
Wood Density and Ring Width in Quercus rotundifolia Trees in Southern Portugal


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sites and Sampling
2.3. X-Ray Microdensitometry
2.4. Data Analysis
3. Results and Discussions
3.1. Within-Ring Structure
3.2. Ring Width and Wood Density
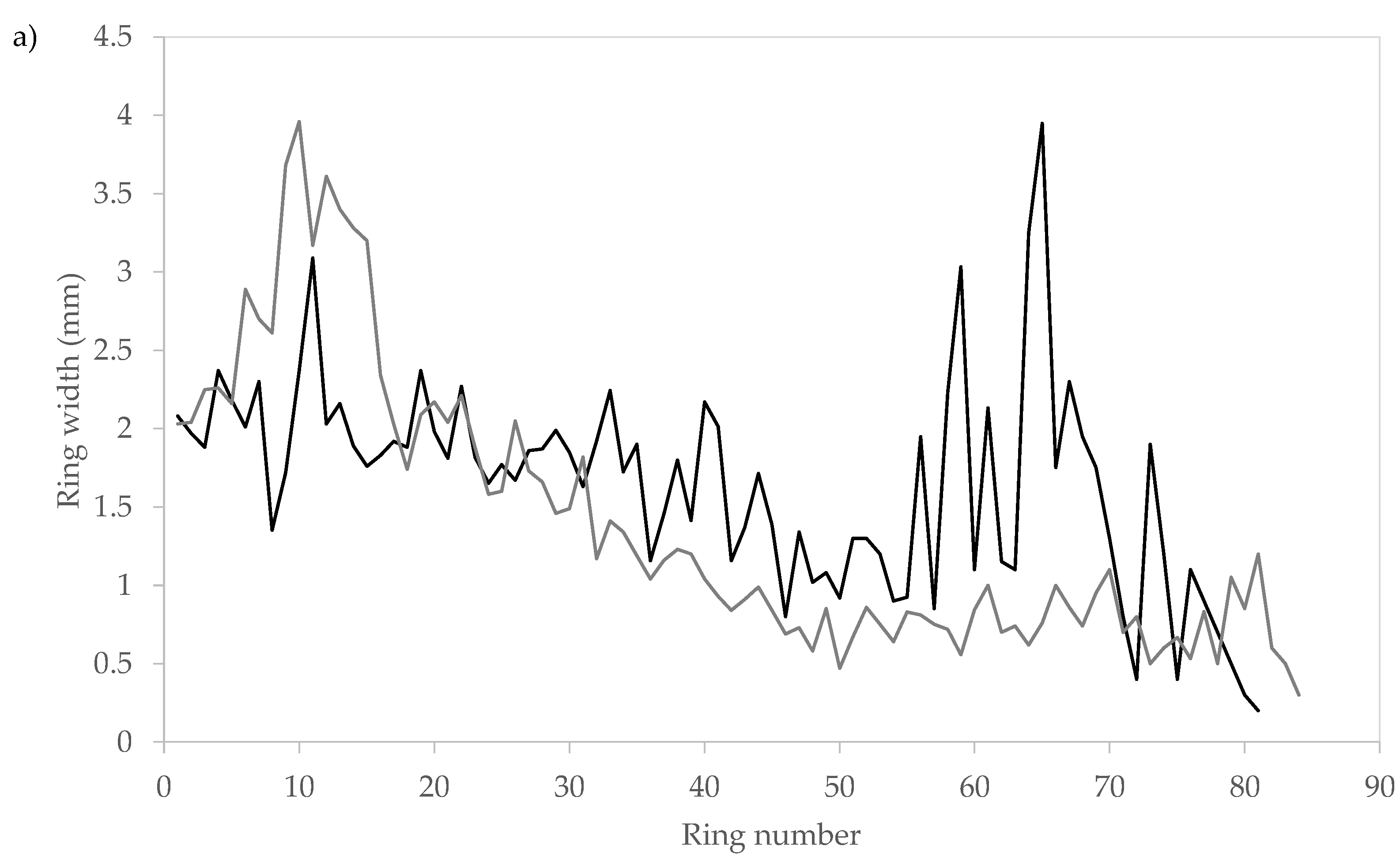
3.3. Radial Variation of Ring Width
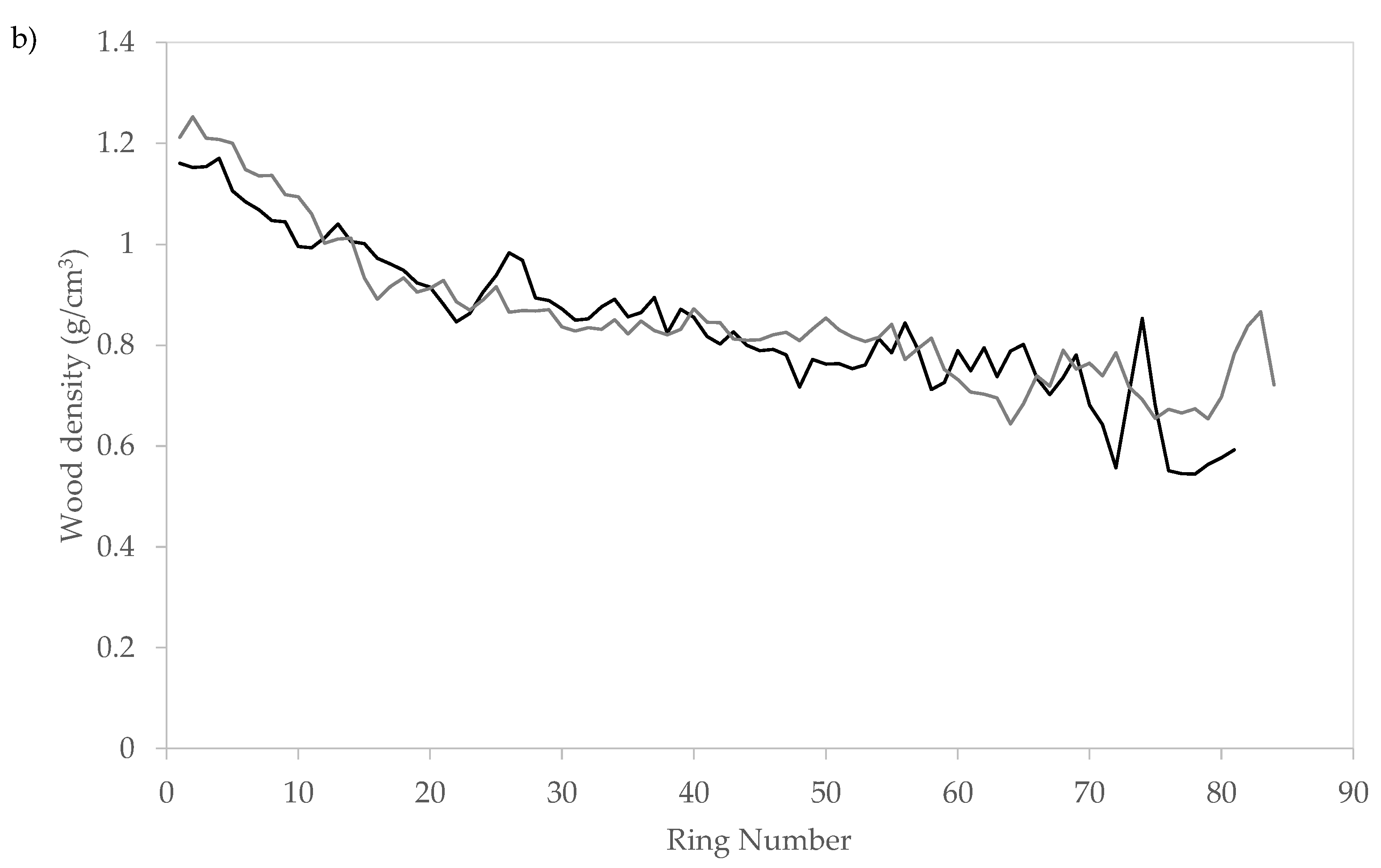
3.4. Radial Variation of Wood Density
3.5. Wood Density and Ring Width Relationships
3.6. Ring Width and Wood Density Variability Sources
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Rigo, D.; Caudullo, G. Quercus ilex in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Durrant, T.H., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- FAO. L’état des ressources génétiques forestières. In Rapport National. Le Royaume Du Maroc; Haut Commissariat aux Eaux et Forêts et à la Lutte contre la Désertification. Projet TCP/RAB/3303-BABY 2 M; FAO: Rome, Italy, 2014; Available online: https://www.fao.org/3/i3825e/i3825e45.pdf (accessed on 26 October 2021).
- ICNF. 6º Inventário Florestal Nacional. Relatório Final; Instituto da Conservação da Natureza e das Florestas: Lisbon, Portugal, 2015. [Google Scholar]
- Soto, A.; Lorenzo, Z.; Gil, L. Differences in fine-scale genetic structure and dispersal in Quercus ilex L. and Q. suber L.: Consequences for regeneration of mediterranean open woods. Heredity 2007, 99, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Capelo, J.; Catry, F. Biologia, ecologia e distribuição da azinheira (Biology, ecology and distribution of holm oak). Os montados-Muito para além das árvores 2007, 3, 119–129. [Google Scholar]
- Pelegrín, E.; Peguero-Pina, J.J.; Sancho-Knapik, D. Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Lousã, M.; Fabião, A. A azinheira: Quercus ilex ou Quercus rotundifolia? In Proceedings of the Sobreiro e Cortiça/Cork oak and Cork, European Conference on Cork oak and Cork, Lisboa, Portugal, 5–7 May 1997; Pereira, H., Ed.; Centro de Estudos Florestais: Lisboa, Portugal, 1998; pp. 248–251. [Google Scholar]
- Fady, B.; Médail, F. Temperate and mediterranean Forests | Mediterranean Forest Ecosystems. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Oxford, UK, 2004; pp. 1403–1414. ISBN 978-0-12-145160-8. [Google Scholar]
- López-Tirado, J.; Vessella, F.; Schirone, B.; Hidalgo, P.J. Trends in evergreen oak suitability from assembled species distribution models: Assessing climate change in south-western Europe. New For. 2018, 49, 471–487. [Google Scholar] [CrossRef]
- Natalini, F.; Alejano, R.; Vázquez-Piqué, J.; Cañellas, I.; Gea-Izquierdo, G. The role of climate change in the widespread mortality of holm oak in open woodlands of Southwestern Spain. Dendrochronologia 2016, 38, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Mayoral, C.; Pardos, M.; González, M.S.; Brendel, O.; Pita, P. Ecological implications of different water use strategies in three coexisting mediterranean tree species. For. Ecol. Manag. 2016, 382, 76–87. [Google Scholar] [CrossRef]
- Mayor, X.; Rodà, F. Growth response of holm oak (Quercus ilex L) to commercial thinning in the Montseny mountains (NE Spain). Ann. des Sci. For. 1993, 50, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Montserrat-Martí, G.; Camarero, J.J.; Palacio, S.; Pérez-Rontomé, C.; Milla, R.; Albuixech, J.; Maestro, M. Summer-drought constrains the phenology and growth of two coexisting Mediterranean oaks with contrasting leaf habit: Implications for their persistence and reproduction. Trees 2009, 23, 787–799. [Google Scholar] [CrossRef] [Green Version]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Effects of a severe drought on Quercus ilex radial growth and xylem anatomy. Trees 2003, 18, 83–92. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Ma, S.; Rambal, S.; Misson, L.; Ourcival, J.-M.; Limousin, J.-M.; Pereira, J.; Papale, D. On the differential advantages of evergreenness and deciduousness in mediterranean oak woodlands: A flux perspective. Ecol. Appl. 2010, 20, 1583–1597. [Google Scholar] [CrossRef]
- Silva, S.; Costa, E.M.; Borges, A.; Carvalho, A.P.; Monteiro, M.J.; Pintado, M.M.E. Nutritional characterization of acorn flour (a traditional component of the Mediterranean gastronomical folklore). J. Food Meas. Charact. 2016, 10, 584–588. [Google Scholar] [CrossRef]
- Carvalho, A. Madeiras Portuguesas—Estrutura Anatómica, Propriedades, Utilizações; Direcção-Geral das Florestas: Lisbon, Portugal, 1997. [Google Scholar]
- Barboutis, J.A.; Philippou, J.L. Evergreen Mediterranean hardwoods as particleboard raw material. Build. Environ. 2007, 42, 1183–1187. [Google Scholar] [CrossRef]
- Dilem, A. Etude de Quelques Propriétés Du Bois de Chêne Vert (Quercus ilex) Dans La Région d’El-Hassasna (Saida-Algérie). For. Méditerr. 1995, XVI, 74–78. [Google Scholar]
- Zobel, B.J.; Van Buijtenen, J. Wood Variation. Its Causes and Control; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Woodcock, D.; Shier, A. Wood specific gravity and its radial variations: The many ways to make a tree. Trees 2002, 16, 437–443. [Google Scholar] [CrossRef]
- Bastin, J.-F.; Fayolle, A.; Tarelkin, Y.; Bulcke, J.V.D.; De Haulleville, T.; Mortier, F.; Beeckman, H.; Van Acker, J.; Serckx, A.; Bogaert, J.; et al. Wood Specific Gravity Variations and Biomass of Central African Tree Species: The Simple Choice of the Outer Wood. PLoS ONE 2015, 10, e0142146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.-Y.; Owoundi, R.E.; Nepveu, G.; Mothe, F.; Dhôte, J.-F. Modelling wood density in European oak (Quercus petraea and Quercus robur) and simulating the silvicultural influence. Can. J. For. Res. 1993, 23, 2587–2593. [Google Scholar] [CrossRef]
- Bergès, L.; Dupouey, J.-L.; Franc, A. Long-term changes in wood density and radial growth of Quercus petraea Liebl. In northern France since the middle of the nineteenth century. Trees 2000, 14, 398–408. [Google Scholar] [CrossRef]
- Sousa, V.B.; Louzada, J.-L.; Pereira, H. Age trends and within-site effects in wood density and radial growth in Quercus faginea mature trees. For. Syst. 2016, 25, 8. [Google Scholar] [CrossRef] [Green Version]
- Knapic, S.; Louzada, J.L.; Leal, S.; Pereira, H. Within-tree and between-tree variation of wood density components in cork oak trees in two sites in Portugal. Forestry 2008, 81, 465–473. [Google Scholar] [CrossRef]
- Polge, H.; Keller, R. Qualité du bois et largeur d’accroissements en forêt Tronçais. Ann. Sci. Forest 1973, 30, 91–125. [Google Scholar] [CrossRef]
- Ackermann, F. Influence du type de station forestière sur les composantes intracernes de la densité du bois du chêne pédonculé (Quercus robur L.) dans les chênaies de l’Adour et des coteaux basco-béarnais. Ann. For. Sci. 1995, 52, 635–652. [Google Scholar] [CrossRef] [Green Version]
- Bergès, L.; Nepveu, G.; Franc, A. Effects of ecological factors on radial growth and wood density components of sessile oak (Quercus petraea Liebl.) in Northern France. For. Ecol. Manag. 2008, 255, 567–579. [Google Scholar] [CrossRef]
- Sousa, V.B.; Louzada, J.L.; Pereira, H. Variation of Ring Width and Wood Density in Two Unmanaged Stands of the Mediterranean Oak Quercus faginea. Forests 2018, 9, 44. [Google Scholar] [CrossRef] [Green Version]
- Lebourgeois, F. Les Chênes Sessile et Pédonculé (Quercus petraea Liebl. et Quercus robur L.) Dans Le Réseau RENECOFOR: Rythme de Croissance Radiale, Anatomie Du Bois, de l’aubier et de l’écorce. Rev. For. Française 1999, 51, 522–536. [Google Scholar] [CrossRef] [Green Version]
- Guilley, E.; Hervé, J.-C.; Nepveu, G. The influence of site quality, silviculture and region on wood density mixed model in Quercus petraea Liebl. For. Ecol. Manag. 2004, 189, 111–121. [Google Scholar] [CrossRef]
- Zhang, S.; Romane, F. Variations de la croissance radiale de Quercus ilex L en fonction du climat. Ann. For. Sci. 1991, 48, 225–234. [Google Scholar] [CrossRef]
- Gutiérrez, E.; Campelo, F.; Camarero, J.J.; Ribas, M.; Muntán, E.; Nabais, C.; Freitas, H. Climate controls act at different scales on the seasonal pattern of Quercus ilex L. stem radial increments in NE Spain. Trees 2011, 25, 637–646. [Google Scholar] [CrossRef]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; Saurer, M.; De Micco, V. Contrasting physiological responses to Mediterranean climate variability are revealed by intra-annual density fluctuations in tree rings of Quercus ilex L. and Pinus pinea L. Tree Physiol. 2018, 38, 1213–1224. [Google Scholar] [CrossRef]
- Rozenberg, P.; Franc, A.; Cahalan, C. Incorporating Wood Density in Breeding Programs for Softwoods in Europe: A Strategy and Associated Methods. Silvae Genet. 2001, 50, 1–7. [Google Scholar]
- Wheeler, E.; Baas, P.; Gasson, P. IAWA List of Microscopic Features for Hardwood Identification. IAWA J. Int. Assoc. Wood Anat. 1989, 10, 219–332. [Google Scholar]
- Cherubini, P.; Gartner, B.L.; Tognetti, R.; Bräker, O.U.; Schoch, W.; Innes, J. Identification, measurement and interpretation of tree rings in woody species from Mediterranean climates. Biol. Rev. 2003, 78, 119–148. [Google Scholar] [CrossRef] [Green Version]
- Schweingruber, F.H. Anatomy of European Woods; Swiss Federal Institute for Forest, Snow and Landscape Research: Haupt, Germany, 1990. [Google Scholar]
- Sousa, V.B.; Leal, S.; Quilhó, T.; Pereira, H. Characterization of Cork Oak (Quercus suber) Wood Anatomy. IAWA J. 2009, 30, 149–161. [Google Scholar] [CrossRef]
- Sousa, V.B.; Cardoso, S.; Pereira, H. Age trends in the wood anatomy of Quercus faginea. IAWA J. 2014, 35, 293–306. [Google Scholar] [CrossRef]
- Woodcock, D.W. Climate sensitivity of wood-anatomical features in a ring-porous oak (Quercus macrocarpa). Can. J. For. Res. 1989, 19, 639–644. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Castro-Díez, P.; Pérez-Rontomé, C.; Montserrat-Martí, G. Stem xylem features in three Quercus (Fagaceae) species along a climatic gradient in NE Spain. Trees 1997, 12, 90–96. [Google Scholar] [CrossRef]
- Campelo, F.; Nabais, C.; Gutiérrez, E.; Freitas, H.; García-González, I. Vessel features of Quercus ilex L. growing under Mediterranean climate have a better climatic signal than tree-ring width. Trees 2010, 24, 463–470. [Google Scholar] [CrossRef]
- Tardif, J.C.; Conciatori, F. Influence of climate on tree rings and vessel features in red oak and white oak growing near their northern distribution limit, southwestern Quebec, Canada. Can. J. For. Res. 2006, 36, 2317–2330. [Google Scholar] [CrossRef] [Green Version]
- Matisons, R.; Brumelis, G. Influence of climate on tree-ring and earlywood vessel formation in Quercus robur in Latvia. Trees 2012, 26, 1251–1266. [Google Scholar] [CrossRef]
- Gupta, P.; Gupta, S. Wood anatomy of Indian oaks, with reference to systematic, ecological and evolutionary perspectives. Nord. J. Bot. 2020, 38. [Google Scholar] [CrossRef]
- Crivellaro, A.; Schweingruber, F.H. Atlas of Wood, Bark and Pith Anatomy of Eastern Mediterranean Trees and Shrubs; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-642-37234-6. [Google Scholar]
- Tavares, F.; Louzada, J.L.; Pereira, H. Variation in wood density and ring width in Acacia melanoxylon at four sites in Portugal. Eur. J. For. Res. 2013, 133, 31–39. [Google Scholar] [CrossRef]
- Pirralho, M.; Flores, D.; Sousa, V.B.; Quilhó, T.; Knapic, S.; Pereira, H. Evaluation on paper making potential of nine Eucalyptus species based on wood anatomical features. Ind. Crop. Prod. 2014, 54, 327–334. [Google Scholar] [CrossRef]
- Wheeler, E.A.; Baas, P. Wood Identification -A Review. IAWA J. 1998, 19, 241–264. [Google Scholar] [CrossRef]
- Camarero, J.J. Linking functional traits and climate-growth relationships in Mediterranean species through wood density. IAWA J. 2019, 40, 215-S2. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Sangüesa-Barreda, G.; Vergarechea, M. Prior height, growth, and wood anatomy differently predispose to drought-induced dieback in two Mediterranean oak species. Ann. For. Sci. 2015, 73, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, S.; Guo, J.; Ren, H.; Wang, Y.; Zhang, Y.; Yin, Y. Characterization and comparison of the wood anatomical traits of plantation grown Quercus acutissima and Quercus variabilis. IAWA J. 2021, 42, 244–257. [Google Scholar] [CrossRef]
- Voulgaridis, E.; Passialis, C. Valorisation de Différentes Espèces Méditerranéennes—Characteristics and Technological Properties of the Wood of Mediterranean Evergreen Hardwoods. Forêt. Méllitermnéenne 1995, XVI, 3–12. [Google Scholar]
- Knapic, S.; Louzada, J.L.; Leal, S.; Pereira, H. Radial variation of wood density components and ring width in cork oak trees. Ann. For. Sci. 2007, 64, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Genet, A.; Auty, D.; Achim, A.; Bernier, M.; Pothier, D.; Cogliastro, A. Consequences of faster growth for wood density in northern red oak (Quercus rubra Liebl.). Forestry 2012, 86, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Tsoumis, G. Science and Technology of Wood: Structure, Properties, Utilization; Van Nostrand Reinhold: New York, NY, USA, 1991. [Google Scholar]
- King, D.A.; Davies, S.J.; Tan, S.; Noor, N.S.M. The role of wood density and stem support costs in the growth and mortality of tropical trees. J. Ecol. 2006, 94, 670–680. [Google Scholar] [CrossRef]
- Chao, K.-J.; Phillips, O.; Gloor, E.; Monteagudo, A.; Torres-Lezama, A.; Martínez, R.V. Growth and wood density predict tree mortality in Amazon forests. J. Ecol. 2008, 96, 281–292. [Google Scholar] [CrossRef]
- Lopez-Iglesias, B.; Villar, R.; Poorter, L. Functional traits predict drought performance and distribution of Mediterranean woody species. Acta Oecologica 2014, 56, 10–18. [Google Scholar] [CrossRef]
- Lei, H.; Milota, M.R.; Gartner, B.L. Between- and Within-Tree Variation in the Anatomy and Specific Gravity of Wood in Oregon White oak (Quercus Garryana Dougl.). IAWA J. 1996, 17, 445–461. [Google Scholar] [CrossRef]
- Sousa, V.B.; Cardoso, S.; Quilhó, T.; Pereira, H. Growth rate and ring width variability of teak, Tectona grandis (Verbenaceae) in an unmanaged forest in East Timor. Rev. Biol. Trop. 2012, 60, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Paul, B. The Application of Silviculture in Controlling the Specific Gravity of Wood; Technical Bulletin No.1288; USDA Forest Service: Washington, DC, USA, 1963.
- Cartan-Son, M.; Floret, C.; Galan, M.J.; Grandjanny, M.; Le Floc’h, E.; Maistre, M.; Perret, P.; Romane, F. Factors affecting radial growth of Quercus ilex L. in a coppice stand in Southern France. Vegetatio 1992, 99–100, 61–68. [Google Scholar] [CrossRef]
- Mayor, X.; Belmonte, R.; Rodrigo, A.; Rodà, F.; Piñol, J. Crecimiento Diametral de La Encina (Quercus ilex L.) En Un Año de Abundante Precipitación Estival: Efecto de La Irrigación Previa y de La Fertilización. Orsis Org. I Sistemes 1994, 9, 13–23. [Google Scholar]
- Balzano, A.; Čufar, K.; De Micco, V. Xylem and phloem formation dynamics in Quercus ilex L. at a dry site in Southern Italy. Forests 2021, 12, 188. [Google Scholar] [CrossRef]
- Campelo, F.; Gutiérrez, E.; Ribas, M.; Sánchez-Salguero, R.; Nabais, C.; Camarero, J.J. The facultative bimodal growth pattern in Quercus ilex—A simple model to predict sub-seasonal and inter-annual growth. Dendrochronologia 2018, 49, 77–88. [Google Scholar] [CrossRef]
- Louro, G.; Marques, H.; Salinas, F. Elementos de Apoio à Elaboração de Projectos Florestais; Direcção-Geral das Florestas: Lisboa, Portugal, 2002; ISBN 184/553/02. [Google Scholar]
- Martín, D.; Vázquez-Piqué, J.; Alejano, R. Effect of pruning and soil treatments on stem growth of holm oak in open woodland forests. Agrofor. Syst. 2015, 89, 599–609. [Google Scholar] [CrossRef]
- Ducrey, M.; Toth, J. Effect of cleaning and thinning on height growth and girth increment in holm oak coppices (Quercus ilex L.). Vegetatio 1992, 365–376. [Google Scholar] [CrossRef]
- Fabbio, G.; Cutini, A.; Mascia, V. Silvicultural treatment of Holm Oak (Quercus ilex L.) coppices in Southern Sardinia: Effects of canopy cover and thinning practices on inner microclimate. Ann.-Ist. Giangiacomo Feltrinelli 1998, 27, 55–63. [Google Scholar]
- Cabon, A.; Mouillot, F.; Lempereur, M.; Ourcival, J.-M.; Simioni, G.; Limousin, J.-M. Thinning increases tree growth by delaying drought-induced growth cessation in a Mediterranean evergreen oak coppice. For. Ecol. Manag. 2018, 409, 333–342. [Google Scholar] [CrossRef]
- Fukazawa, K. Juvenile Wood of Hardwoods Judged by Density Variation. IAWA J. 1984, 5, 65–73. [Google Scholar] [CrossRef]
- Sousa, V.; Louzada, J.; Pereira, H.; Sousa, V. Earlywood vessel features in Quercus faginea: Relationship between ring width and wood density at two sites in Portugal. iForest-Biogeosciences For. 2015, 8, 866–873. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.; Aebischer, D.; Denne, M. Latewood Density in Relation to Wood Fibre Diameter, Wall Thickness, and Fibre and Vessel Percentages in Quercus robur L. IAWA J. 1997, 18, 127–138. [Google Scholar] [CrossRef]
- Zhang, S.; Zhong, Y. Structure-property relationship of wood in East-Liaoning oak. Wood Sci. Technol. 1992, 26. [Google Scholar] [CrossRef]
- Leal, S.; Sousa, V.B.; Knapic, S.; Louzada, J.L.; Pereira, H. Vessel size and number are contributors to define wood density in cork oak. Eur. J. For. Res. 2011, 130, 1023–1029. [Google Scholar] [CrossRef]
- Fajardo, A. Insights into intraspecific wood density variation and its relationship to growth, height and elevation in a treeline species. Plant Biol. 2018, 20, 456–464. [Google Scholar] [CrossRef]
- Zhang, S. Mechanical properties in relation to specific gravity in 342 Chinese woods. Wood Fiber Sci. 2007, 26, 512–526. [Google Scholar]
- Degron, R.; Nepveu, G. Prévision de la variabilité intra- et interarbre de la densité du bois de chêne rouvre (Quercus petraea Liebl) par modélisation des largeurs et des densités des bois initial et final en fonction de l’âge cambial, de la largeur de cerne et du niveau dans l’arbre. Ann. For. Sci. 1996, 53, 1019–1030. [Google Scholar] [CrossRef]
- Guilley, E.; Hervé, J.-C.; Huber, F.; Nepveu, G. Modelling variability of within-ring density components in Quercus petraea Liebl. with mixed-effect models and simulating the influence of contrasting silvicultures on wood density. Ann. For. Sci. 1999, 56, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Sousa, V.; Ferreira, J.P.A.; Miranda, I.; Quilhó, T.; Pereira, H. Quercus rotundifolia Bark as a Source of Polar Extracts: Structural and Chemical Characterization. Forests 2021, 12, 1160. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Site 1 | Site 2 | |
|---|---|---|
| Latitude | 38°03′ N | 38°56′ N |
| Longitude | 07°06′ W | 08°07′ W |
| Altitude (m) | 450 | 130 |
| Soil | Dystric leptosols | Cambisols and vertic luvisols |
| Annual temperature (°C) | 15.9 | 16.0 |
| July–August temperature (°C) | 24.8 | 23.5 |
| January temperature (°C) | 9.4 | 9.1 |
| Annual precipitation (mm) | 614 | 555 |
| Tree height (m) | 6.1 ± 1.1 | 6.1 ± 1.7 |
| Diameter (cm) * | 21.5 ± 6.8 | 24.7 ± 6.6 |
| Crown height (m) ** | 5.2 ± 1.3 | 4.3 ± 1.7 |
| Radius crown (m) | 2.8 ± 0.7 | 2.7 ± 1.0 |
| Crown area (m) *** | 25.2 ± 11.5 | 25.8 ± 16.8 |
| Source of Variation | Degrees of Freedom | Expected Variance | Error Term |
|---|---|---|---|
| (1) Sites (S) | s − 1 | σ2ε + r σ2T/S + rt σ2 S | (2) |
| (2) Trees/S (T/S) | (t − 1) s | σ2ε + r σ2T/S | (5) |
| (3) Rings (R) | r − 1 | σ2ε + st σ2R | (5) |
| (4) Rings x Sites (R × S) | (r − 1) (s − 1) | σ2ε + t σ2RS | (5) |
| (5) Residual (R × T/S) | (r − 1) (t − 1) s | σ2ε |
| Tree | Rings | RW (mm) | RD (g/cm3) | MND (g/cm3) | MXD(g/cm3) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| # | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 |
| 1 | 52 | 60 | 1.74 ± 0.81 | 1.37 ± 1.30 | 0.879 ± 0.147 | 0.841 ± 0.158 | 0.799 ± 0.142 | 0.794 ± 0.157 | 0.960 ± 0.167 | 0.887 ± 0.170 |
| 2 | 45 | 68 | 1.40 ± 0.82 | 1.81 ± 1.20 | 0.878 ± 0.079 | 0.901 ± 0.133 | 0.807 ± 0.084 | 0.832 ± 0.155 | 0.959 ± 0.102 | 0.959 ± 0.129 |
| 3 | 33 | 73 | 2.89 ± 1.14 | 1.94 ± 1.44 | 0.873 ± 0.062 | 0.948 ± 0.185 | 0.797 ± 0.063 | 0.866 ± 0.182 | 0.963 ± 0.086 | 1.019 ± 0.198 |
| 4 | 35 | 83 | 1.77 ± 0.93 | 1.13 ± 1.14 | 0.983 ± 0.188 | 0.883 ± 0.230 | 0.922 ± 0.197 | 0.810 ± 0.217 | 1.046 ± 0.186 | 0.945 ± 0.246 |
| 5 | 57 | 58 | 1.81 ± 1.06 | 1.23 ± 1.06 | 0.881 ± 0.120 | 1.139 ± 0.183 | 0.814 ± 0.108 | 1.069 ± 0.207 | 0.952 ± 0.142 | 1.204 ± 0.167 |
| 6 | 31 | 78 | 2.18 ± 1.31 | 1.42 ± 0.86 | 0.944 ± 0.073 | 0.797 ± 0.143 | 0.871 ± 0.079 | 0.749 ± 0.145 | 1.034 ± 0.078 | 0.846 ± 0.146 |
| 7 | 70 | 84 | 2.05 ± 1.17 | 1.70 ± 1.37 | 0.880 ± 0.185 | 0.864 ± 0.193 | 0.777 ± 0.197 | 0.790 ± 0.203 | 0.973 ± 0.179 | 0.929 ± 0.194 |
| 8 | 46 | 61 | 1.27 ± 0.89 | 1.86 ± 1.49 | 1.069 ± 0.179 | 0.893 ± 0.239 | 0.986 ± 0.187 | 0.829 ± 0.237 | 1.143 ± 0.173 | 0.959 ± 0.251 |
| 9 | 81 | 59 | 1.57 ± 1.39 | 1.53 ± 1.27 | 0.834 ± 0.209 | 0.785 ± 0.159 | 0.769 ± 0.210 | 0.741 ± 0.150 | 0.905 ± 0.210 | 0.833 ± 0.178 |
| 10 | 61 | 59 | 1.84 ± 0.91 | 1.47 ± 0.96 | 0.997 ± 0.245 | 0.857 ± 0.248 | 0.941 ± 0.258 | 0.753 ± 0.239 | 1.055 ± 0.230 | 0.956 ± 0.267 |
| Mean | 51 | 68 | 1.81 ± 1.13 | 1.55 ± 1.24 | 0.914 ± 0.183 | 0.888 ± 0.210 | 0.840 ± 0.188 | 0.820 ± 0.209 | 0.991 ± 0.183 | 0.951 ± 0.219 |
| Site 2 | |||||
|---|---|---|---|---|---|
| Site 1 | RW | RD | MND | MXD | |
| RW | 1 | 0.766 | 0.729 | 0.823 | |
| RD | 0.516 | 1 | 0.961 | 0.989 | |
| MND | −0.909 | −0.583 | 1 | 0.945 | |
| MXD | −0.916 | −0.495 | 0.936 | 1 |
| Source of Variation | D.f. | RW | RD | MND | MXD | ||||
|---|---|---|---|---|---|---|---|---|---|
| Sig. | % | Sig. | % | Sig. | % | Sig. | % | ||
| S | 1 | ** | 0.4 | ns | 0.0 | ns | 0.0 | ns | 0.0 |
| T/S | 18 | *** | 7.3 | *** | 18.5 | *** | 17.0 | *** | 18.2 |
| R | 83 | *** | 14.4 | *** | 29.8 | *** | 27.4 | *** | 30.0 |
| S × R | 80 | *** | 10.0 | ns | 0.0 | ns | 0.0 | ns | 0.0 |
| R × T/S | 1011 | 67.9 | 51.7 | 55.6 | 51.9 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, V.; Silva, M.E.; Louzada, J.L.; Pereira, H. Wood Density and Ring Width in Quercus rotundifolia Trees in Southern Portugal. Forests 2021, 12, 1499. https://doi.org/10.3390/f12111499
Sousa V, Silva ME, Louzada JL, Pereira H. Wood Density and Ring Width in Quercus rotundifolia Trees in Southern Portugal. Forests. 2021; 12(11):1499. https://doi.org/10.3390/f12111499
Chicago/Turabian StyleSousa, Vicelina, Maria Emília Silva, José Luís Louzada, and Helena Pereira. 2021. "Wood Density and Ring Width in Quercus rotundifolia Trees in Southern Portugal" Forests 12, no. 11: 1499. https://doi.org/10.3390/f12111499