1. Introduction
Bark beetles interrupt two important transport systems in the trees they attack. First, the gallery construction by adult beetles and their feeding larvae can cause significant damage to the tree by the partial girdling of its phloem, reducing or preventing the movement of photosynthate. Second, phytopathogenic fungi carried by bark beetles, most notably those collectively called blue-stain fungi (Fam. Ophiostomatacea) that grow into the xylem tracheids to access water and nutrients, block water transport from roots to leaves [
1]. Either the phloem girdling caused by bark beetles or the xylem blockage by fungal penetration can potentially kill a tree. However, the speed in which these take effect on the tree can differ and may show which organism is the leading driver of mortality. Hubbard and others [
2] found that lodgepole pine (
Pinus contorta Douglas) attacked by the mountain pine beetle (
Dendroctonus ponderosae Hopk.) exhibited a faster decrease in leaf water potential and transpiration than mechanically girdled trees and in fact, girdled trees survived for two years following treatment. This finding implicates that fungal blockage of the xylem is likely the leading driver of mortality in
Pinus species attacked by bark beetles associated with phytopathogenic blue-stain fungi. Although the physiological response of dying
Pinus species colonized by bark beetle/fungal systems has been compared to the maximum damage that can be caused by bark beetles alone (girdling) [
2], this has not been studied in other genera within the family Pinaceae. Increasing our knowledge about these responses in other genera of Pinaceae moves us closer to understand bark beetle/fungi/host interactions in the largest conifer family. While
Pinus experience highly synchronized bark beetle attacks, these occur less synchronously in
Abies,
Larix, Picea, and
Pseudotsuga species [
3]; thus, studying other genera in this family offers information about how defense responses are adapted to the different attack patterns. Within Pinaceae,
Pseudotsuga is a good candidate in which to contrast the findings in
Pinus, since both genera are attacked by
Dendroctonus bark beetles that are univoltine (single generation per year) and that are associated with several blue-stain fungi [
3].
In North America, the Douglas-fir beetle (
D. pseudotsugae Hopk.) is the main bark beetle killing Douglas-fir (
Pseudotsuga menziesii (Mirbel) Franco). Like other bark beetles that become irruptive, the Douglas-fir beetle is an important driver of tree mortality, nutrient cycling, and habitat creation in Douglas-fir-dominated forests. At endemic population levels, their attack is limited to trees previously weakened by factors that include drought stress, windfall, defoliation, and fire scorching [
4,
5]. The ecological impacts of their irruptive populations are like those of other bark beetles such as reducing canopy cover, accelerating successional patterns, and increasing streamflow [
6,
7,
8,
9]. However, irruptive populations of Douglas-fir beetle have historically never reached the size of species like the mountain pine beetle or the spruce beetle (
D. rufipennis Kirby).
The Douglas-fir beetle uses a semiochemical communication system in which attraction pheromones [
10] enable it to perform attacks in numbers large enough to overcome the defenses of healthy trees, while an anti-aggregation pheromone, 3-methylcyclohex-2-en-1-one (MCH) [
11], is released when the number of beetle attacks saturates a tree [
12]. Manipulations of the populations of this bark beetle have been used to create biological legacies that benefit the ecosystem, among other uses [
13] with a push–pull technique in which attracting and anti-aggregation pheromones are used simultaneously to displace beetle activity away from undesired areas and into areas of interest [
14]. These manipulations indicate that the use of this technique could work for tree research and may offer results similar to manual thinning in management operations.
Our study broadly aimed to further our understanding of the physiological process of transpiration decline in response to bark beetle (girdling) and associated fungi colonization (xylem blockage) of Pinaceae. We also hoped to demonstrate that Douglas-fir beetles could be lured to attack selected trees in a small area where trees are near each other (less than 60 m). Specifically, our primary objective was to compare Douglas-fir physiological responses to fungal infection (inoculated by beetles) with manual phloem girdling to quantify which is the primary factor causing mortality. Based on previous work in lodgepole pine [
2], we hypothesized that beetle-attacked trees would exhibit decline first and die faster than mechanically girdled trees due to the effects of their associated fungi. Elucidating the response of Douglas-fir to simulated beetle girdling or blue-stain fungal colonization will increase understanding of their role in mortality and broaden our overall knowledge about these processes across some of the most broadly occurring conifer genera in the western United States.
2. Materials and Methods
2.1. Study Site and Treatments Establishement
Our study site was located in the Arapahoe/Roosevelt National Forest near the town of Red Feather Lakes in northern Colorado, at an altitude of 2600 m above sea level. Vegetation was of a mix of Douglas-fir, lodgepole pine, ponderosa pine (
P. ponderosa Douglas ex C. Lawson), and scattered aspen (
Populus tremuloides Michx.) with an undergrowth dominated by dwarf juniper (
Juniperus communis L.) and
Vaccinium spp. The average temperature is 4 °C ranging between −12 °C and 28 °C. Mean annual precipitation is 38 cm, most of which occurs as snowfall between October and April [
15].
On 21 May 2018, we selected 30 Douglas-fir trees inside a 60 m diameter plot. Trees were over 20 cm in diameter at breast height (dbh), of similar size (mean = 24 cm, s = 3.0 cm), and did not show bark beetle activity. We randomly assigned three treatment groups of ten trees each: Control, Girdled, and Beetle-Attacked and installed sap flow sensors on all (details below). On 6 June 2018, we attached Douglas-fir beetle attracting pheromone packets (seudenol, frontalin, ethanol, and a Douglas-fir kairomone, Synergy Semiochemicals Corp., BC, Canada) to the Beetle-Attacked trees subgroup to entice the attack by host-searching beetles. Immediately after the beetle attack was initiated (9 to 14 June 2018), we used a single side razor and a hatchet to mechanically remove a 30 cm tall strip of bark and phloem from around the entire circumference 30 cm above the sap flow sensors of the Girdled treatment group. Control trees were left untreated. To protect the Control and Girdled treatment trees, two days after beetle attacks began, we affixed one Methylcyclohexanone bubble (MCH, Synergy Semiochemicals Corp., Delta, BC, Canada) to each tree. In addition, to prevent spillover beyond our plot, we attached additional MCH bubbles to a series of >30.5 cm diameter Douglas-fir trees SE, E, and NE of our plot as the wind blew mostly towards those directions, and treatment trees were set at the western side of the plot.
2.2. Data Collection
We quantified total daily transpiration with homemade Granier style sapflow sensors installed on the north side of each tree (dbh) [
16]. Each probe was covered with insulation (Reflectix, Inc., Markleville, IN, USA) to reduce heating from solar radiation. Sap velocity (
v) was measured every 60 seconds and 10 minute averages were stored on a data logger (CR1000, Campbell Scientific, Logan, UT, USA). Data were collected during the first growing season between 27 May 2018 and 4 November 2018 (23 weeks) and not collected during winter. Data collection resumed on 7 March 2019 until 22 October 2019 (24 weeks). Sap velocity was estimated according to the equation by Granier [
16] as follows:
Δ
TM and Δ
T are the temperature differences between the two probes when
v = 0 and
v > 0, respectively. We estimated sapwood area (
ASW) using a linear regression between radial sapwood thickness (obtained with an increment borer) and dbh, measured from random trees in the area fitting our selection parameters. We estimated total transpiration (
Ec) as the product of sap velocity and sapwood area:
We measured pre-dawn leaf water potential weekly from three randomly selected trees per treatment group. We took measurements for 15 weeks during the 2018 growing season (11 July 2018–24 October 2018) and 18 weeks during the 2019 season (17 May 2019–20 September 2019). Because individual Douglas-fir needles were too small to fit in our pressure chamber, we used short branchlets. Using a pole pruner, we cut one branch from the mid-canopy (>4 m) of each tree. From those, we cut three small branchlets and trimmed them of phloem to fit into the pressure chamber. We measured water potential using a PMS 600 Pressure Chamber (PMS Instrument Comp., Albany, OR, USA). We rotated collections on sampled trees to limit defoliation throughout the growing season. On 14 and 23–29 July 2018, a faulty power supply compromised the sap flux equipment affecting the measurements of seven trees (one Control, three Girdled, and three Beetle-Attacked) for about two weeks; thus, we omitted this data. We removed one Girdled tree from the analysis which died unexpectedly for reasons unrelated to the girdling treatment. In addition, we removed one Beetle-Killed tree that was attacked later than the others and died at the end of the 2019 sampling period.
In winter, a year after Douglas-fir beetle attacks, we quantified their attack densities from two- one dm
2 bark samples taken at dbh from the north and south sides of the trees. Attack density was used to determine if all trees experienced similar attack rates. We cut and removed samples with a chisel and hammer and examined them under a MZ16 dissecting microscope (Leica Microsystems Inc., Buffalo Grove, IL, USA). We measured three attributes to determine attack density. First, we measured the number of holes made by the Douglas-fir beetles in the outer bark, after shaving off the outer cork layer (phellem) using a 25.4 cm drawknife. Secondly, we measured phloem penetrating holes made by attacking parents located at the initial (lower extreme) of the parental galleries. Third, we measured the number of complete or partial vertical galleries present at the phloem and cambium tissue layers. We also noted the presence of resin (pitch) pockets [
17], defined as a resin accumulation between the phloem and xylem formed in response to biotic agents that indicates a successful tree defense.
2.3. Statistical Analyses
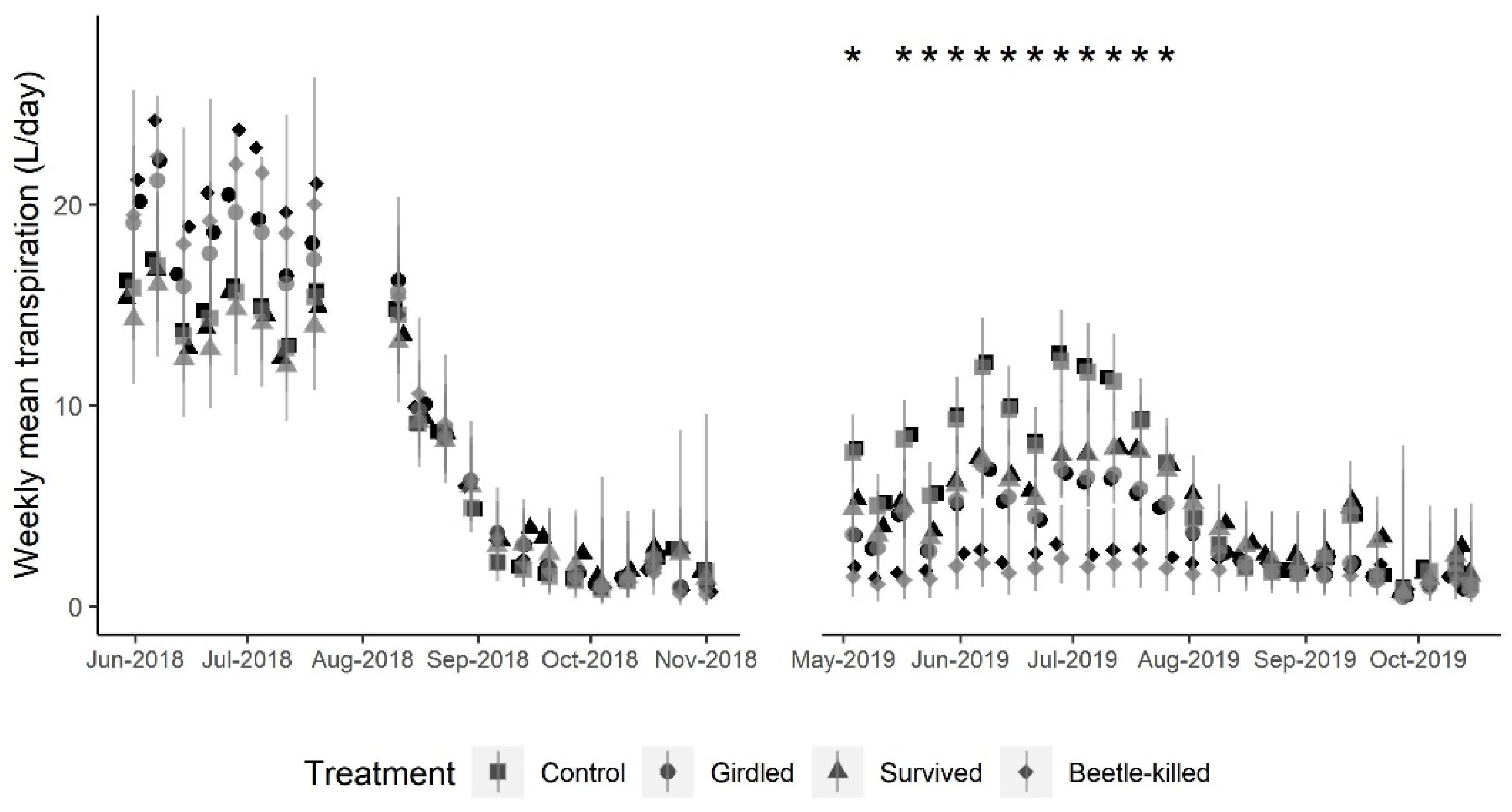
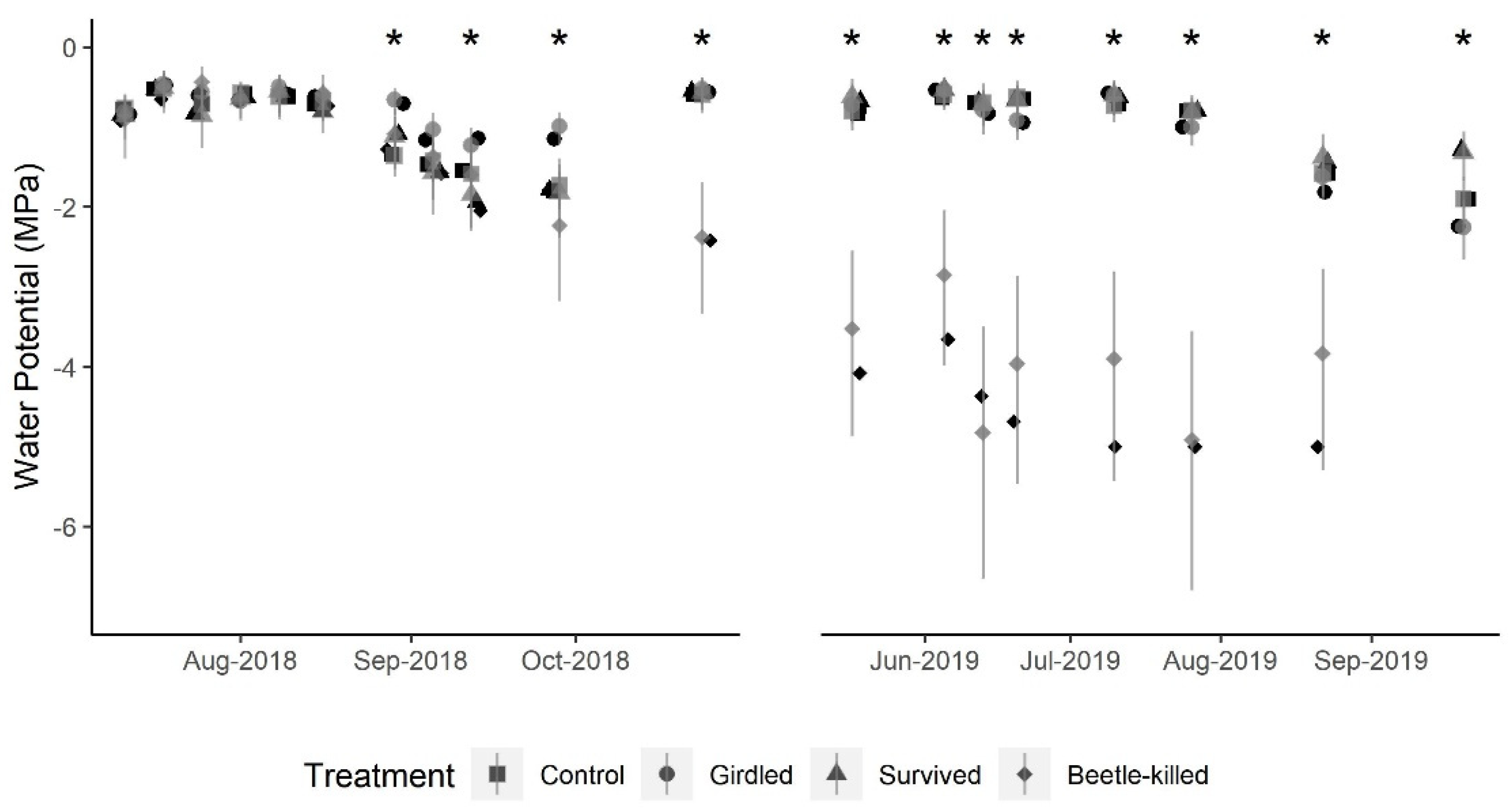
The experimental unit for all analyses was the individual tree. We estimated total daily transpiration for each tree using the sap flux data according to the procedure outlined above (Sap Flux/Transpiration) and averaged values across weeks. We used a Gaussian generalized linear mixed model (GLMM) with a log link mean function that included the fixed effects of the week, treatment, and their interaction to test whether weekly mean transpiration per day differed between treatments across the course of the study period (excluding winter months) (n = 47 weeks; 29 May 2018–15 October 2019). A random effect for individual trees was included to account for repeated measures. We tested for all pairwise comparisons between treatment means for each week and adjusted
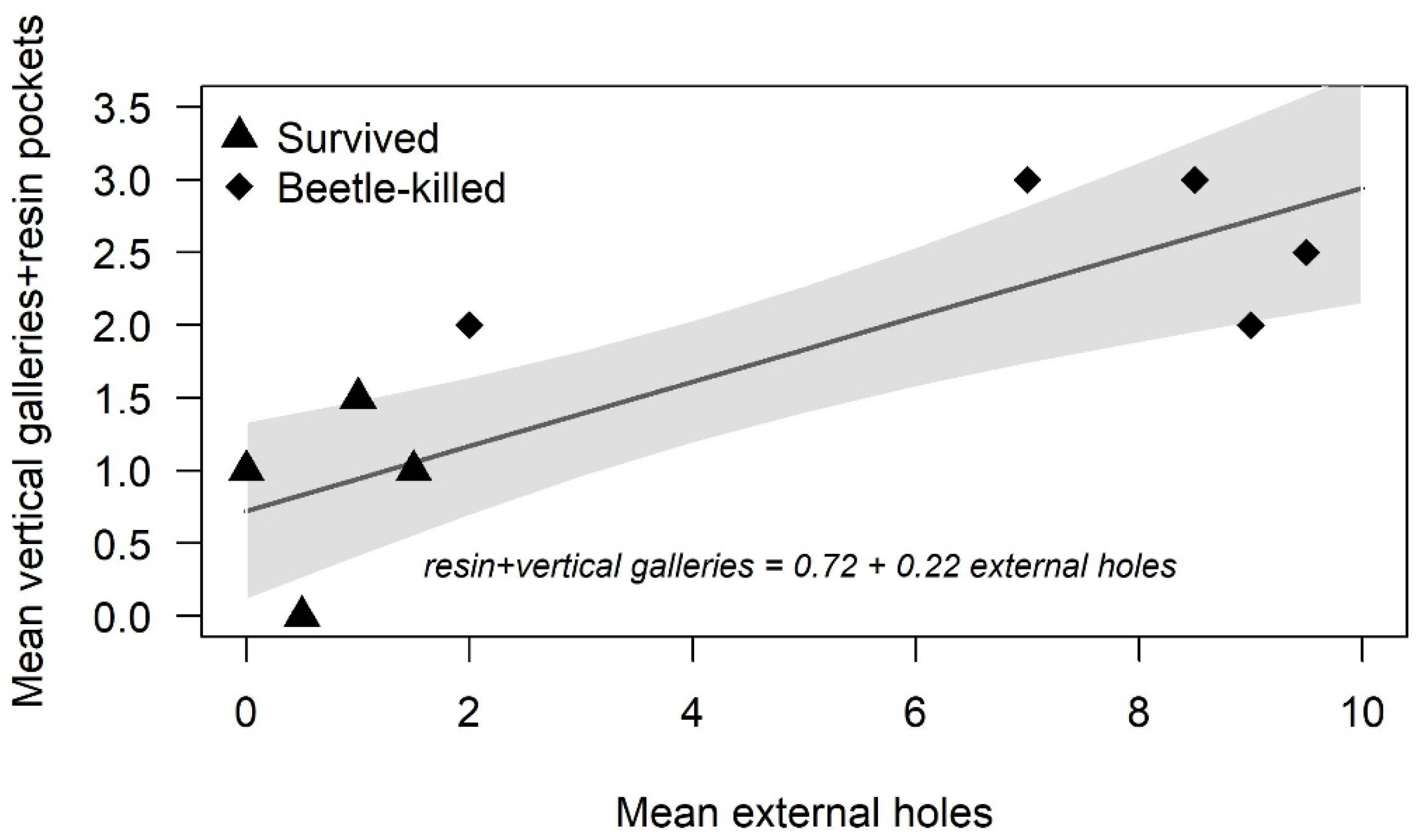
p-values to account for multiple comparisons using the Tukey method. The same procedure was used to analyze water potential by week; however, to fit an appropriate statistical model that was bounded above by zero, water potential was multiplied by −1 and the model was fit on the transformed data while results were presented on the back-transformed response. Since measurement of external holes is much less labor-intensive than measurements of vertical galleries or phloem penetrating holes, we used two simple linear regressions of mean vertical galleries and mean entry holes on mean external holes to determine whether external holes could be used as a proxy for either of the two other measurements. All analyses were performed using R version 4.1.1 (R Core Team, 2021, Vienna, Austria) with the packages
glmmTMB [
18],
lme4 [
19], and
emmeans [
20].
4. Discussion
This study supports the assertion that blue-stain fungi are the key driver of mortality in Douglas-fir attacked by the Douglas-fir beetle. However, the physiological response took significantly longer than the approximately 21 days documented in lodgepole pines attacked by the mountain pine beetle [
2]. Although leaf water potential in Beetle-Killed trees differed significantly from Girdled or Control trees during late-October precipitation events, their death was not confirmed with the transpiration data until the 2019 season. Thus, it took trees nearly an entire year to succumb to the beetle’s fungal colonization. While at the beginning of the 2019 growing season, we observed external signs of mortality that were typical of Douglas-fir successfully attacked by Douglas-fir beetle, these signs later became somewhat atypical [
5]. That is, while early in the season the typical, red-colored needles were present in the bottom-most limbs of Beetle-Killed trees early in the season, the discolored (yellow) needles above the red needles rapidly senesced without ever turning to typical red.
We can think of three possible factors to explain why some beetle-attacked Douglas-fir Survived: (1) low attack density, (2) a diminished or lack of phytopathogenic fungi vectored by the beetles, or (3) variations in tree resistance among attacked trees. First, beetle attack density may not have exceeded a required minimal beetle attack threshold. We confirmed that while our estimated attacked density for the Beetle-Killed trees matched reported lethal densities for this insect in Douglas-fir in the study region, they were on the lower end of reported lethal densities, and in the case of trees in the Survived treatment, their attack densities (defined by brood galleries) did not even reach lethal levels [
5]. Boone and others [
22] found that the attack success of mountain pine beetle on lodgepole pine is highly dependent on beetle population density. This was evident when all attacked trees in an experiment died during the peak of an epidemic of that species [
2].
Secondly, we forcibly recruited Douglas-fir beetles from potentially collapsing populations with attractants. That is, we understand that Douglas-fir beetle populations can increase following disturbance events such as wildfire [
23,
24] and that their activity around wildfire edges usually lasts three to four years after wildfire and then collapses [
4]. Since the High Park fire had burned in 2012 close to our plot, it is likely that the beetle attacking our trees were part of a five-year post-wildfire population. This collapse may also be influenced by, or influence characteristics of, the associated fungi carried by the beetles. As seen in mountain pine beetle eruptive populations, a collapse in their populations can occur even when suitable hosts are available due to a reduction in the frequency of their mutualistic blue-stain fungi [
25]. However, we do not know if Douglas-fir beetle associated fungi (
O. pseudotsugae) (Rumb.) von Arx change in frequency in collapsing beetle populations (as in mountain pine beetle) and only suspect that Douglas-fir beetle has a similar dependence on its blue-stain associate [
26]. Understanding the interaction between the Douglas-fir beetle and its associated fungi is further complicated because
Ophiostoma species. and other carrier
Dendroctonus beetles have shown interactions that range from mutualistic to antagonistic [
27,
28]. A low frequency or lack of phytopathogenic blue-stain fungi may also help explain the observed similarities in estimated transpiration between the Survived and Girdled groups during the 2019 season. Hypothetically, by removing fungal effects, the damage done by beetles would only have partially girdled the Survived trees when attempting to build their brood galleries. This disruption of photosynthate transport would signal the tree to reduce photosynthesis, causing stomata to close and reducing transpiration and observed xylem sap flux [
29].
Lastly, Survived and Beetle-Killed trees might have had different defensive capabilities at the time of the attack. Survived trees were able to reject beetle attacks right after the initial attack and blue-stain fungal infections, as indicated by resin pocket formation, and were able to stop beetle reproduction as indicated by parental galleries lacking larval galleries. Resin pockets or pitch pockets occurring beneath the site of a Douglas-fir beetle initial attacks have been examined and attributed before to this beetle [
30]. Resin pockets are formed in other conifers attacked by
Dendroctonus bark beetles (summarized in [
30]). These pockets indicate an effective defense response of the tree being attacked [
31,
32]. The lower number of vertical galleries found on Survived trees reflects that colonizing beetles were being deterred by the tree defenses. Moreover, the presence of resin pockets just outside the cambium in Survived trees indicated that attacking beetles penetrating the phloem were being stopped relatively quickly in these trees. Resin or pitch pockets have been described in Douglas-fir trees that survived Douglas-fir beetle attacks of similar densities (0.54/dm
2) as those found in this study (0.60/dm
2) [
30]. Resin pockets can be much shorter than full-length brood galleries made by the Douglas-fir beetle and other
Dendroctonus species that make elongated galleries. Adding the count of resin pockets to the count of parental galleries still resulted in a lower attack density, which would result in an underestimation of parental gallery density when using these metrics as a gallery density proxy. Consequently, it is possible that attack density was similar in the two groups of trees, and our observed mortality rate was a sign of the different defensive capacities in our experimental trees. This is sustained by earlier reports of mass attacks made by Douglas-fir beetle to healthy standing trees that resulted in even lower (i.e., 22.4%, 42.3%) mortality rates than ours [
21]. The use of external bark beetle holes for estimating gallery density as a proxy of attack density would be a helpful alternative to removing bark and counting galleries. Therefore, it would be useful to develop a calibration for vertical galleries predicted by external holes.
In conclusion, we found that blue-stain fungi were the primary cause of mortality in Douglas-fir beetle attacked Douglas-fir trees and that we were able to lure the beetles to successfully attack individual trees of our choosing, which may be useful for some forest management applications. Until now, we have generalized that conifers attacked by bark beetles and their fungal associates were killed by fungi based on studies of Pinus species and associated bark beetles. Differences in the defenses (mainly constitutive vs induced) of these two tree genera suggested the need to reevaluate this assumption in more genera of Pinaceae. Here we addressed this lack of knowledge by showing that while fungi are the main driver of mortality in different Pinaceae genera, those with fast induced defenses may not die as quickly as those relying mainly on their constitutive defenses. We believe our contribution will serve as a baseline to understand the range of interactions between Pinaceae conifer genera and their enemies and to contribute to the understanding of mechanisms of tree defense in this large family.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}