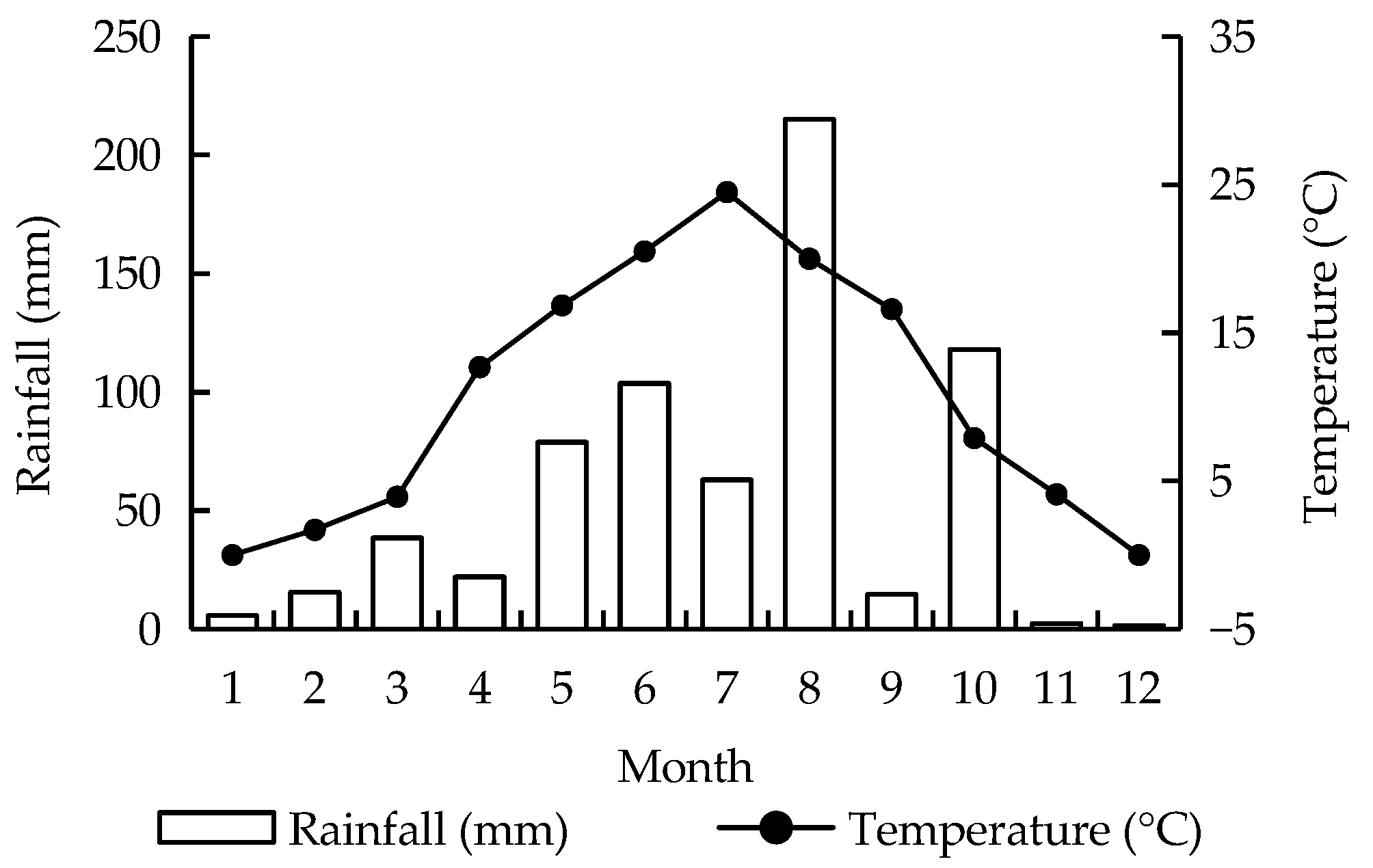
2.1. Study Site
2.2. Soil Sampling
2.3. Soil Physicochemical Analysis
2.4. Nematode Extraction and Identification
2.5. Nematode Community Analysis
2.6. Data Analysis
3. Results
3.1. Vegetation Characteristics and Soil Physicochemical Properties
3.2. Composition and Structure of Soil Nematode Communities
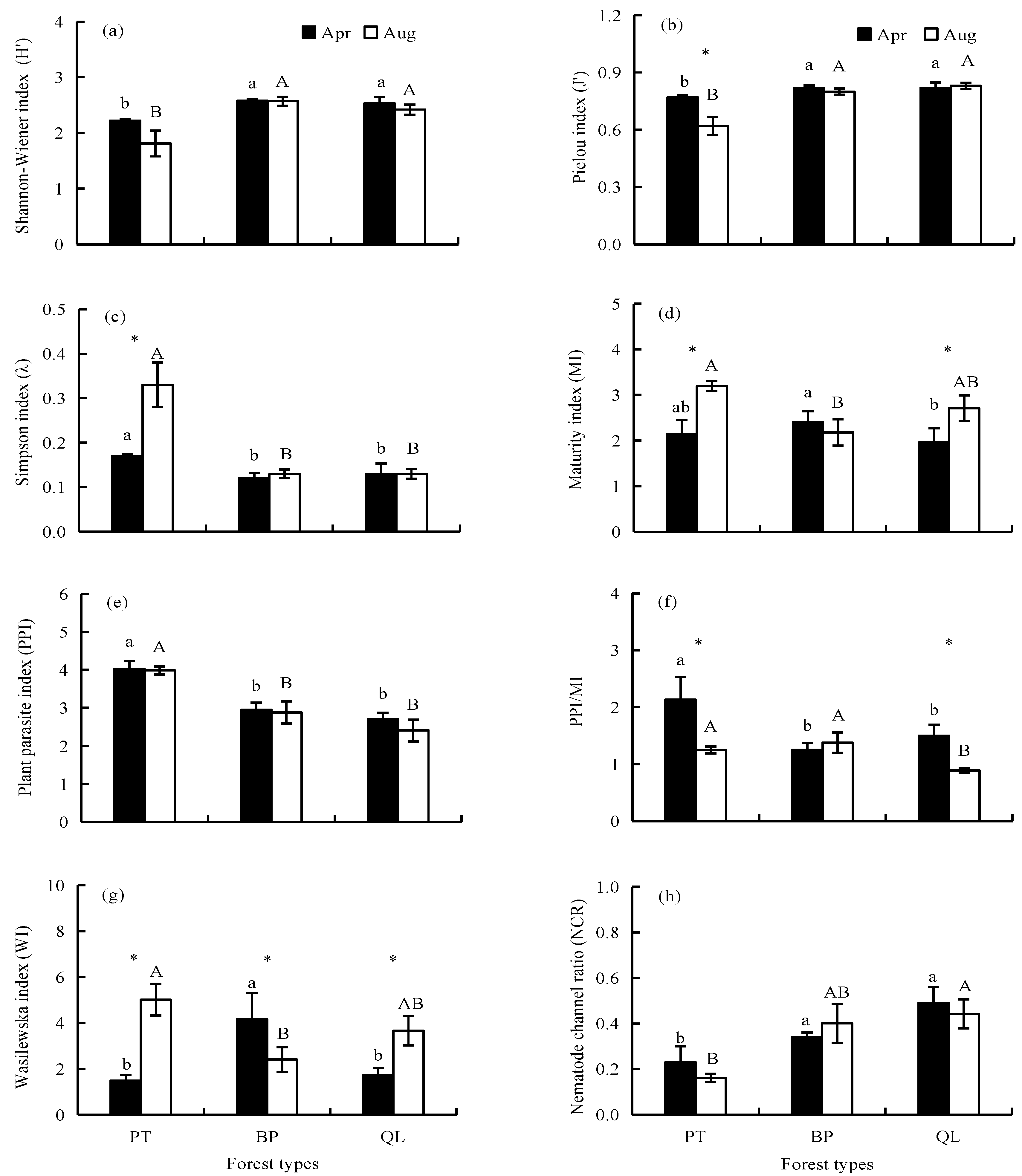
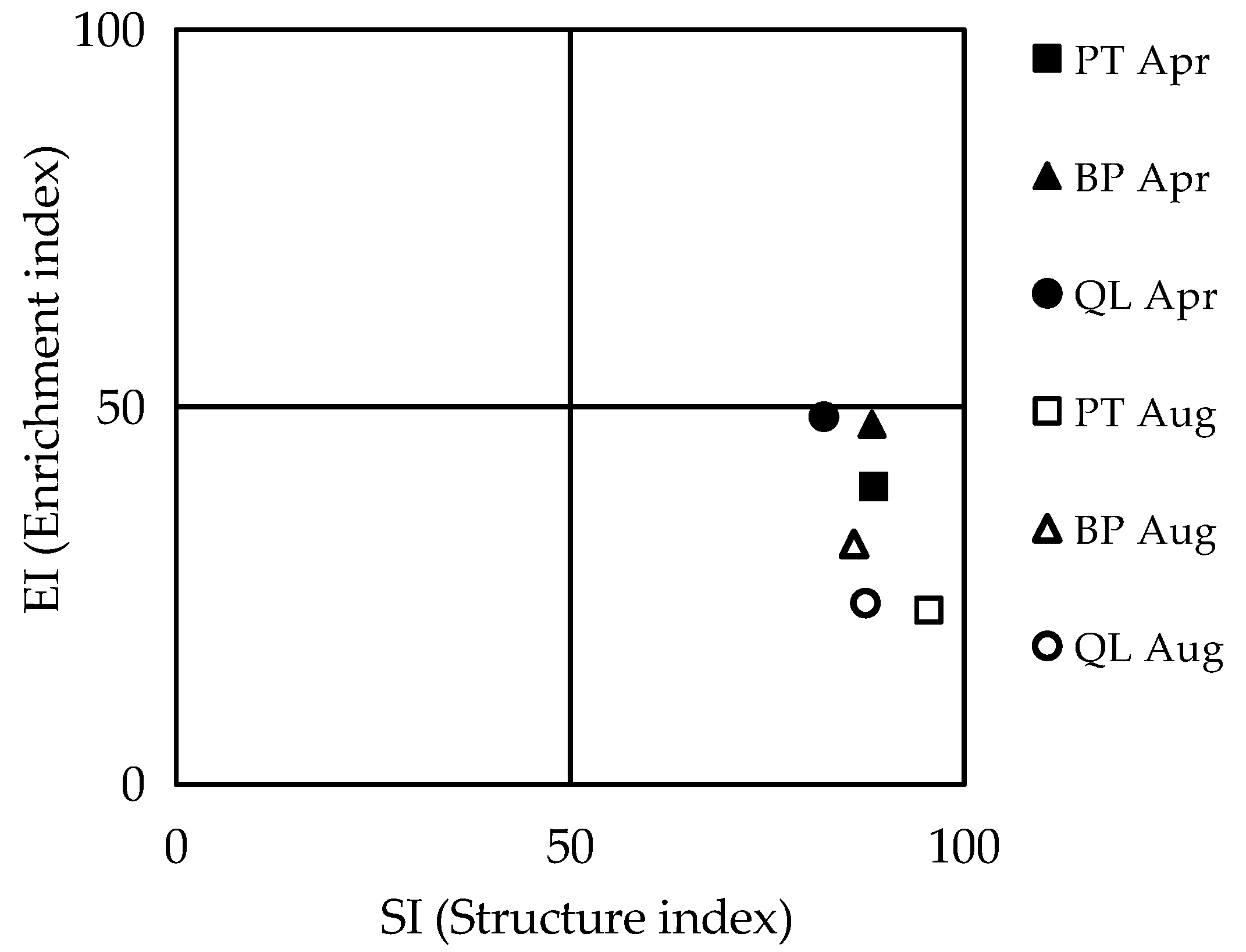
3.3. Ecological Indices of Soil Nematode Communities
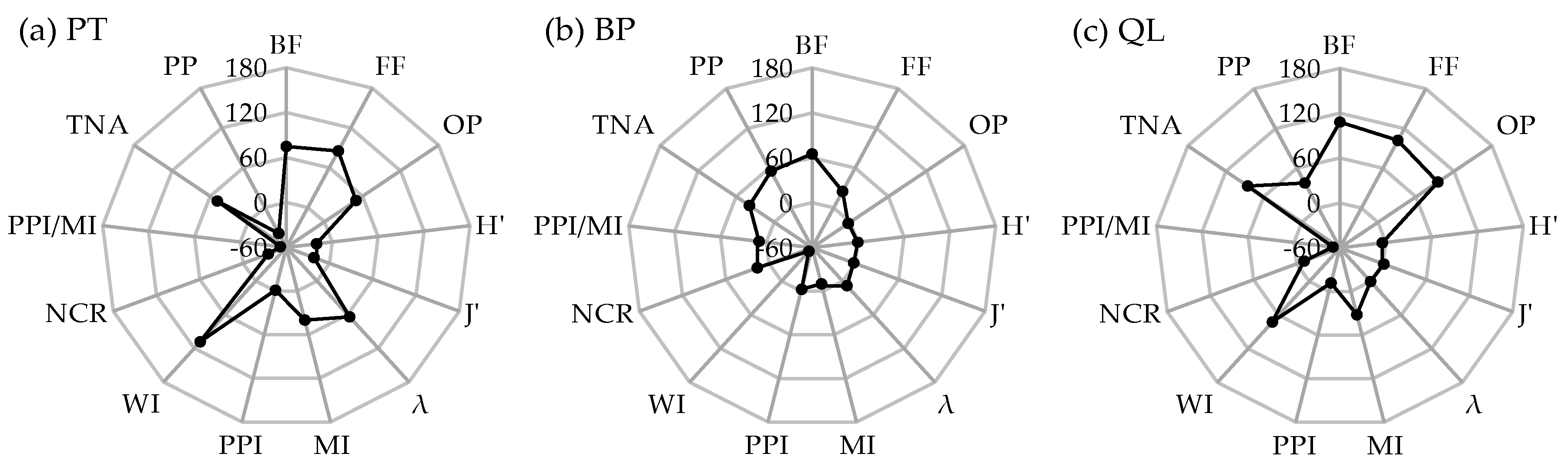
3.4. Seasonal Stability of Soil Nematode Communities under Different Forests
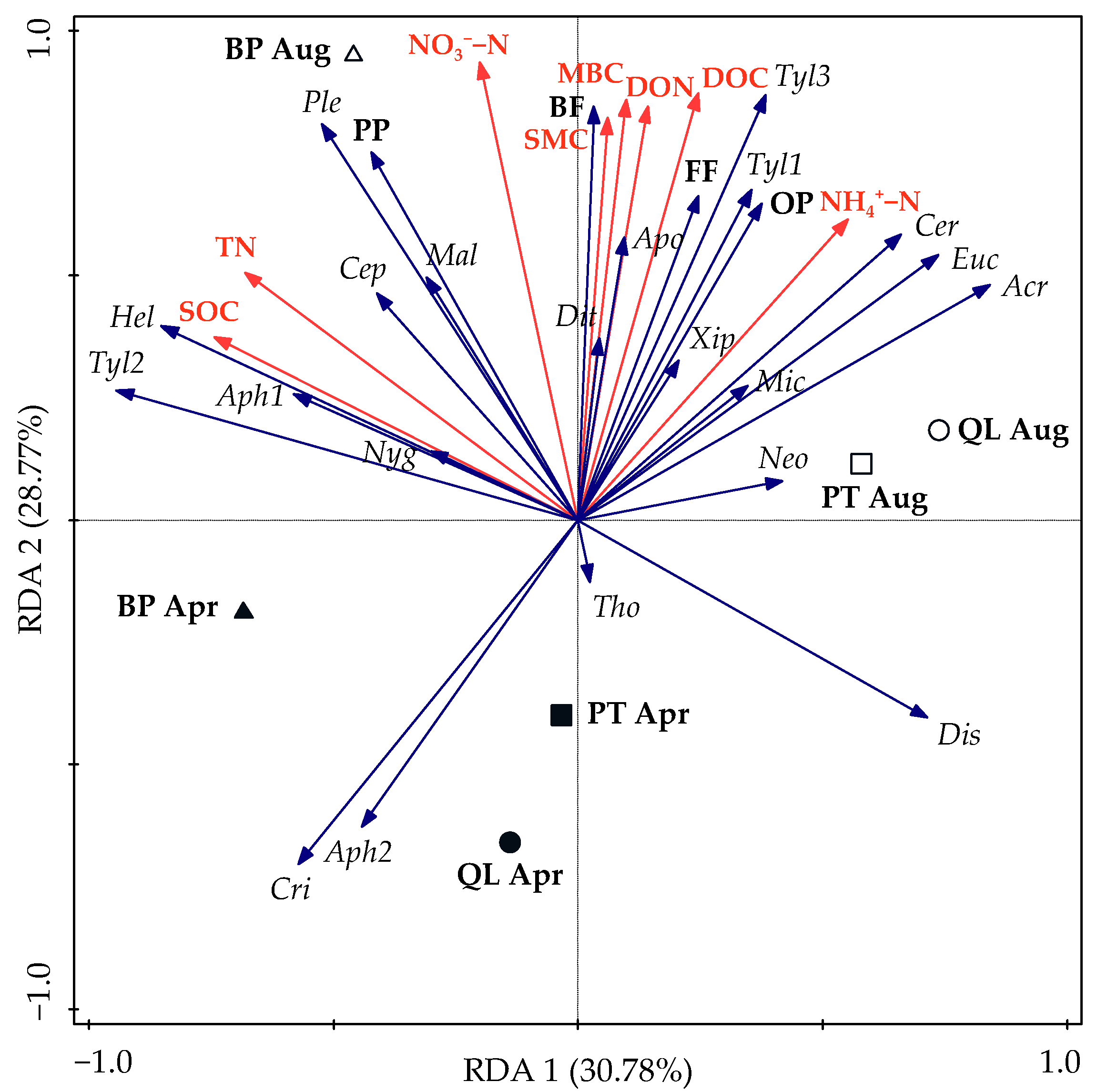
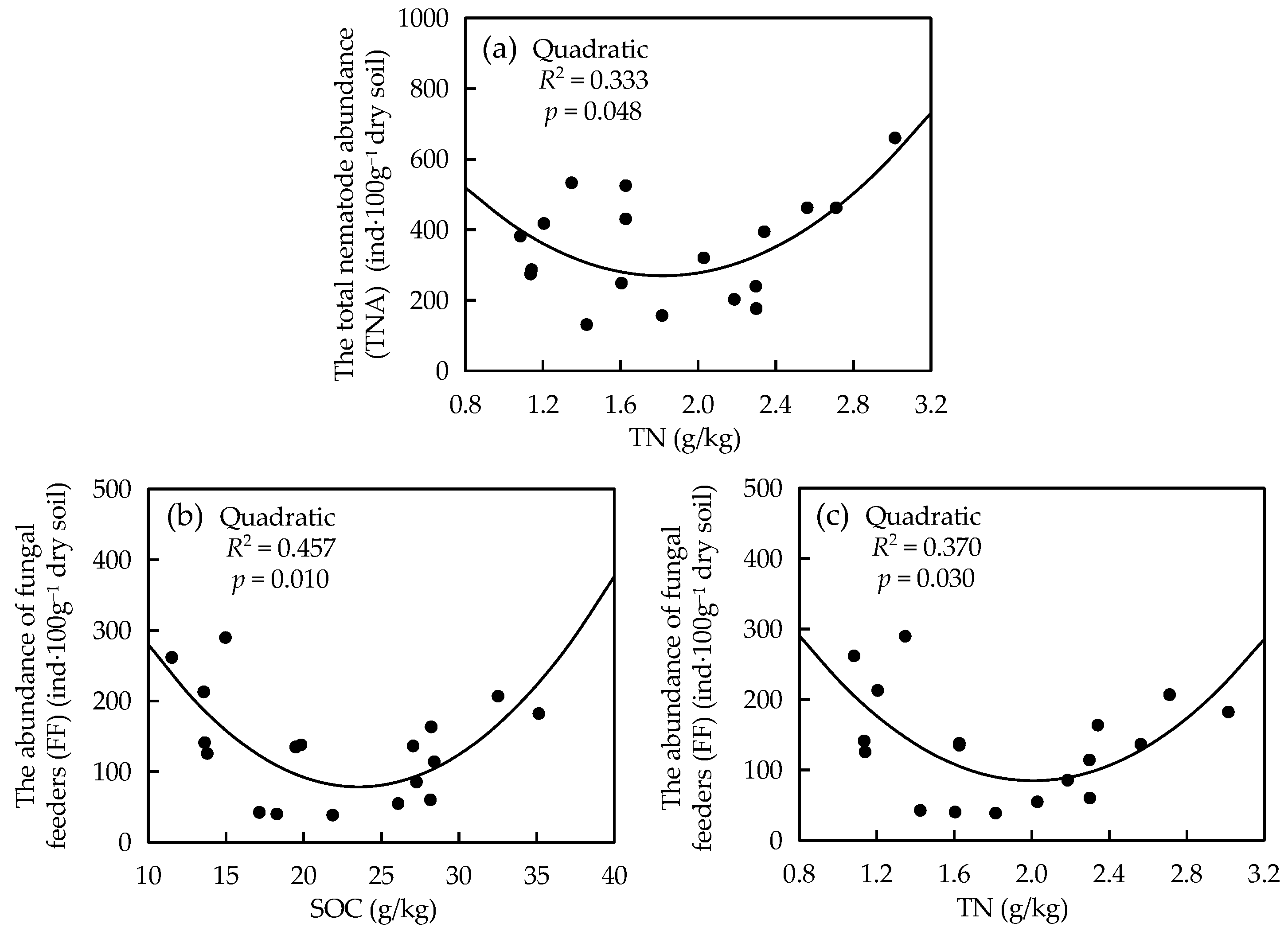
3.5. Relationship among Soil Nematode Communities and Soil Properties
4. Discussion
4.1. Effects of Forest Types on the Soil Nematode Community Characteristics
4.2. Effects of Forest Type on the Seasonal Stability of Soil Nematode Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; Degoede, R.; Freckman, D.W.; Georgieva, S.S. Feeding-habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Yeates, G.W.; Bongers, T. Nematode diversity in agroecosystems. Agr. Ecosyst. Environ. 1999, 74, 113–135. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Bongers, T.; Ferris, H. Nematode community structure as a bioindicator in environmental monitoring. Trends Ecol. Evol. 1999, 14, 224–228. [Google Scholar] [CrossRef]
- Ferris, H.; Matute, M.M. Structural and functional succession in the nematode fauna of a.soil food web. Appl. Soil Ecol. 2003, 23, 93–110. [Google Scholar] [CrossRef]
- Neher, D.A. Role of nematodes in soil health and their use as indicators. J. Nematol. 2001, 33, 161–168. [Google Scholar]
- Caixeta, L.B.; Pereira, T.J.; Castaneda, N.E.N.; Cares, J.E. Nematode communities as indicators of the status of a soil ecosystem influenced by mining practices in Brazil. Nematology 2016, 18, 265–276. [Google Scholar] [CrossRef]
- Da Silva, J.V.C.D.; Hirschfeld, M.N.C.; Cares, J.E.; Esteves, A.M. Land use, soil properties and climate variables influence the nematode communities in the Caatinga dry forest. Appl. Soil Ecol. 2020, 150, 103474. [Google Scholar] [CrossRef]
- Van Den, H.J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194. [Google Scholar] [CrossRef] [Green Version]
- Hu, N.; Li, H.; Tang, Z.; Li, Z.F.; Tian, J.; Lou, Y.L.; Li, J.W.; Li, G.C.; Hu, X.M. Community diversity, structure and carbon footprint of nematode food web following reforestation on degraded Karst soil. Sci. Rep. 2016, 6, 28138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.K.; Guan, P.T.; Wang, Y.L.; Li, Q.; Zhang, S.X.; Zhang, Z.Y.; Bezemer, T.M.; Liang, W.J. Community composition, diversity and metabolic footprints of soil nematodes in differently-aged temperate forests. Soil Biol. Biochem. 2015, 80, 118–126. [Google Scholar] [CrossRef]
- Shao, Y.H.; Wang, X.L.; Zhao, J.; Wu, J.P.; Zhang, W.X.; Neher, D.A.; Li, Y.X.; Lou, Y.P.; Fu, S.L. Subordinate plants sustain the complexity and stability of soil micro-food webs in natural bamboo forest ecosystems. J. Appl. Ecol. 2016, 53, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Liang, W.J.; Zhang, X.K. Soil nematode abundance and diversity in different forest types at changbai mountain, China. Zool. Stud. 2012, 51, 619–626. [Google Scholar]
- Cardoso, M.S.O.; Pedrosa, E.M.R.; Ferris, H.; Rolim, M.M.; Oliveira, L.S.C. Nematode fauna of tropical rainforest in brazil: A descriptive and seasonal approach. J. Nematol. 2016, 48, 116–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Deyn, G.B.; Raaijmakers, C.E.; van Ruijven, J.; Berendse, F.; van der Putten, W.H. Plant species identity and diversity effects on different trophic levels of nematodes in the soil food web. Oikos 2004, 106, 576–586. [Google Scholar] [CrossRef]
- Cesarz, S.; Ruess, L.; Jacob, M.; Jacob, A.; Schaefer, M.; Scheu, S. Tree species diversity versus tree species identity: Driving forces in structuring forest food webs as indicated by soil nematodes. Soil Biol. Biochem. 2013, 62, 36–45. [Google Scholar] [CrossRef]
- Cesarz, S.; Reich, P.B.; Scheu, S.; Ruess, L.; Schaefer, M.; Eisenhauer, N. Nematode functional guilds, not trophic groups, reflect shifts in soil food webs and processes in response to interacting global change factors. Pedobiologia 2015, 58, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Wardle, D.A.; Yeates, G.W. The dual importance of competition and predation as regulatory forces in terrestrial ecosystems—Evidence from decomposer food–webs. Oecologia 1993, 93, 303–306. [Google Scholar] [CrossRef]
- De Ruiter, P.C.; Neutel, A.M.; Moore, J.C. Energetics, patterns of interaction strengths, and stability in real ecosystems. Science 1995, 269, 1257–1260. [Google Scholar] [CrossRef]
- Holtkamp, R.; Kardol, P.; van der Wal, A.; Dekker, S.C.; van der Putten, W.H.; de Ruiter, P.C. Soil food web structure during ecosystem development after land abandonment. Appl. Soil Ecol. 2008, 39, 23–34. [Google Scholar] [CrossRef]
- Keith, A.M.; Brooker, R.W.; Osler, G.H.R.; Chapman, S.J.; Burslem, D.F.R.P.; van der Wal, R. Strong impacts of belowground tree inputs on soil nematode trophic composition. Soil Biol. Biochem. 2009, 41, 1060–1065. [Google Scholar] [CrossRef]
- Kang, D.; Deng, J.; Qin, X.W.; Hao, F.; Guo, S.J.; Han, X.H.; Yang, G.H. Effect of competition on spatial patterns of oak forests on the Chinese Loess Plateau. J. Arid Land 2017, 9, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Wang, K.B.; Chen, M.L.; Shangguan, Z.P.; Sweeney, S. Soil organic carbon storage capacity positively related to forest succession on the Loess Plateau, China. Catena 2013, 110, 1–7. [Google Scholar] [CrossRef]
- Jia, G.M.; Cao, J.; Wang, C.Y.; Wang, G. Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin, northwest China. Forest Ecol. Manag. 2005, 217, 117–125. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.B.; Xue, S.; Wang, G.L. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Zhong, Y.Q.W.; Yan, W.M.; Wang, R.W.; Wang, W.; Shangguan, Z.P. Decreased occurrence of carbon cycle functions in microbial communities along with long–term secondary succession. Soil Biol. Biochem. 2018, 123, 207–217. [Google Scholar] [CrossRef]
- Ma, R.T.; Hu, F.N.; Liu, J.F.; Wang, C.L.; Wang, Z.L.; Liu, G.; Zhao, S.W. Shifts in soil nutrient concentrations and C:N:P stoichiometry during long–term natural vegetation restoration. Peerj 2020, 8, e8382. [Google Scholar] [CrossRef]
- Dang, P.; Ngoc, H.V.; Shen, Z.; Liu, J.L.; Zhao, F.; Zhu, H.L.; Yu, X.; Zhao, Z. Changes in soil fungal communities and vegetation following afforestation with Pinus tabulaeformis on the Loess Plateau. Ecosphere 2018, 9, e24018. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.Y.; Liu, G.B.; Wang, H.S. The vegetation development in North Ziwulin forest region in last fifty years. Acta Bot. Boreal. Occident. Sin. 2002, 22, 1–8. [Google Scholar]
- Cheng, Y.B.; Xia, Y.D. Soil microbial and enzymatic activities across a chronosequence of chinese pine plantation development on the loess plateau of china. Pedosphere 2012, 22, 1–12. [Google Scholar]
- Zhong, Z.K.; Wang, X.; Zhang, X.Y.; Zhang, W.; Xu, Y.D.; Ren, C.J.; Han, X.H.; Yang, G.H. Edaphic factors but not plant characteristics mainly alter soil microbial properties along a restoration chronosequence of Pinus tabulaeformis stands on Mt. Ziwuling, China. For. Ecol. Manag. 2019, 453, 117625. [Google Scholar] [CrossRef]
- Liu, Y.L.; Zhu, G.Y.; Hai, X.Y.; Li, J.W.; Shangguan, Z.P.; Peng, C.H.; Deng, L. Long–term forest succession improves plant diversity and soil quality but not significantly increase soil microbial diversity: Evidence from the Loess Plateau. Ecol. Eng. 2020, 142, 105631. [Google Scholar] [CrossRef]
- Li, Y.Y.; Shao, M.A. Change of soil physical properties under long-term natural vegetation restoration in the Loess Plateau of China. J. Arid Environ. 2006, 64, 77–96. [Google Scholar] [CrossRef]
- Blake, G.R.; Hartge, K.H. Bulk Density. In Methods of Soil Analysis. Part 1, Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; American Society of Agronomy, Inc. and Soil Science Society of America: Madison, WI, USA, 1986; pp. 363–375. [Google Scholar]
- Jabro, J.D.; Stevens, W.B.; Iversen, W.M. Comparing two methods for measuring soil bulk density and moisture content. J. Soil Sci. 2020, 10, 233–243. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis. Part 2: Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 539–579. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen–total. In Methods of Soil Analysis. Part 2, Chemical and Microbial Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Agronomy Society of America, Agronomy Monograph 9: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Wu, J.; O’Donnell, A.G.; He, Z.L.; Syers, J.K. Fumigation–extraction method for the measurement of soil microbial biomass–S. Soil Biol. Biochem. 1994, 26, 117–125. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Liang, W.J.; Lou, Y.L.; Li, Q.; Zhong, S.; Zhang, X.K.; Wang, J.K. Nematode faunal response to long–term application of nitrogen fertilizer and organic manure in Northeast China. Soil Biol. Biochem. 2009, 41, 883–890. [Google Scholar] [CrossRef]
- Bongers, T. De Nematoden van Nederland. In Vormgeving en Technische Realisatie; Uitgeverij Pirola: Schoorl, The Netherlands, 1994; pp. 68–385. [Google Scholar]
- Ahmad, W.; Jairjpuri, M.S. Mononchida: The Predaceous Nematodes. In Nematology Monographs and Perspectives; Brill: Plantijnstraat, The Netherlands, 2010; Volume 7, pp. 1–299. [Google Scholar]
- Yeates, G.W.; Wardle, D.A. Nematodes as predators and prey: Relationships to biological control and soil processes. Pedobiologia 1996, 40, 43–50. [Google Scholar]
- Wagner, D.; Eisenhauer, N.; Cesarz, S. Plant species richness does not attenuate responses of soil microbial and nematode communities to a flood event. Soil Biol. Biochem. 2015, 89, 135–149. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; p. 144. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. J. Cardiothor. Vasc. An. 1972, 27, 261. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T.; de Goede, R. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- De Long, J.R.; Laudon, H.; Blume–Werry, G.; Kardol, P. Nematode community resistant to deep soil frost in boreal forest soils. Pedobiologia 2016, 59, 243–251. [Google Scholar] [CrossRef]
- Urzelai, A.; Hernandez, A.J.; Pastor, J. Biotic indices based on soil nematode communities for assessing soil quality in terrestrial ecosystems. Sci. Total Environ. 2000, 247, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Wasilewska, L. The effect of age of meadows on succession and diversity in soil nematode communities. Pedobiologia 1994, 38, 1–11. [Google Scholar]
- Yeates, G.W. Nematodes as soil indicators: Functional and biodiversity aspects. Biol. Fert. Soils 2003, 37, 199–210. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Hortal, S.; Armas, C.; Pugnaire, F.I. Interactions among soil, plants, and microorganisms drive secondary succession in a dry environment. Soil Biol. Biochem. 2014, 78, 298–306. [Google Scholar] [CrossRef]
- Li, Z.P.; Wei, Z.F.; Yang, X.D. Seasonal variations of soil nematode community at different secondary succession stages of evergreen broad–leaved forests in Ailao Mountain. Chinese Journal of Ecology. Chin. J. Ecol. 2016, 35, 3023–3031. [Google Scholar]
- Ye, Y.Y.; Rui, Y.C.; Zeng, Z.X.; He, X.Y.; Wang, K.L.; Zhao, J. Responses of soil nematode community to monoculture or mixed culture of a grass and a legume forage species in China. Pedosphere 2020, 30, 791–800. [Google Scholar] [CrossRef]
- Jourdan, M.; Piedallu, C.; Baudry, J.; Defossez, E.; Morin, X. Tree diversity and the temporal stability of mountain forest productivity: Testing the effect of species composition, through asynchrony and overyielding. Eur. J. Forest Res. 2020. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.B.; Shangguan, Z.P. Long–term natural succession improves nitrogen storage capacity of soil on the Loess Plateau, China. Soil Res 2014, 52, 262. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.B.; Tang, Z.S.; Shangguan, Z.P. Soil organic carbon dynamics following natural vegetation restoration: Evidence from stable carbon isotopes (delta C–13). Agr. Ecosyst. Environ. 2016, 221, 235–244. [Google Scholar] [CrossRef]
- Cortois, R.; Veen, G.F.C.; Duyts, H.; Abbas, M.; Strecker, T.; Kostenko, O.; Eisenhauer, N.; Scheu, S.; Gleixner, G.; de Deyn, G.B.; et al. Possible mechanisms underlying abundance and diversity responses of nematode communities to plant diversity. Ecosphere 2017, 8, e01719. [Google Scholar] [CrossRef]
- Deng, X.W.; Han, S.J.; Hu, Y.L.; Zhou, Y.M. Carbon and nitrogen transformations in surface soils under ermans birch and dark coniferous forests. Pedosphere 2009, 19, 230–237. [Google Scholar] [CrossRef]
- Priha, O.; Grayston, S.J.; Hiukka, R.; Pennanen, T.; Smolander, A. Microbial community structure and characteristicsof the organic matter in soils under Pinus sylvestris, Picea abies and Betula pendula at two forest sites. Biol. Fert. Soils 2001, 33, 17–24. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Shangguan, Z.P. The coupling interaction of soil water and organic carbon storage in the long vegetation restoration on the Loess Plateau. Ecol. Eng. 2016, 91, 574–581. [Google Scholar] [CrossRef]
- Asplund, J.; Kauserud, H.; Ohlson, M.; Nybakken, L. Spruce and beech as local determinants of forest fungal community structure in litter, humus and mineral soil. Fems Microbiol. Ecol. 2019, 95, fiy232. [Google Scholar] [CrossRef] [PubMed]
- Welti, E.; Helzer, C.; Joern, A. Impacts of plant diversity on arthropod communities and plant–herbivore network architecture. Ecosphere 2017, 8, e1983. [Google Scholar] [CrossRef] [Green Version]
- Salazar, D.; Jaramillo, A.; Marquis, R.J. The impact of plant chemical diversity on plant–herbivore interactions at the community level. Oecologia 2016, 181, 1199–1208. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T. Nematode indicators of organic enrichment. J. Nematol. 2006, 38, 3–12. [Google Scholar] [PubMed]
- Bongers, T.; Vandermeulen, H.; Korthals, G. Inverse relationship between the nematode maturity index and plant parasite index under enriched nutrient conditions. Appl. Soil Ecol. 1997, 6, 195–199. [Google Scholar] [CrossRef]
- Wang, K.B.; Chen, M.L.; Qin, J.; Liu, Y.; An, H.; Shangguan, Z.P. Plant species diversity and the relation with soil properties in natural succession process in Ziwuling area. Acta Bot. Boreal. Occident. Sin. 2007, 27, 2089–2096. [Google Scholar]
- Liang, J.; Wang, X.; Yu, Z.; Dong, Z.; Wang, J. Effects of vegetation succession on soil fertility within Farming-Plantation Ecotone in Ziwuling mountains of the Loess Plateau in China. Agr. Sci. China 2010, 9, 1481–1491. [Google Scholar] [CrossRef]
- Fu, B.J.; Liu, S.L.; Ma, K.M.; Zhu, Y.G. Relationships between soil characteristics, topography and plant diversity in a heterogeneous deciduous broad–leaved forest near Beijing, China. Plant Soil 2004, 261, 47–54. [Google Scholar] [CrossRef]
- Heydari, M.; Eslaminejad, P.; Kakhki, F.V.; Mirab–Balou, M.; Omidipour, R.; Prevosto, B.; Kooch, Y.; Lucas–Borja, M.E. Soil quality and mesofauna diversity relationship are modulated by woody species and seasonality in semiarid oak forest. Forest Ecol. Manag. 2020, 473, 118332. [Google Scholar] [CrossRef]
- Bargali, K.; Manral, V.; Padalia, K.; Bargali, S.S.; Upadhyay, V.P. Effect of vegetation type and season on microbial biomass carbon in Central Himalayan forest soils, India. Catena 2018, 171, 125–135. [Google Scholar] [CrossRef]
- Kitagami, Y.; Matsuda, Y. Temperature changes affect multi–trophic interactions among pines, mycorrhizal fungi, and soil nematodes in a microcosm experiment. Pedobiologia 2020, 78, 150595. [Google Scholar] [CrossRef]
- Wang, X.L.; Yu, S.Q.; Zhou, L.X.; Fu, S.L. Soil microbial characteristics and the influencing factors in subtropical forests. Acta Ecol. Sin. 2016, 36, 8–15. [Google Scholar] [CrossRef]
- Guan, P.T.; Zhang, X.K.; Yu, J.; Cheng, Y.Y.; Li, Q.; Andriuzzi, W.S.; Liang, W.J. Soil microbial food web channels associated with biological soil crusts in desertification restoration: The carbon flow from microbes to nematodes. Soil Biol. Biochem. 2018, 116, 82–90. [Google Scholar] [CrossRef]
- Thakur, M.P.; Tilman, D.; Purschke, O.; Ciobanu, M.; Cowles, J.; Isbell, F.; Wragg, P.D.; Eisenhauer, N. Climate warming promotes species diversity, but with greater taxonomic redundancy, in complex environments. Sci. Adv. 2017, 3, e1700866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, K.H.; Vogt, K.A.; Clark, H.J.; Schmitz, O.J.; Vogt, D.J. Biodiversity and the productivity and stability of ecosystems. Trends Ecol. Evol. 1996, 11, 372–377. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, Y.T.; Liu, S.R.; Fu, S.L.; Ming, A.G.; Li, X.Z.; Yao, M.J.; Li, H.; Tian, C. Mixture of tree species enhances stability of the soil bacterial community through phylogenetic diversity. Eur. J. Soil Sci. 2019, 70, 644–654. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; de Mazancourt, C.; Scherer–Lorenzen, M.; Bugmann, H. Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics. Ecol. Lett. 2014, 17, 1526–1535. [Google Scholar] [CrossRef]
- Geng, S.B.; Shi, P.L.; Song, M.H.; Zong, N.; Zu, J.X.; Zhu, W.R. Diversity of vegetation composition enhances ecosystem stability along elevational gradients in the Taihang Mountains, China. Ecol. Indic. 2019, 104, 594–603. [Google Scholar] [CrossRef]
- Haddad, N.M.; Crutsinger, G.M.; Gross, K.; Haarstad, J.; Tilman, D. Plant diversity and the stability of foodwebs. Ecol. Lett. 2011, 14, 42–46. [Google Scholar] [CrossRef]
- Borer, E.T.; Seabloom, E.W.; Tilman, D. Plant diversity controls arthropod biomass and temporal stability. Ecol. Lett. 2012, 15, 1457–1464. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Types | Forest Age | Location | Altitude (m) | Tree CW (m) | Tree DBH (cm) | Tree Height (m) | Canopy Density (%) | H’plant | Main Plant Species |
|---|---|---|---|---|---|---|---|---|---|
| PT | 26y | 36°04′05″ N 108°32′03″ E | 1377 | 1.63 ± 0.25c | 27.30 ± 0.80b | 6.56 ± 0.15ab | 60.33 ± 3.18c | 0.59 ± 0.07c | Pinus tabuliformis Carrière, Acer ginnala Maxim., Lonicera japonica Thunb. |
| BP | 27y | 36°04′22″ N 108°31′41″ E | 1341 | 2.95 ± 0.20b | 30.79 ± 6.08b | 5.97 ± 1.37b | 74.67 ± 2.73b | 1.43 ± 0.05a | Betula platyphylla Sukaczev, Larix gmelinii Kuzen., Acer ginnala, Lonicera japonica |
| QL | 30y | 36°02′57″ N 108°32′13″ E | 1435 | 3.88 ± 0.74a | 64.83 ± 17.51a | 8.46 ± 0.98a | 85.33 ± 3.18a | 1.02 ± 0.05b | Quercus liaotungensis Koidz., Rosa hugonis Hemsl., Spiraea salicifolia Lour., Carex lanceolata Boott |
| Soil Properties | Season | PT | BP | QL |
|---|---|---|---|---|
| SBD (g/cm3) | April | 1.07 ± 0.07a | 1.02 ± 0.07a | 1.14 ± 0.09a |
| August | 1.09 ± 0.08A | 1.15 ± 0.09A | 1.07 ± 0.14A | |
| p value | 0.97 | 0.661 | 0.732 | |
| SMC (%) | April | 7.05 ± 0.35b | 14.93 ± 1.55a | 16.29 ± 2.41a |
| August | 16.54 ± 0.85B | 30.34 ± 2.02A | 25.06 ± 1.70A | |
| p value | 0.001 | 0.004 | 0.041 | |
| pH | April | 7.60 ± 0.01a | 7.64 ± 0.04a | 7.65 ± 0.03a |
| August | 7.71 ± 0.02A | 7.70 ± 0.01A | 7.72 ± 0.02A | |
| p value | 0.055 | 0.204 | 0.127 | |
| SOC (g/kg) | April | 18.23 ± 2.52b | 28.99 ± 1.88a | 22.39 ± 2.70ab |
| August | 13.35 ± 1.00B | 30.13 ± 2.52A | 22.19 ± 2.54AB | |
| p value | 0.146 | 0.737 | 0.960 | |
| TN (g/kg) | April | 1.29 ± 0.16b | 2.35 ± 0.20a | 1.85 ± 0.25ab |
| August | 1.21 ± 0.08B | 2.64 ± 0.20A | 1.81 ± 0.19B | |
| p value | 0.661 | 0.354 | 0.920 | |
| MBC (mg/kg) | April | 627.08 ± 23.65b | 1010.90 ± 49.91a | 902.87 ± 69.99a |
| August | 1178.89 ± 154.73A | 1385.54 ± 120.17A | 1218.70 ± 153.36A | |
| p value | 0.024 | 0.045 | 0.032 | |
| MBN (mg/kg) | April | 95.23 ± 0.98c | 164.48 ± 3.44a | 145.18 ± 4.82b |
| August | 129.54 ± 18.24A | 125.39 ± 15.29A | 108.89 ± 1.86A | |
| p value | 0.134 | 0.067 | 0.052 | |
| DOC (mg/kg) | April | 107.05 ± 2.19b | 163.55 ± 4.49a | 145.09 ± 8.21a |
| August | 199.34 ± 9.06B | 286.77 ± 6.86A | 273.09 ± 5.09A | |
| p value | 0.001 | 0.001 | 0.001 | |
| DON (mg/kg) | April | 20.25 ± 0.76c | 32.29 ± 0.67a | 26.84 ± 0.27b |
| August | 34.90 ± 5.44A | 41.74 ± 1.96A | 37.43 ± 4.38A | |
| p value | 0.056 | 0.010 | 0.073 | |
| NO3−–N (mg/kg) | April | 0.91 ± 0.48a | 1.49 ± 0.50a | 0.83 ± 0.11a |
| August | 1.41 ± 0.10B | 5.67 ± 1.04A | 2.60 ± 0.18B | |
| p value | 0.365 | 0.022 | 0.001 | |
| NH4+–N (mg/kg) | April | 2.07 ± 0.42a | 4.10 ± 0.40a | 3.43 ± 1.01a |
| August | 5.62 ± 0.44B | 6.06 ± 1.00B | 9.00 ± 2.33A | |
| p value | 0.004 | 0.144 | 0.006 |
| Genus | Family | Trophic Group | c–p Value | April | August | ||||
|---|---|---|---|---|---|---|---|---|---|
| PT | BP | QL | PT | BP | QL | ||||
| Acrobeles | Cephalobidae | BF | 2 | 4.11 II | 0.76 I | 2.50 II | 11.52 II | 7.75 II | 34.70 II |
| Acrobeloides | Cephalobidae | BF | 2 | – | 6.94 II | 2.64 II | 7.19 II | 4.75 I | 1.99 I |
| Cephalobus | Cephalobidae | BF | 2 | 6.56 II | 10.13 II | 10.98 II | 3.74 I | 20.52 II | 13.34 II |
| Cervidellus | Cephalobidae | BF | 2 | 5.02 II | 5.92 II | 2.92 II | 10.64 II | 7.39 II | 15.67 II |
| Chiloplacus | Cephalobidae | BF | 2 | – | 3.70 II | 1.81 II | 5.36 II | 5.93 II | – |
| Eucephalobus | Cephalobidae | BF | 2 | 3.52 II | 0.76 I | 2.91 II | 7.39 II | 10.13 II | 35.42 II |
| Panagrolaimus | Panagrolaimidae | BF | 1 | – | – | 0.42 I | – | 1.47 I | – |
| Placodira | Cephalobidae | BF | 2 | – | 4.06 II | – | – | – | – |
| Rhabditis | Rhabditidae | BF | 1 | 0.91 I | 10.49 II | 4.73 II | – | 2.65 I | 4.32 II |
| Protorhabditis | Rhabditidae | BF | 1 | – | – | – | 1.31 I | 1.19 I | – |
| Alaimus | Alaimidae | BF | 4 | 0.91 I | 7.11 II | 0.97 I | – | 1.19 I | – |
| Plectus | Plectidae | BF | 2 | – | 10.38 II | – | 2.94 I | 56.70 III | 0.66 I |
| Wilsonema | Plectidae | BF | 2 | 1.82 I | 3.30 I | 1.81 II | – | 5.38 II | – |
| Bacterivores | 22.86 ± 6.28b | 63.56 ± 19.11a | 31.68 ± 2.93b | 50.09 ± 9.38B | 125.04 ± 49.14A | 106.11 ± 36.47AB | |||
| Aphelenchoides | Aphelenchoididae | FF | 2 | 28.58 III | 22.74 II | 3.06 II | 3.04 I | 21.72 II | 7.99 II |
| Aphelenchus | Aphelenchidae | FF | 2 | 4.39 II | 17.30 II | 16.11 III | 5.67 II | 3.84 I | 5.05 II |
| Filenchus | Tylenchidae | FF | 2 | 0.87 I | 5.27 II | 0.69 I | 4.15 I | 6.84 II | 3.66 I |
| Nothotylenchus | Anguinidae | FF | 2 | – | – | 0.56 I | – | – | – |
| Diphtherophora | Diphtherophoridae | FF | 3 | – | 1.47 I | – | – | 4.12 I | – |
| Tylencholaimus | Leptonchidae | FF | 4 | 68.55 III | 78.54 III | 18.31 III | 241.96 III | 124.28 III | 102.81 III |
| Fungivores | 102.40 ± 31.39ab | 125.32 ± 44.24a | 38.73 ± 13.68b | 254.81 ± 22.46A | 160.80 ± 13.29B | 119.51 ± 17.03C | |||
| Criconemoides | Criconematidae | PP | 3 | 3.64 II | 12.42 II | 3.89 II | – | – | 0.66 I |
| Ditylenchus | Anguinidae | PP | 2 | 1.66 I | 3.55 II | 1.95 II | – | 6.50 II | 20.78 II |
| Helicotylenchus | Hoplolaimidae | PP | 3 | 18.18 II | 24.58 II | 7.23 II | 6.08 II | 42.64 II | 4.18 II |
| Malenchus | Tylenchidae | PP | 2 | 5.15 II | 2.48 I | 7.37 II | 6.88 II | 20.52 II | 1.99 I |
| Neopsilenchus | Tylenchidae | PP | 2 | 0.87 I | 6.74 II | 20.05 III | 8.21 II | 6.77 II | 39.22 III |
| Pararotylenchus | Hoplolaimidae | PP | 3 | – | 3.50 II | – | – | – | – |
| Paratylenchus | Tylenchulidae | PP | 2 | – | 14.67 II | – | – | 17.4 II | – |
| Pratylenchus | Pratylenchidae | PP | 3 | – | – | 2.77 II | – | 2.10 I | 1.39 I |
| Psilenchus | Tylenchidae | PP | 2 | 0.87 I | – | – | – | – | – |
| Rotylenchus | Hoplolaimidae | PP | 3 | 4.90 II | 3.05 I | 5.96 II | – | – | – |
| Tylenchorhynchus | Dolichodoridae | PP | 3 | 1.58 I | 15.13 II | 4.85 II | – | 25.15 II | – |
| Tylenchus | Tylenchidae | PP | 2 | 3.28 II | 0.76 I | 1.11 I | 5.98 II | 16.97 II | 9.32 II |
| Xiphinema | Longidoridae | PP | 5 | 62.52 III | 4.51 II | 1.95 II | 42.02 II | 23.26 II | 6.24 II |
| Plant parasites | 102.65 ± 24.35a | 91.41 ± 38.38ab | 57.13 ± 20.48b | 69.16 ± 20.03B | 161.31 ± 51.87A | 83.78 ± 37.06AB | |||
| Aporcelaimellus | Aporcelaimidae | OP | 5 | 1.70 I | 19.22 II | 7.49 II | 14.87 II | 18.44 II | 23.85 II |
| Dorydorella | Qudsianematidae | OP | 4 | 3.49 II | – | 0.42 I | – | – | – |
| Epidorylaimus | Qudsianematidae | OP | 4 | – | 1.02 I | – | – | – | – |
| Eudorylaimus | Qudsianematidae | OP | 4 | 2.62 I | 2.93 I | 5.26 II | 5.93 II | 0.66 I | |
| Kochinema | Qudsianematidae | OP | 4 | – | – | 0.56 I | 5.67 II | – | 7.31 II |
| Microdorylaimus | Qudsianematidae | OP | 4 | 0.87 I | 4.06 II | 4.31 II | 14.37 II | 4.19 I | 4.00 II |
| Paraxonchium | Paraxonchidae | OP | 5 | – | 0.76 I | 0.42 I | 1.72 I | – | 0.66 I |
| Prodorylaimus | Thornenematidae | OP | 5 | – | – | – | – | 6.29 II | – |
| Thonus | Qudsianematidae | OP | 4 | 9.33 II | 23.00 II | 1.80 II | 2.63 I | 2.37 I | 20.86 II |
| Discolaimium | Discolaimidae | OP | 5 | 12.36 II | 4.97 II | 4.03 II | 19.65 II | 1.19 I | 9.78 II |
| Discolaimus | Discolaimidae | OP | 5 | 0.87 I | – | 1.67 II | 1.21 I | 2.10 I | – |
| Nygolaimus | Nygolaimidae | OP | 5 | 10.91 II | 4.46 II | 6.11 II | 1.72 I | 10.40 II | 7.63 II |
| Paravulvus | Nygolaimidae | OP | 5 | – | – | 0.69 I | 1.31 I | 5.59 II | 0.66 I |
| Prionchulus | Mononchidae | OP | 4 | – | – | – | – | 2.10 I | – |
| Trischistoma | Tripylidae | OP | 3 | – | – | – | 1.72 I | – | 1.39 I |
| Omnivore-predators | 42.15 ± 9.10ab | 60.41 ± 14.76a | 27.49 ± 7.15b | 70.15 ± 30.18A | 58.58 ± 18.14A | 76.81 ± 16.81A | |||
| Total | 270.06 ± 11.30ab | 340.69 ± 65.03a | 155.02 ± 13.13b | 444.21 ± 45.60AB | 505.73 ± 79.63A | 386.21 ± 95.66B | |||
| Genus number | 29 | 35 | 35 | 28 | 36 | 29 | |||
| Nematode Characteristics | SMC | SOC | DOC | MBC | TN | DON | MBN | NH4+–N | NO3−–N |
|---|---|---|---|---|---|---|---|---|---|
| TNA | 0.529 * | 0.179 | 0.596 ** | 0.688 ** | 0.258 | 0.623 ** | −0.084 | 0.634 ** | 0.391 |
| PP | 0.273 | 0.237 | 0.276 | 0.252 | 0.333 | 0.224 | −0.277 | 0.404 | 0.125 |
| BF | 0.659 ** | 0.457 | 0.668 ** | 0.729 ** | 0.476* | 0.623 ** | 0.143 | 0.803 ** | 0.367 |
| FF | 0.208 | −0.196 | 0.309 | 0.461 | −0.126 | 0.439 | −0.004 | 0.227 | 0.247 |
| OP | 0.304 | 0.071 | 0.354 | 0.347 | 0.053 | 0.336 | −0.101 | 0.250 | 0.357 |
| H’ | 0.374 | 0.807 ** | 0.119 | −0.067 | 0.748 ** | 0.251 | 0.181 | 0.314 | −0.008 |
| J’ | 0.248 | 0.634 ** | 0.063 | −0.073 | 0.564 * | 0.055 | 0.044 | 0.226 | 0.049 |
| λ | −0.261 | −0.655 ** | −0.049 | 0.132 | −0.598 ** | −0.106 | 0.013 | −0.254 | 0.021 |
| MI | 0.120 | −0.209 | 0.196 | 0.231 | −0.233 | 0.232 | 0.078 | −0.017 | 0.328 |
| PPI | −0.563 * | −0.619 ** | −0.449 | −0.254 | −0.578 * | −0.463 | −0.222 | −0.307 | −0.365 |
| PPI/MI | −0.483 * | −0.234 | −0.472 * | −0.443 | −0.187 | −0.548 * | −0.284 | −0.194 | −0.494 * |
| WI | 0.090 | −0.021 | 0.196 | 0.199 | −0.045 | 0.203 | 0.320 | 0.066 | 0.216 |
| NCR | 0.318 | 0.442 | 0.260 | 0.232 | 0.409 | 0.170 | 0.241 | 0.352 | 0.044 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, N.; Zhao, S.; Huang, J.; Geng, D.; Wang, N.; Yang, P. Seasonal Stabilities of Soil Nematode Communities and Their Relationships with Environmental Factors in Different Temperate Forest Types on the Chinese Loess Plateau. Forests 2021, 12, 246. https://doi.org/10.3390/f12020246
Huo N, Zhao S, Huang J, Geng D, Wang N, Yang P. Seasonal Stabilities of Soil Nematode Communities and Their Relationships with Environmental Factors in Different Temperate Forest Types on the Chinese Loess Plateau. Forests. 2021; 12(2):246. https://doi.org/10.3390/f12020246
Chicago/Turabian StyleHuo, Na, Shiwei Zhao, Jinghua Huang, Dezhou Geng, Nan Wang, and Panpan Yang. 2021. "Seasonal Stabilities of Soil Nematode Communities and Their Relationships with Environmental Factors in Different Temperate Forest Types on the Chinese Loess Plateau" Forests 12, no. 2: 246. https://doi.org/10.3390/f12020246