
Radial Growth Response of Picea crassifolia to Climatic Conditions in a Dryland Forest Ecosystem in Northwest China

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
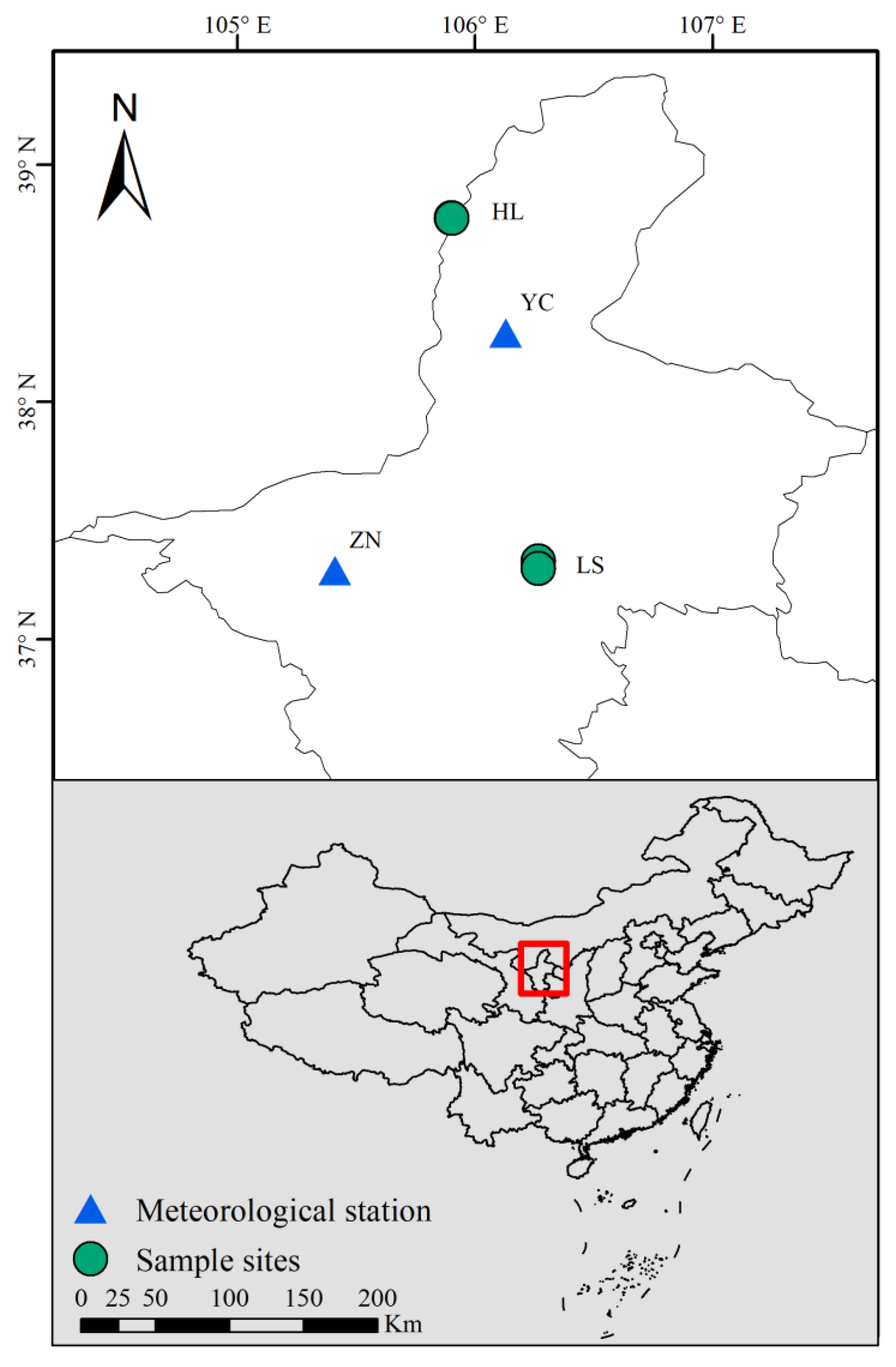
2.1. Study Area
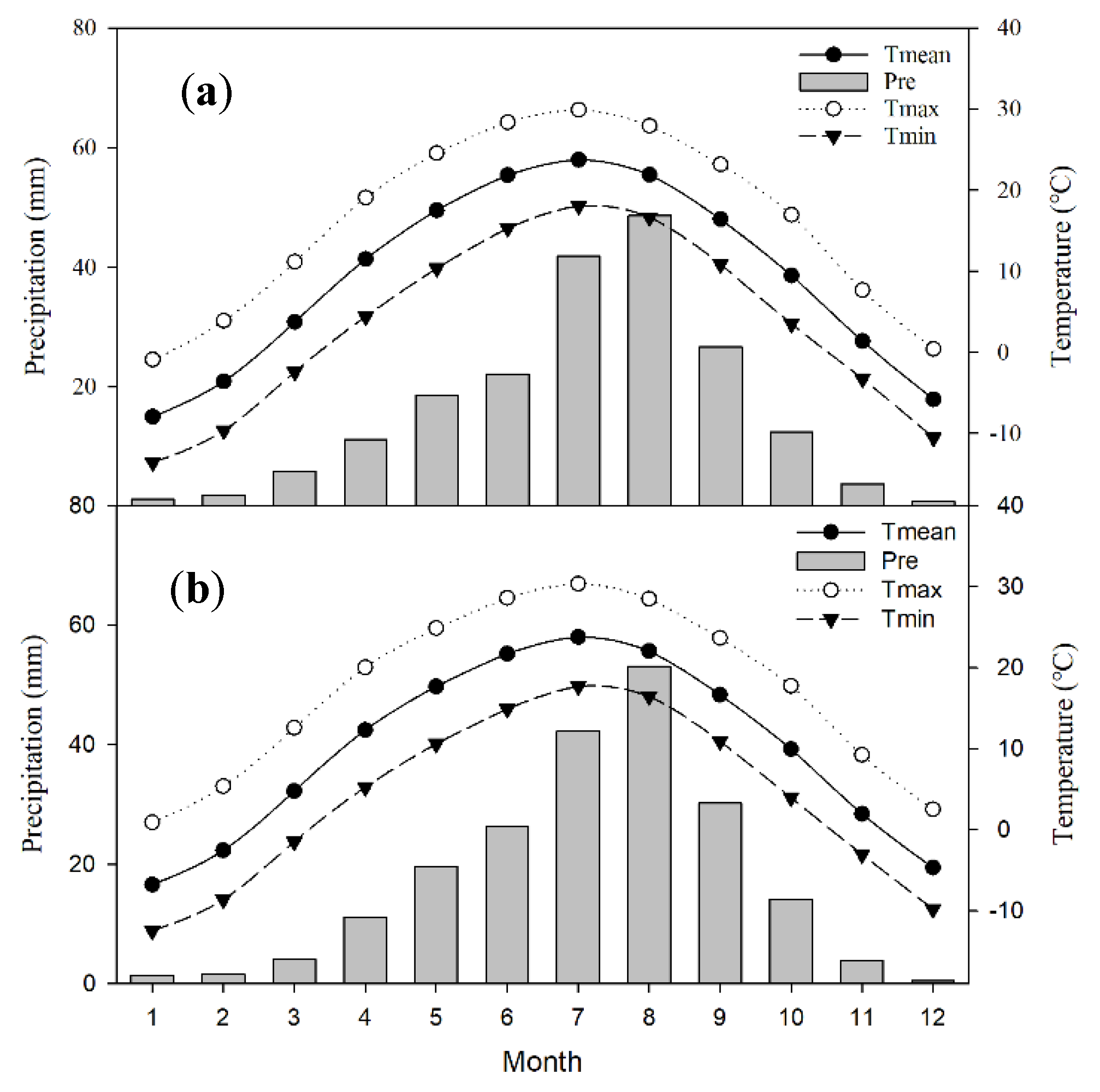
2.2. Meteorological Data
2.3. Tree-Ring Data
2.4. Data Analysis
3. Results
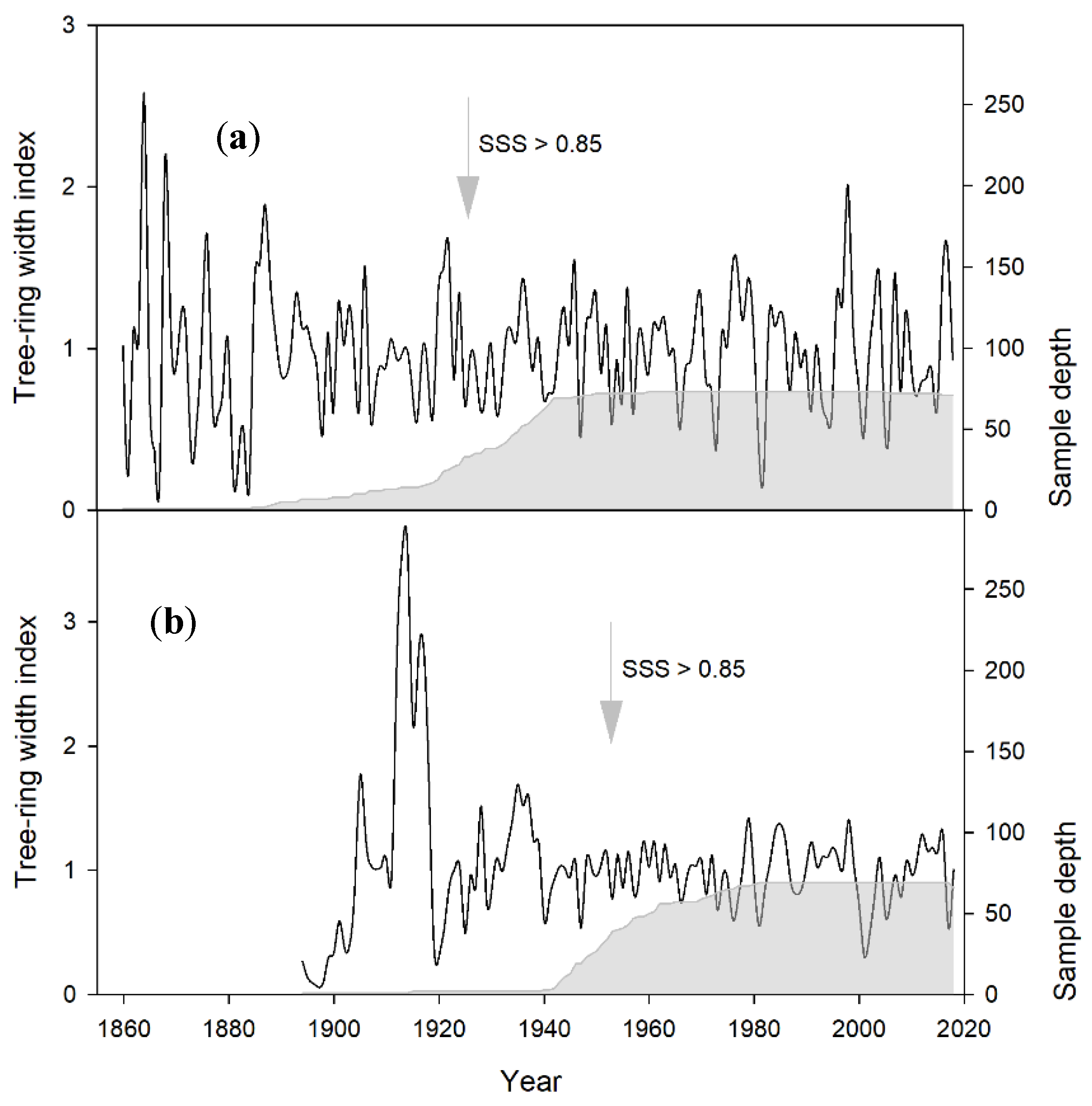
3.1. Statistical Parameters of the Chronologies
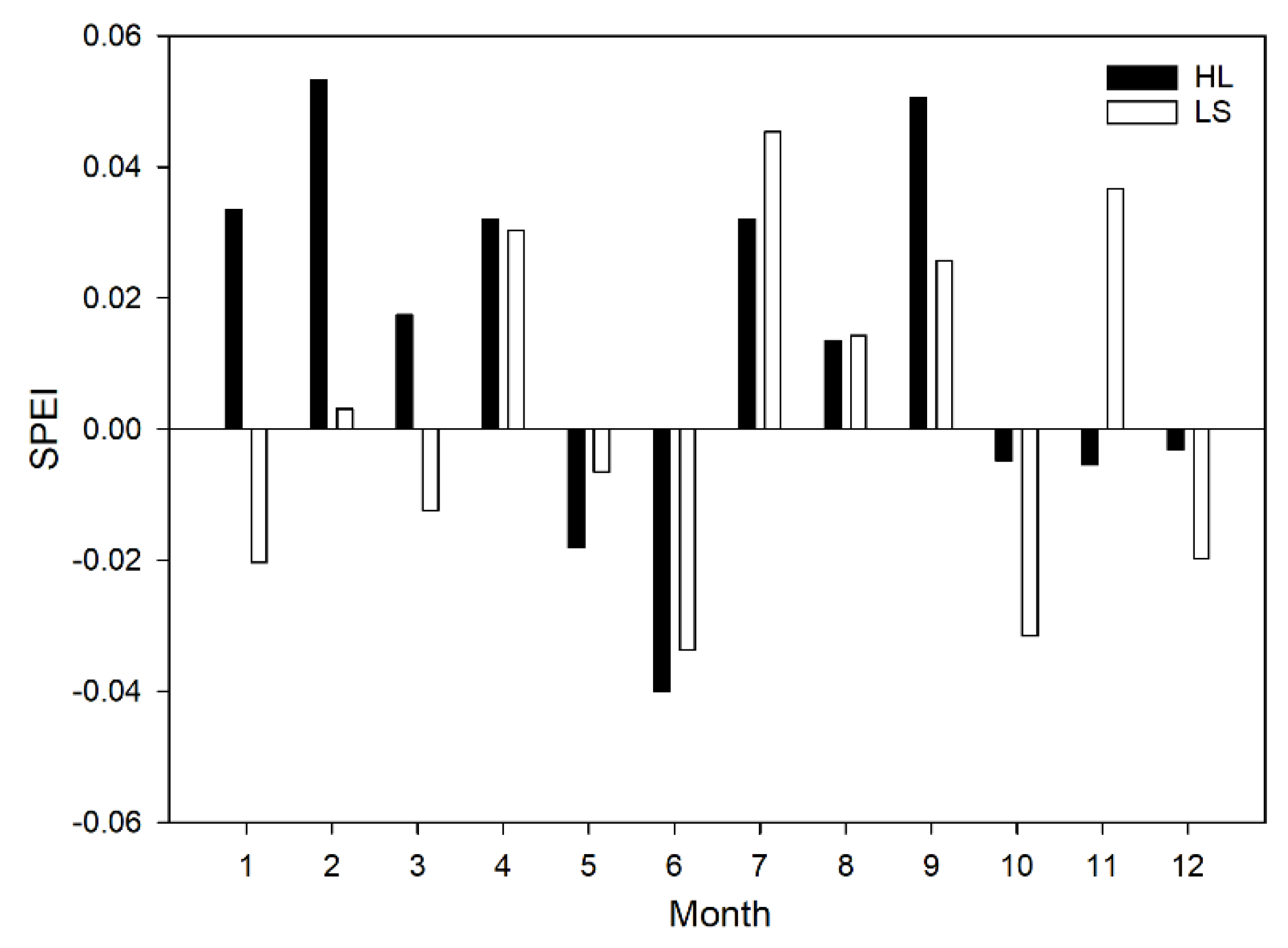
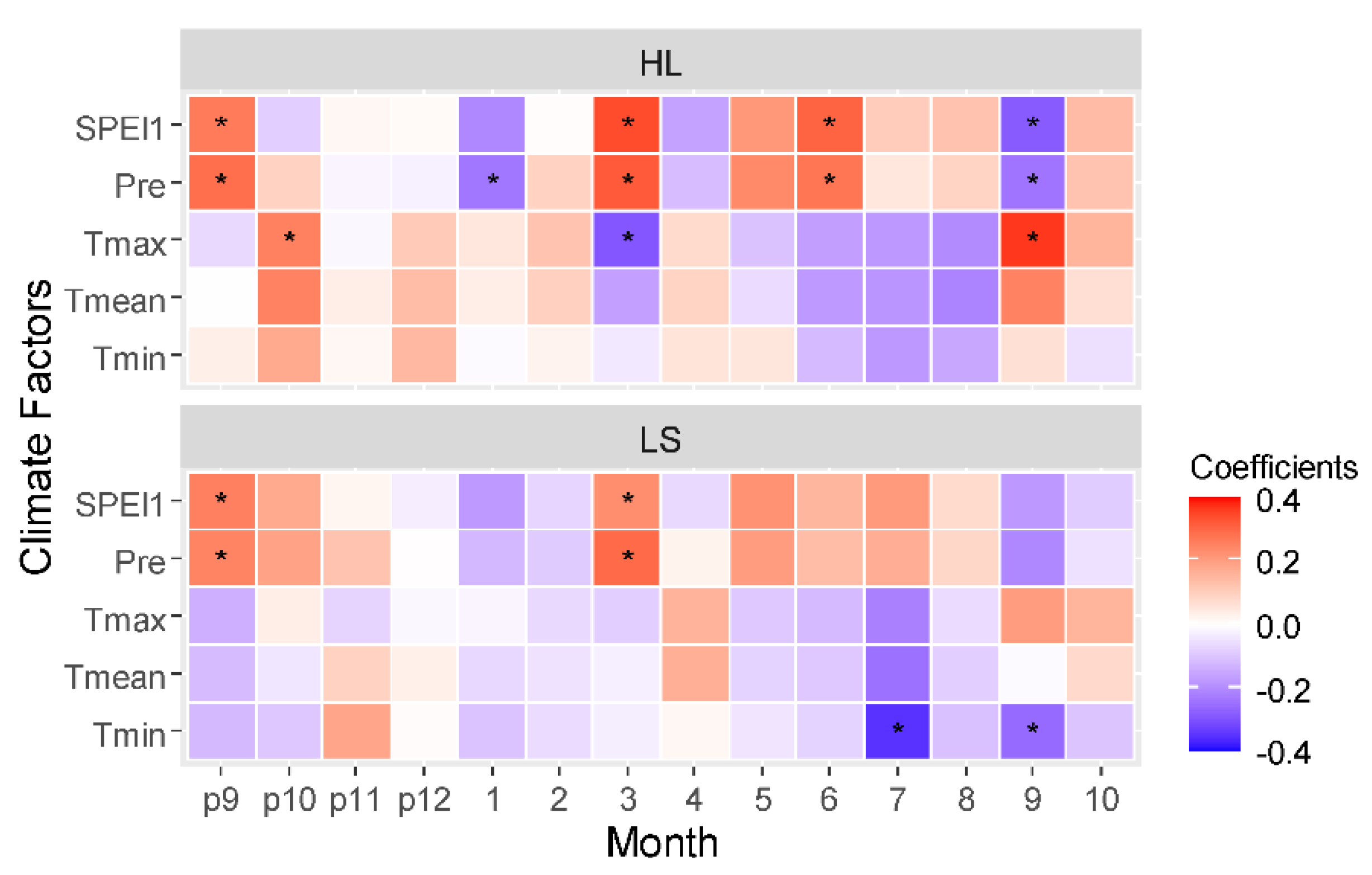
3.2. Growth–Climate Relationship
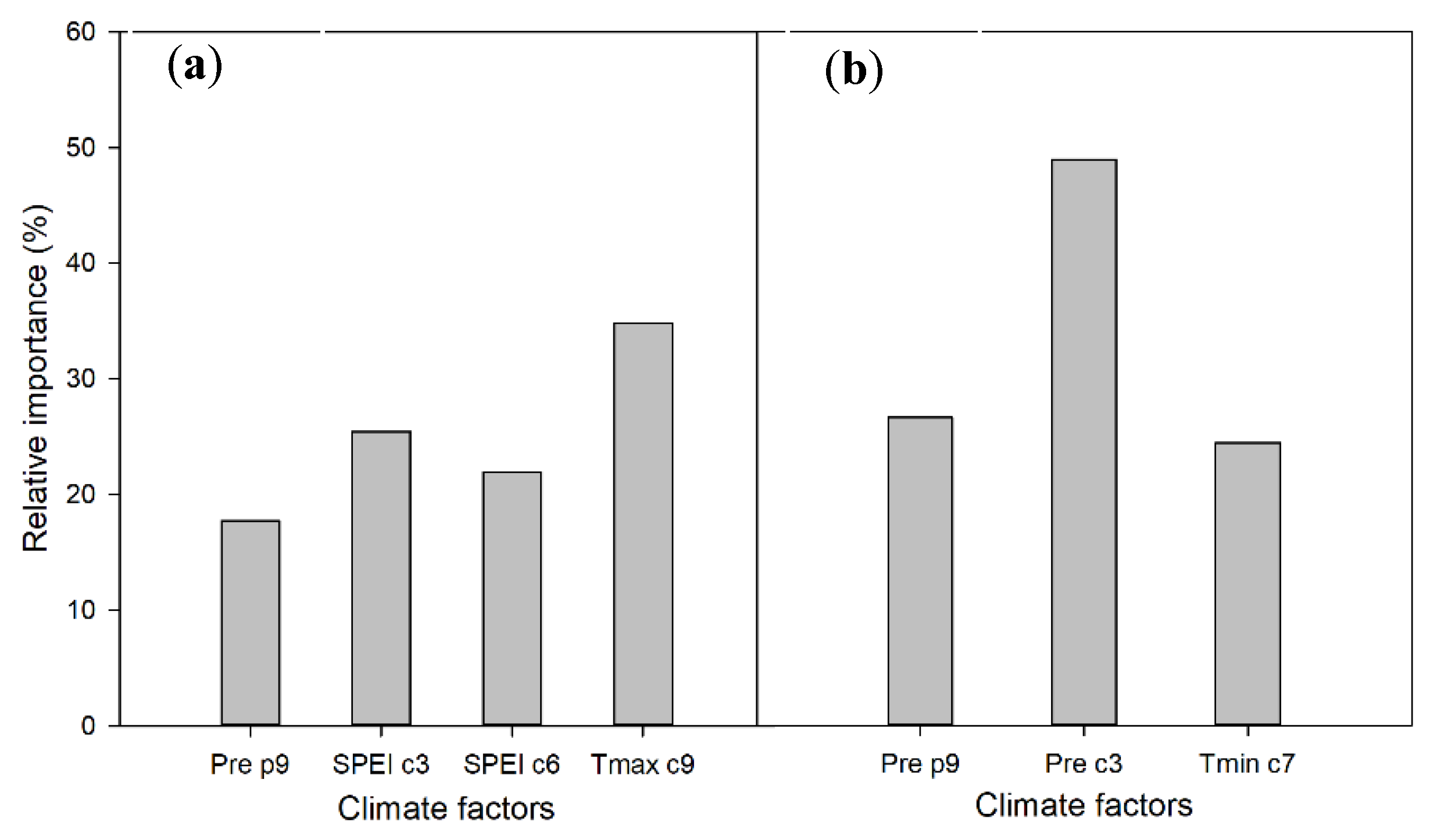
3.3. Multiple Regression and Relative Contributions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | HL1 | HL2 | HL3 | LS1 | LS2 |
|---|---|---|---|---|---|
| HL1 | 1 | 0.867165 | 0.871953 | 0.242871 | 0.276868 |
| HL2 | 1 | 0.801229 | 0.272293 | 0.303752 | |
| HL3 | 1 | 0.277903 | 0.326476 | ||
| LS1 | 1 | 0.764179 | |||
| LS2 | 1 |
References
- Zhang, J.; Huang, S.; He, F. Half-century evidence from western Canada shows forest dynamics are primarily driven by competition followed by climate. Proc. Natl. Acad. Sci. USA 2015, 112, 4009–4014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, E.; Wang, Y.; Piao, S.; Lu, X.; Camarero, J.J.; Zhu, H.; Zhu, L.; Ellison, A.M.; Ciais, P.; Penuelas, J. Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2016, 113, 4380–4385. [Google Scholar] [CrossRef] [Green Version]
- Assal, T.J.; Anderson, P.J.; Sibold, J. Spatial and temporal trends of drought effects in a heterogeneous semi-arid forest ecosystem. For. Ecol. Manag. 2016, 365, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, R.; Rossi, S.; Zhang, S.; Froment, I.; Huang, J.G.; Saracino, A. From phenology to forest management: Ecotypes selection can avoid early or late frosts, but not both. For. Ecol. Manag. 2019, 436, 21–26. [Google Scholar] [CrossRef]
- Ma, Q.; Huang, J.-G.; Hänninen, H.; Berninger, F. Divergent trends in the risk of spring frost damage to trees in Europe with recent warming. Glob. Chang. Biol. 2019, 25, 351–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadad, M.A.; Molina, J.A.; Juñent, F.A.R.; Amoroso, M.M.; Müller, G.; Araneo, D.; Tardif, J.C. Frost record in tree rings linked to atmospheric circulation in northern Patagonia. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 524, 201–211. [Google Scholar] [CrossRef]
- Chen, L.; Huang, J.-G.; Alam, S.A.; Zhai, L.; Dawson, A.; Stadt, K.J.; Comeau, P.G. Drought causes reduced growth of trembling aspen in western Canada. Glob. Chang. Biol. 2017, 23, 2887–2902. [Google Scholar] [CrossRef]
- Huang, J.; Tardif, J.C.; Bergeron, Y.; Denneler, B.; Berninger, F.; Girardin, M.P. Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest. Glob. Chang. Biol. 2010, 16, 711–731. [Google Scholar] [CrossRef]
- Chen, L.; Huang, J.; Dawson, A.; Zhai, L.; Stadt, K.J.; Comeau, P.G.; Whitehouse, C. Contributions of insects and droughts to growth decline of trembling aspen mixed boreal forest of western Canada. Glob. Chang. Biol. 2018, 24, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Robson, J.R.M.; Conciatori, F.; Tardif, J.C.; Knowles, K. Tree-ring response of jack pine and scots pine to budworm defoliation in central Canada. For. Ecol. Manag. 2015, 347, 83–95. [Google Scholar] [CrossRef]
- Shi, C.; Schneider, L.; Hu, Y.; Shen, M.; Sun, C.; Xia, J.; Forbes, B.C.; Shi, P.; Zhang, Y.; Ciais, P. Warming-induced unprecedented high-elevation forest growth over the monsoonal Tibetan Plateau. Environ. Res. Lett. 2020, 15, 054011. [Google Scholar] [CrossRef]
- Jiang, S.; Liang, H.; Zhou, P.; Wang, Z.; Zhu, H.; Kang, J.; Huang, J.-G. Spatial and temporal differences in the response of Larix sibirica to climate change in the central Altai Mountains. Dendrochronologia 2021, 67, 125827. [Google Scholar] [CrossRef]
- Qi, Z.; Liu, H.; Wu, X.; Hao, Q. Climate-driven speedup of alpine treeline forest growth in the Tianshan Mountains, Northwestern China. Glob. Chang. Biol. 2015, 21, 816–826. [Google Scholar] [CrossRef]
- Jochner, M.; Bugmann, H.; Nötzli, M.; Bigler, C. Tree growth responses to changing temperatures across space and time: A fine-scale analysis at the treeline in the Swiss Alps. Trees 2017, 32, 645–660. [Google Scholar] [CrossRef]
- Shi, Y.; Shen, Y.; Kang, E.; Li, D.; Ding, Y.; Zhang, G.; Hu, R. Recent and Future Climate Change in Northwest China. Clim. Chang. 2006, 80, 379–393. [Google Scholar] [CrossRef]
- Wang, T.; Ren, H.; Ma, K. Climatic signals in tree ring of Picea schrenkiana along an altitudinal gradient in the central Tianshan Mountains, northwestern China. Trees 2005, 19, 736–742. [Google Scholar] [CrossRef]
- Qin, L.; Yuan, Y.; Zhang, R.; Wei, W.; Yu, S.; Fan, Z.; Chen, F.; Zhang, T.; Shang, H. Tree-Ring Response to Snow Cover and Reconstruction of Century annual Maximum Snow Depth for Northern Tianshan Mountains, China. Geochronometria 2016, 43, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Yang, B.; Deslauriers, A.; Bräuning, A. Intra-annual stem radial increment response of Qilian juniper to temperature and precipitation along an altitudinal gradient in northwestern China. Trees 2015, 29, 25–34. [Google Scholar] [CrossRef]
- Gao, L.; Gou, X.; Deng, Y.; Liu, W.; Yang, M.; Zhao, Z. Climate-growth analysis of Qilian juniper across an altitudinal gradient in the central Qilian Mountains, northwest China. Trees 2013, 27, 379–388. [Google Scholar] [CrossRef]
- Gao, L.; Gou, X.; Deng, Y.; Yang, M.; Zhao, Z.; Cao, Z. Dendroclimatic Response of Picea crassifolia along an Altitudinal Gradient in the Eastern Qilian Mountains, Northwest China. Arctic Antarct. Alp. Res. 2013, 45, 491–499. [Google Scholar] [CrossRef]
- Li, B.Y.; Pan, B.T.; Cheng, W.M.; Han, J.F.; Qi, D.L.; Zhu, C. Research on geomorphologic regionalization of China. Acta Geogr. Sin. 2013, 68, 291–306, (In Chinese with English abstract). [Google Scholar]
- Liu, Y.; Cai, Q.; Shi, J.; Hughes, M.K.; Kutzbach, J.E.; Liu, Z.; Ni, F.; An, Z. Seasonal precipitation in the south-central Helan Mountain region, China, reconstructed from tree-ring width for the past 224 years. Can. J. For. Res. 2005, 35, 2403–2412. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y. January to August temperature variability since 1776 inferred from tree-ring width of Pinus tabulaeformis in Helan Mountain. J. Geogr. Sci. 2007, 17, 293–303. [Google Scholar] [CrossRef]
- Gao, J.; Yang, B.; He, M.; Shishov, V. Intra-annual stem radial increment patterns of Chinese pine, Helan Mountains, Northern Central China. Trees 2019, 33, 751–763. [Google Scholar] [CrossRef]
- Wang, Y.J.; Ma, Y.Z.; Zheng, Y.H.; Lu, R.J.; Meng, H.W. Response of tree-ring width of Pinus tabulaeformis to climate factors in Luoshan Mountains of Ningxia. J. Desert Res. 2009, 29, 971–976. [Google Scholar]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. J. Int. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Shen, Y.J. Spatio-temporal variations in extreme drought in China during 1961–2015. J. Geogr. Sci. 2019, 29, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Cook, E.R.; Holmes, R.L. Users Manual for Program ARSTAN; Laboratory of Tree-Ring Research, University of Arizona: Tucson, AZ, USA, 1986. [Google Scholar]
- Wigley, T.M.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendro-climatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Buras, A. A comment on the expressed population signal. Dendrochronologia 2017, 44, 130–132. [Google Scholar] [CrossRef]
- Biondi, F.; Waikul, K. DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Groemping, U. Relative Importance for Linear Regression in R: The Package relaimpo. J. Stat. Softw. 2006, 17, 925–933. [Google Scholar]
- Liu, Y.; Shi, J.; Shishov, V.; Vaganov, E.; Yang, Y.; Cai, Q.; Sun, J.; Wang, L.; Djanseitov, I. Reconstruction of May–July precipitation in the north Helan Mountain, Inner Mongolia since A.D. 1726 from tree-ring late-wood widths. Chin. Sci. Bull. 2004, 49, 405–409. [Google Scholar] [CrossRef]
- Fang, K.; Peng, J.; Zhang, Y.; Tian, Q.; Chen, F.; Gou, X. A 1232-year tree-ring record of climate variability in the Qilian Mountains, northestern China. IAWA J. 2009, 30, 407–420. [Google Scholar]
- Huang, J.; Deslauriers, A.; Rossi, S. Xylem formation can be modeled statistically as a function of primary growth and cambium activity. New Phytol. 2014, 203, 831–841. [Google Scholar] [CrossRef]
- Li, M.-M.; Li, G. Relationship between phenology of vegetation canopy and phenology of tree cambium in Helan Mountains, China. J. Appl. Ecol. 2021, 32, 495–502. [Google Scholar]
- Shi, J.F.; Liu, Y.; Cai, Q.F.; Yi, L. A case study of physiological characteristics of statistical correlation between Pinus tabulaeformis tree-ring widths and climatic factors. Acta Ecol. Sin. 2006, 26, 697–705. [Google Scholar]
- Wu, Z.; Yu, L.; Du, Z.; Zhang, H.; Fan, X.; Lei, T. Recent changes in the drought of China from 1960 to 2014. Int. J. Clim. 2019, 40, 3281–3296. [Google Scholar] [CrossRef]
- Richardson, S.; Dinwoodie, J. Studies on the physiology of xylem development. I. The effect of night temperature on tracheid size and wood density in conifers. J. Inst. Wood Sci. 1960, 6, 3–13. [Google Scholar]
- Hosoo, Y.; Yoshida, M.; Imai, T.; Okuyama, T. Diurnal difference in the amount of immunogold-labeled glucomannans detected with field emission scanning electron microscopy at the innermost surface of developing secondary walls of differentiating conifer tracheids. Planta 2002, 215, 1006–1012. [Google Scholar] [CrossRef]
- Liang, H.; Lyu, L.; Wahab, M. A 382-year reconstruction of August mean minimum temperature from tree-ring maximum latewood density on the southeastern Tibetan Plateau, China. Dendrochronologia 2016, 37, 1–8. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, Y.-J.; Wei, W.-S.; Yu, S.-L.; Zhang, T.-W.; Shang, H.-M.; Zhang, R.-B.; Qin, L.; Fan, Z.-A. Tree-ring recorded hydroclimatic change in Tienshan mountains during the past 500 years. Quat. Int. 2015, 358, 35–41. [Google Scholar] [CrossRef]






| Site Information | Common Period Analysis | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Site | Lat. (E) | Lon. (N) | Ele. (m) | Trees/Cores | Time Span | AR1 | Rbar | EPS | SNR |
| HL1 | 38°46′56″ | 105°54′06″ | 2608 | 13/23 | 1860–2018 | 0.428 | 0.688 | 0.972 | 34.648 |
| HL2 | 38°46′26″ | 105°54′07″ | 2660 | 9/18 | 1917–2018 | 0.781 | 0.735 | 0.984 | 31.1397 |
| HL3 | 38°46′25″ | 105°54′03″ | 2920 | 18/32 | 1885–2018 | 0.638 | 0.737 | 0.969 | 62.66 |
| LS1 | 37°18′00″ | 106°16′02″ | 2479 | 16/29 | 1944–2018 | 0.710 | 0.595 | 0.96 | 23.82 |
| LS2 | 37°19′48″ | 106°16′02″ | 2497 | 20/40 | 1894–2018 | 0.600 | 0.54 | 0.966 | 28.237 |
| HL | - | - | - | 40/73 | 1860–2018 | 0.633 | 0.66 | 0.989 | 93.085 |
| LS | - | - | - | 36/69 | 1894–2018 | 0.646 | 0.509 | 0.976 | 41.086 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, H.; Jiang, S.; Muhammad, A.; Kang, J.; Zhu, H.; Li, X.; Chen, L.; Zhu, L.; Huang, J.-G. Radial Growth Response of Picea crassifolia to Climatic Conditions in a Dryland Forest Ecosystem in Northwest China. Forests 2021, 12, 1382. https://doi.org/10.3390/f12101382
Liang H, Jiang S, Muhammad A, Kang J, Zhu H, Li X, Chen L, Zhu L, Huang J-G. Radial Growth Response of Picea crassifolia to Climatic Conditions in a Dryland Forest Ecosystem in Northwest China. Forests. 2021; 12(10):1382. https://doi.org/10.3390/f12101382
Chicago/Turabian StyleLiang, Hanxue, Shaowei Jiang, Ali Muhammad, Jian Kang, Huoxing Zhu, Xuebin Li, Lin Chen, Liangjun Zhu, and Jian-Guo Huang. 2021. "Radial Growth Response of Picea crassifolia to Climatic Conditions in a Dryland Forest Ecosystem in Northwest China" Forests 12, no. 10: 1382. https://doi.org/10.3390/f12101382