
The Significance of Aggregation Methods in Functional Group Modeling


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Model Formulation
2.2. Forest System
2.3. Aggregation and the Creation of Plant Functional Types
- (1)
- One PFT—All 32 tree species amalgamated into one functional group.
- (2)
- Evergreenness —Lump all 32 tree species into two groups according to the attributes of evergreen or deciduous leaves.
- (3)
- Tolerance—Lump the 32 tree species into five shade-tolerance groups (see Appendix A for details on how tolerance was defined and applied to these species).
- (4)
- Genus—Lump the 32 tree species by genus for the 20 genera represented by the 32 species.
3. Results and Discussion
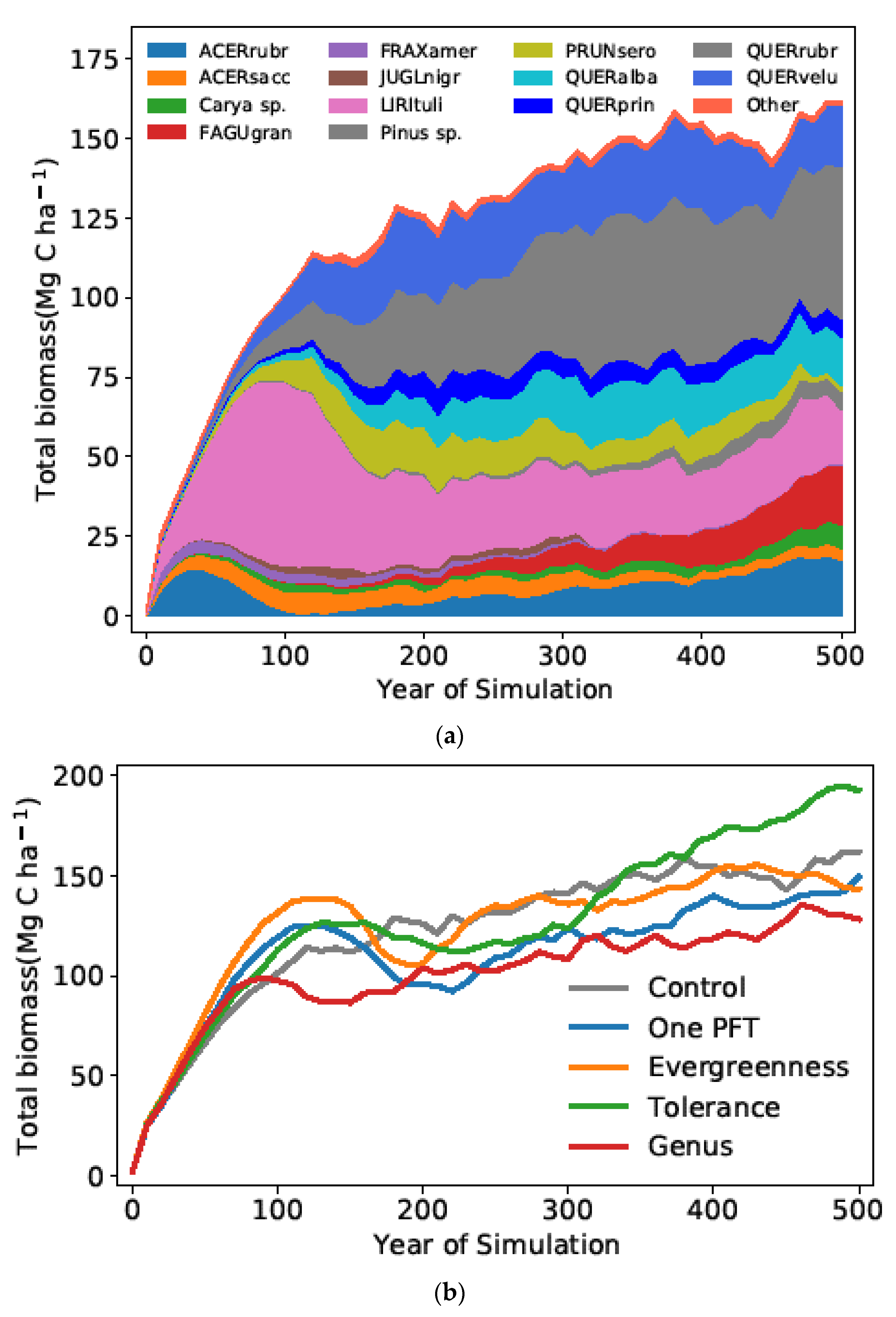
3.1. Inter-Protocol Comparisons: The Effect of Lumping Protocol
3.2. Intra-Protocol Comparisons: The Effect of Weighting within a Functional Group
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Shade Tolerance Responses
References
- Smith, T.M.; Shugart, H.H.; Woodward, F.I. (Eds.) Plant Functional Types: Their Relevance to Ecosystem Properties and Global Change; Cambridge University Press: Cambridge, UK, 1997; 369p. [Google Scholar]
- Hooper, D.U.; Chapin, F.S., III; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Richardson, L.F. Weather Prediction by Numerical Processes; Cambridge University Press: Cambridge, UK, 1922. [Google Scholar]
- Deardorff, J.W. Efficient prediction of ground surface temperature and moisture, with inclusion of a layer of vegetation. J. Geophys. Res. Space Phys. 1978, 83, 1889–1903. [Google Scholar] [CrossRef] [Green Version]
- Anten, N.P.; Hirose, T. Limitations on photosynthesis of competing individuals in stands and the consequences for canopy structure. Oecologia 2001, 129, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Dale, M.B. Systems analysis and ecology. Ecology 1970, 51, 2–16. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Shugart, H.H. FAREAST: A forest gap model to simulate dynamics and patterns of eastern Eurasian forests. J. Biogeogr. 2005, 32, 1641–1658. [Google Scholar]
- Foster, A.C.; Shuman, J.K.; Shugart, H.H.; Dwire, K.A.; Fornwalt, P.; Sibold, J.; Negron, J. Validation and application of a forest gap model to the southern Rocky Mountains. Ecol. Model. 2017, 351, 109–128. [Google Scholar] [CrossRef]
- Shugart, H.H.; Foster, A.; Wang, B.; Druckenbrod, D.; Ma, J.; Lerdau, M.; Saatchi, S.; Yang, X.; Yan, X. Gap models across micro- to mega-scales of time and space: Examples of Tansley’s ecosystem concept. For. Ecosyst. 2020, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Shugart, H.H.; Shuman, J.K.; Lerdau, M.T. Forests and ozone: Productivity, carbon storage and feedbacks. Sci. Rep. 2016, 6, 22133. [Google Scholar] [CrossRef] [Green Version]
- Shuman, J.K.; Foster, A.C.; Shugart, H.H.; Hoffman-Hall, A.; Krylov, A.; Loboda, T.; Ershov, D.; Sochilova, E. Fire disturbance and climate change: Implications for Russian forests. Environ. Res. Lett. 2017, 12, 035003. [Google Scholar] [CrossRef]
- Foster, A.C.; Shuman, J.K.; Shugart, H.H.; Negron, J. Modeling the interactive effects of spruce beetle infestation and climate on subalpine vegetation. Ecosphere 2018, 9, e02437. [Google Scholar] [CrossRef]
- Wang, B.; Shuman, J.; Shugart, H.H.; Lerdau, M.T. Biodiversity matters in feedbacks between climate change and air quality: A study using an individual-based model. Ecol. Appl. 2018, 28, 1223–1231. [Google Scholar] [CrossRef] [Green Version]
- Shugart, H.H.; West, D.C. Development of an Appalachian deciduous forest succession model and its application to assessment of the impact of the chestnut blight. J. Environ. Manag. 1977, 5, 161–170. [Google Scholar]
- Wang, B.; Shugart, H.H.; Lerdau, M.T. An individual-based model of forest volatile organic compound emissions—UVAFME-VOCnv1.0. Ecol. Model. 2017, 350, 69–78. [Google Scholar] [CrossRef]
- Noble, I.R.; Slatyer, R.O. The use of vital attributes to predict successional changes in plant communities subject to recurrent disturbances. Vegetatio 1980, 43, 5–21. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.; Vose, J.M.; Volin, J.C.; GreshamWilliam, C.; Bowman, W.D. Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: A test across biomes and functional groups. Oecologia 1998, 114, 471–482. [Google Scholar] [CrossRef]
- Shugart, H.H.; Saatchi, S.; Hall, F.G. Importance of structure and its measurement in quantifying function of forest ecosystems. J. Geophys. Res. Biogeochem. 2010, 115, G00E13. [Google Scholar] [CrossRef]
- Druckenbrod, D.L.; Martin-Benito, D.; Orwig, D.A.; Pederson, N.; Poulter, B.; Renwick, K.M.; Shugart, H.H. Redefining temperate forest responses to climate and disturbance in the eastern United States: New insights at the mesoscale. Glob. Ecol. Biogeogr. 2019, 28, 557–575. [Google Scholar] [CrossRef]
- Quegan, S.; Le Toan, T.; Chave, J.; Dall, J.; Exbrayat, J.-F.; Minh, D.H.T.; Lomas, M.; D’Alessandro, M.M.; Paillou, P.; Papathanassiou, K.; et al. The European Space Agency BIOMASS mission: Measuring forest above-ground biomass from space. Remote Sens. Environ. 2019, 227, 44–60. [Google Scholar] [CrossRef] [Green Version]
- Le Toan, T.; Quegan, S.; Davidson, M.; Balzter, H.; Paillou, P.; Papathanassiou, K.; Plummer, S.; Rocca, F.; Saatchi, S.; Shugart, H.; et al. The BIOMASS mission: Mapping global forest biomass to better understand the terrestrial carbon cycle. Remote Sens. Environ. 2011, 115, 2850–2860. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Baker, F.S. A revised tolerance table. J. For. 1949, 47, 179–181. [Google Scholar]
- Mooney, H.A. The Carbon Balance of Plants. Annu. Rev. Ecol. Syst. 1972, 3, 315–346. [Google Scholar] [CrossRef]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, 4th ed.; Springer: Berlin, Germany, 2003. [Google Scholar]
- Horn, H.S. The Adaptive Geometry of Trees; Princeton Press: Princeton, NJ, USA, 1971. [Google Scholar]
- Kramer, P.J.; Kozlowski, T.T. Physiology of Trees; McGraw-Hill Book, Co.: New York, NY, USA, 1960. [Google Scholar]
- Urban, D.; Bonan, G.; Smith, T.; Shugart, H. Spatial applications of gap models. For. Ecol. Manag. 1991, 42, 95–110. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Shugart, H.H.; Wang, B.; Lerdau, M. The Significance of Aggregation Methods in Functional Group Modeling. Forests 2021, 12, 1560. https://doi.org/10.3390/f12111560
Zhang H, Shugart HH, Wang B, Lerdau M. The Significance of Aggregation Methods in Functional Group Modeling. Forests. 2021; 12(11):1560. https://doi.org/10.3390/f12111560
Chicago/Turabian StyleZhang, Huan, Herman H. Shugart, Bin Wang, and Manuel Lerdau. 2021. "The Significance of Aggregation Methods in Functional Group Modeling" Forests 12, no. 11: 1560. https://doi.org/10.3390/f12111560