Internal Reference Gene Selection under Different Hormone Stresses in Multipurpose Timber Yielding Tree Neolamarckia cadamba

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Identification of Candidate Internal Reference Genes
2.3. Total RNA Isolation
2.4. cDNA Synthesis and Real-Time RT-PCR
2.5. Data Analysis
2.6. Validation of Internal Reference Genes
3. Results
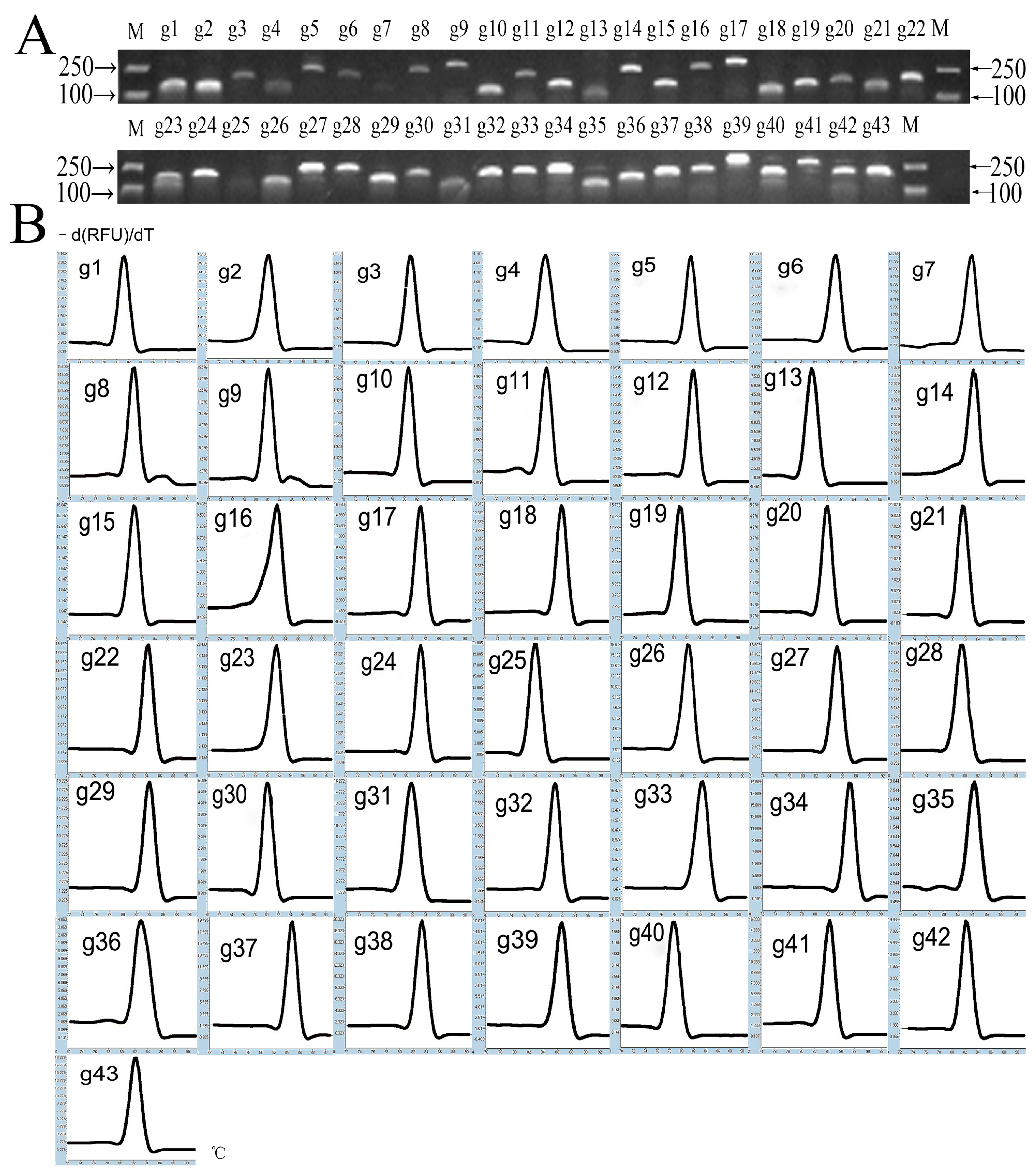
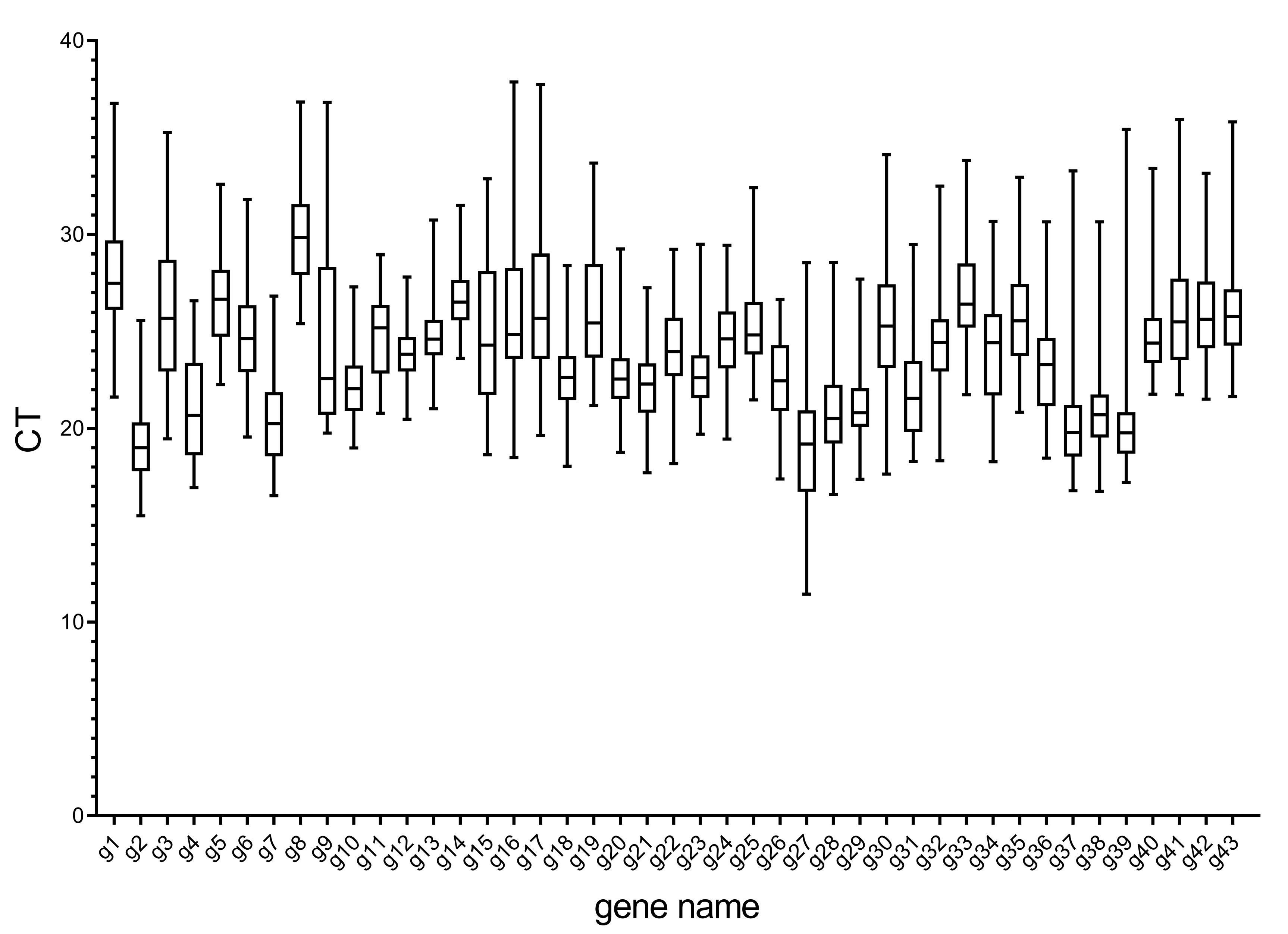
3.1. Primer Quality and CT Analysis of Candidate Internal Reference Genes
3.2. Analysis of Gene Expression Stability
3.3. Optimization of the Number of Reference Genes Required for RT-qPCR Analysis
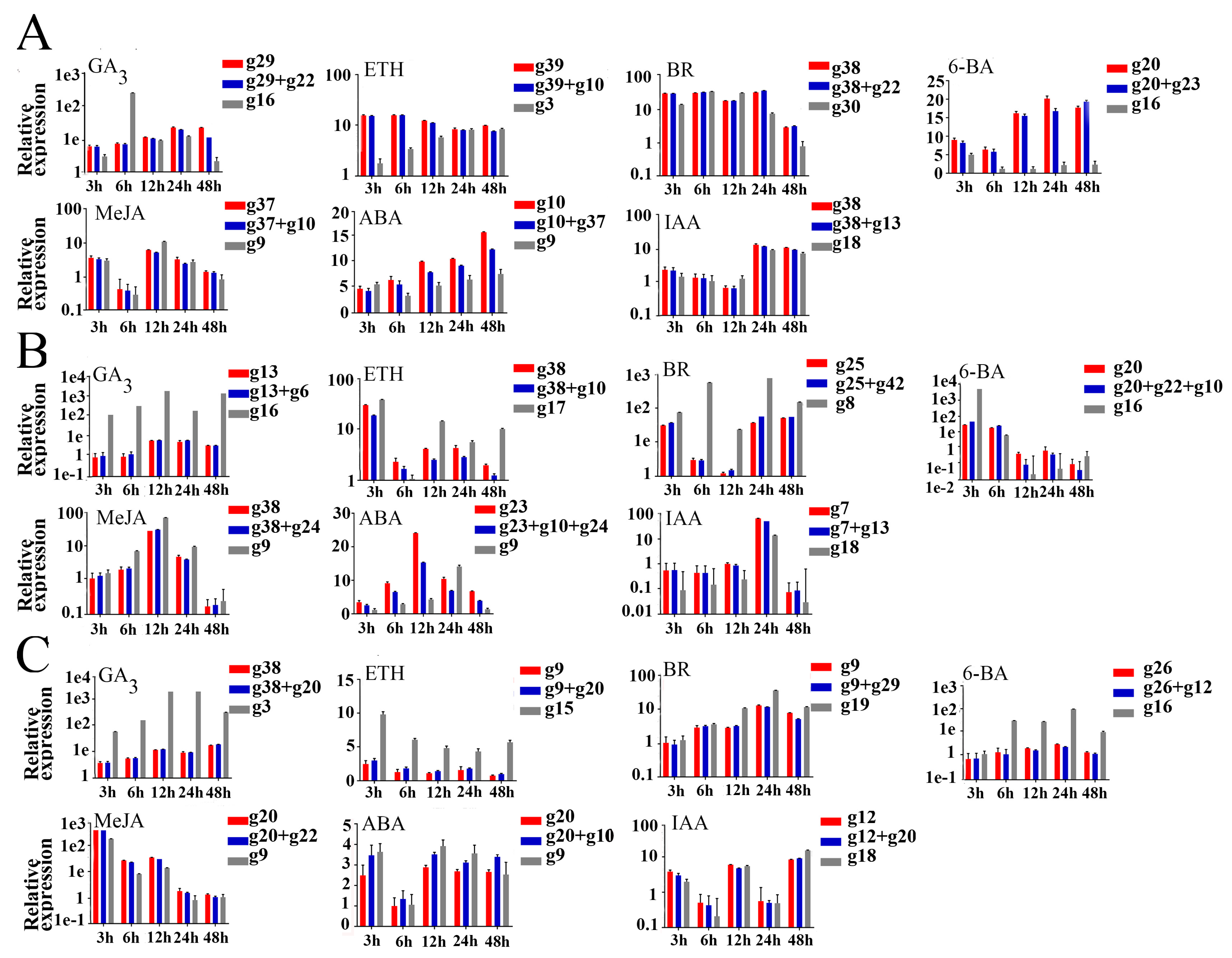
3.4. Validation of Selected Reference Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 6-BA | 6-Benzylaminopurine |
| ΔCt | Delta cycle threshold |
| ABA | Abscisic acid |
| ACT | Actin |
| APT | Adenine phosphoribosyl transferase |
| BR | Brassinolide |
| CAC | Clathrin adaptor complex medium |
| CTAB | Cetyltrimethylammonium bromide |
| CYP | Cyclophilin |
| DEPC | Diethyl pyrocarbonate |
| EF1α | Elongation factor 1α |
| eIF | Eukaryotic initiation factor |
| ETH | Ethephon |
| FBK | F-Boxkelch repeat protein |
| FPS1 | Farnesyl pyrophosphate synthase 1 |
| GA | Gibberellic acid |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| IAA | Indole-3-acetic acid |
| MeJA | Methyl Jasmonate |
| NCBI | National Center for Biotechnology Information |
| PEPKR1 | Phosphoenolpyruvate carboxylase-related kinase 1 |
| PP2 A | Protein phosphatase 2 A |
| RAN | GTP-binding nuclear protein |
| RPL | Ribosomal protein L |
| RPS | Ribosomal protein S |
| RuBP | Ribulose 1,5-bisphosphate carboxylase |
| SAMDC | S-adenosylmethionine decarboxylase |
| TEF | Translation elongation factor |
| Tub-α | Tubulin α |
| Tub-β | Tubulin β |
| UBCE | Ubiquitin conjugating enzyme |
| UBQ | Ubiquitin |
| UPL | Ubiquitin protein ligase |
References
- Chuaqui, R.F.; Bonner, R.F.; Best, C.J.; Gillespie, J.W.; Flaig, M.J.; Hewitt, S.M.; Phillips, J.L.; Krizman, D.B.; Tangrea, M.A.; Ahram, M.; et al. Post-analysis follow-up and validation of microarray experiments. Nat. Genet. 2002, 32, 509–514. [Google Scholar] [CrossRef]
- Die, J.V.; Roman, B.; Flores, F.; Rowland, L.J. Design and Sampling Plan Optimization for RT-qPCR Experiments in Plants: A Case Study in Blueberry. Front. Plant Sci. 2016, 7, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginzinger, D.G. Gene quantification using real-time quantitative PCR: An emerging technology hits the mainstream. Exp. Hematol. 2002, 30, 503–512. [Google Scholar] [CrossRef]
- Nolan, T.; Hands, R.E.; Bustin, S.A. Quantification of mRNA using real-time RT-PCR. Nat. Protoc. 2006, 1, 1559–1582. [Google Scholar] [CrossRef] [PubMed]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thellin, O.; Zorzi, W.; Lakaye, B.; De Borman, B.; Coumans, B.; Hennen, G.; Grisar, T.; Igout, A.; Heinen, E. Housekeeping genes as internal standards: Use and limits. J. Biotechnol. 1999, 75, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Long, J.; Liu, S.W.; Yang, Z.W.; Zhu, Q.J.; Zhao, X.L.; Peng, C. Selection and Validation of Reference Genes for mRNA Expression by Quantitative Real-Time PCR Analysis in Neolamarckia cadamba. Sci. Rep. 2018, 8, 9311. [Google Scholar] [CrossRef]
- Jain, N.; Vergish, S.; Khurana, J.P. Validation of house-keeping genes for normalization of gene expression data during diurnal/circadian studies in rice by RT-qPCR. Sci. Rep. 2018, 8, 3203. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.Q.; Fortunato, A.S.; Rodrigues, W.P.; Partelli, F.L.; Campostrini, E.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Selection and Validation of Reference Genes for Accurate RT-qPCR Data Normalization in Coffea spp. under a Climate Changes Context of Interacting Elevated [CO2] and Temperature. Front. Plant Sci. 2017, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Lu, M.; Chen, Y.; Zhan, Z.; Cui, Q.; Wang, Y. Selection of reliable reference genes for gene expression studies using real-time PCR in tung tree during seed development. PLoS ONE 2012, 7, e43084. [Google Scholar] [CrossRef]
- Guenin, S.; Mauriat, M.; Pelloux, J.; Van Wuytswinkel, O.; Bellini, C.; Gutierrez, L. Normalization of qRT-PCR data: The necessity of adopting a systematic, experimental conditions-specific, validation of references. J. Exp. Bot. 2009, 60, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, H34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Li, J.; Hu, X.; Huang, X.; Huo, H.; Li, J.; Zhang, D.; Li, P.; Ouyang, K.; Chen, X. Functional identification of an EXPA gene (NcEXPA8) isolated from the tree Neolamarckia cadamba. Biotechnol. Biotechnol. Equip. 2017, 31, 1116–1125. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, K.X.; Liu, M.Q.; Pian, R.Q.; Liu, S.S.; Chen, X.Y. Isolation and analysis of alpha-expansin genes in the tree Anthocephalus chinensis (Rubiaceae). Genet. Mol. Res. 2013, 12, 1061–1073. [Google Scholar] [CrossRef]
- Pandey, A.; Negi, P.S. Phytochemical composition, in vitro antioxidant activity and antibacterial mechanisms of Neolamarckia cadamba fruits extracts. Nat. Prod. Res. 2018, 32, 1189–1192. [Google Scholar] [CrossRef]
- Chandel, M.; Kumar, M.; Sharma, U.; Kumar, N.; Singh, B.; Kaur, S. Isolation and characterization of flavanols from Anthocephalus cadamba and evaluation of their antioxidant, antigenotoxic, cytotoxic and COX-2 inhibitory activities. Rev. Bras. Farmacogn. 2016, 26, 474–483. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Negi, P.S. Traditional uses, phytochemistry and pharmacological properties of Neolamarckia cadamba: A review. J. Ethnopharmacol. 2016, 181, 118–135. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Pavlovic, I.; Pencik, A.; Novak, O.; Vujcic, V.; Radic, B.S.; Lepedus, H.; Strnad, M.; Salopek-Sondi, B. Short-term salt stress in Brassica rapa seedlings causes alterations in auxin metabolism. Plant Physiol. Biochem. 2018, 125, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Song, L.X.; Xu, X.C.; Wang, F.N.; Wang, Y.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Zhou, J.; Yu, J.Q. Brassinosteroids act as a positive regulator for resistance against root-knot nematode involving RESPIRATORY BURST OXIDASE HOMOLOG-dependent activation of MAPKs in tomato. Plant Cell Environ. 2018, 41, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, R.; Zhang, H. Effects of plant growth regulators (DA-6 and 6-BA) and EDDS chelator on phytoextraction and detoxification of cadmium by Amaranthus hybridus Linn. Int. J. Phytoremediat. 2018, 20, 1121–1128. [Google Scholar] [CrossRef]
- Sabater-Jara, A.B.; Almagro, L.; Pedreno, M.A. Induction of extracellular defense-related proteins in suspension cultured-cells of Daucus carota elicited with cyclodextrins and methyl jasmonate. Plant Physiol. Biochem. 2014, 77, 133–139. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Rep. 2013, 32, 959–970. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Ouyang, K.; Li, J.; Huang, H.; Que, Q.; Li, P.; Chen, X. A simple method for RNA isolation from various tissues of the tree Neolamarckia cadamba. Biotechnol. Biotechnol. Equip. 2014, 28, 1008–1013. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, K.; Li, J.; Zhao, X.; Que, Q.; Li, P.; Huang, H.; Deng, X.; Singh, S.K.; Wu, A.M.; Chen, X. Transcriptomic Analysis of Multipurpose Timber Yielding Tree Neolamarckia cadamba during Xylogenesis Using RNA-Seq. PLoS ONE 2016, 11, e159407. [Google Scholar] [CrossRef]
- Shcherban, T.Y.; Shi, J.; Durachko, D.M.; Guiltinan, M.J.; McQueen-Mason, S.J.; Shieh, M.; Cosgrove, D.J. Molecular cloning and sequence analysis of expansins—A highly conserved, multigene family of proteins that mediate cell wall extension in plants. Proc. Natl. Acad. Sci. USA 1995, 92, 9245–9249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampedro, J.; Cosgrove, D.J. The expansin superfamily. Genome Biol. 2005, 6, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santner, A.; Estelle, M. Recent advances and emerging trends in plant hormone signalling. Nature 2009, 459, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.J. Oxidative stress: The paradox of aerobic life. Biochem. Soc. Symp. 1995, 61, 1–31. [Google Scholar]
- Shigenaga, A.M.; Argueso, C.T. No hormone to rule them all: Interactions of plant hormones during the responses of plants to pathogens. Semin. Cell Dev. Biol. 2016, 56, 174–189. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Y.; Li, S.S.; Han, G.Z. Insights into the origin and evolution of the plant hormone signaling machinery. Plant Physiol. 2015, 167, 872–886. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.R.; Nam, H.Y.; Kim, S.U.; Kim, S.I.; Chang, Y.J. Normalization of reverse transcription quantitative-PCR with housekeeping genes in rice. Biotechnol. Lett. 2003, 25, 1869–1872. [Google Scholar] [CrossRef]
- Murthi, P.; Fitzpatrick, E.; Borg, A.J.; Donath, S.; Brennecke, S.P.; Kalionis, B. GAPDH, 18S rRNA and YWHAZ are suitable endogenous reference genes for relative gene expression studies in placental tissues from human idiopathic fetal growth restriction. Placenta 2008, 29, 798–801. [Google Scholar] [CrossRef]
- Sikela, J.M.; Auffray, C. Finding new genes faster than ever. Nat. Genet. 1993, 3, 189–191. [Google Scholar] [CrossRef]
- Bai, J.F.; Wang, Y.K.; Wang, P.; Yuan, S.H.; Gao, J.G.; Duan, W.J.; Wang, N.; Zhang, F.T.; Zhang, W.J.; Qin, M.Y.; et al. Genome-wide identification and analysis of the COI gene family in wheat (Triticum aestivum L.). BMC Genomics 2018, 19, 754. [Google Scholar] [CrossRef]
- Choi, D.; Lee, Y.; Cho, H.T.; Kende, H. Regulation of expansin gene expression affects growth and development in transgenic rice plants. Plant Cell 2003, 15, 1386–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kende, H.; Van der Knaap, E.; Cho, H.T. Deepwater rice: A model plant to study stem elongation. Plant Physiol. 1998, 118, 1105–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | UniGene ID | Reference Gene ID | F/R Primer (5′—3′) | Amplicator Length (bp) | Efficiency (%) | Correlation Coefficient (R2) |
|---|---|---|---|---|---|---|
| ACT | comp52737_c0 | g1a | ATGTTGAAGCCTGTTCCATTGT | 114 | 97.6 | 0.997 |
| TAACTAATAACAGAAGCATTCATCCA | ||||||
| comp79635_c0 | g2 | CTTCTGAGGTTATGGAGCAATCT | 101 | 105 | 0.993 | |
| CGATAAATCAAAACTTCAAGCC | ||||||
| CAC | comp48976_c0 | g3 | CTCAGAGAACGCTGCTGACTAC | 161 | 104.8 | 0.996 |
| GAGCCAAGGGAAACAAGATAA | ||||||
| CYP | comp67418_c0 | g4 | GGGGTCTCACGCTCTTTACT | 83 | 96.8 | 0.993 |
| GGATTGGATTGGGTTGGTT | ||||||
| comp75463_c0 | g5 | CCCCAGCAAGAAGACCACT | 213 | 102.1 | 0.996 | |
| TTGACCATGAATCCCAACCA | ||||||
| comp77969_c0 | g6 | ATAGCATCCCAACCGAACA | 187 | 102.1 | 0.997 | |
| CCCTCTTGCCTCCTGTGTAT | ||||||
| EF1α | comp87079_c1 | g7 | ACCAGCATCACCGTTCTTCA | 123 | 98 | 0.996 |
| GTCCTCGATTGCCACACCT | ||||||
| comp87526_c0 | g8 | AATCAGACAGAAACCCCTCAA | 245 | 101.8 | 0.994 | |
| GAACCTCTCAATCACACGCTT | ||||||
| eIF | comp6386_c0 | g9 | GTTGAAACTTCTTGGACATCG | 250 | 103.3 | 0.991 |
| CTTGAGACACTGATTTGTATGAGA | ||||||
| FPS1 | comp72548_c0 | g10 | TGATAATCTGGCTTCCACCTT | 112 | 103.6 | 0.992 |
| TGGGAGGAACTCAATCTCCTAC | ||||||
| comp75377_c0 | g11 | TATCAGGCTCAGCATTCCACT | 212 | 101.6 | 0.994 | |
| TTGCCACAATAACACATCCAT | ||||||
| FBK | comp78454_c0 | g12 | AAGGCCAATTCTGTTCAAGC | 143 | 96.1 | 0.997 |
| CCTAGAGGGAAAGACATGACTG | ||||||
| comp78817_c0 | g13 | GCAAACGGGGTAAAAGGA | 102 | 99.6 | 0.993 | |
| AAAGGGTAAGAGTGACGACAGC | ||||||
| GAPDH | comp78593_c0 | g14 | TGTTCCAAGTGGGCATTTAC | 247 | 104.3 | 0.99 |
| CGCTCTGAGGTGTTAATAAGTG | ||||||
| comp80828_c1 | g15 | CTGAGCATTTTTTAGGCTTGTC | 151 | 103.5 | 0.992 | |
| TCAGATTCATGTGGCAGTCG | ||||||
| RAN | comp85262_c0 | g16 | TCTCGCAACCTGCCTCTT | 257 | 101.2 | 0.99 |
| TATCACTCCCATCTTCGCAC | ||||||
| PEPKR1 | comp75525_c0 | g17 | CGACCTCACATTCCTCATTAC | 291 | 97.7 | 0.995 |
| ACATAGACCATCCAGAGCCCA | ||||||
| comp80613_c0 | g18 | TACATAGACCATCCAGAGCCA | 112 | 102.4 | 0.991 | |
| GCAAAAGGGCAAGCAACAG | ||||||
| PP2A | comp81334_c1 | g19 | CTGGGTGGGAAAGATGTG | 142 | 104.6 | 0.995 |
| CTTGGGCAATAGGCTGAC | ||||||
| comp52412_c0 | g20 | ATGTTGGATGATATTAGTGGTGTG | 161 | 100.3 | 0.992 | |
| TCATAGGAAAATAGACCTCTGGTT | ||||||
| RPL | comp46755_c0 | g21 | CTGAGGATTGTTAGCAGTTGAC | 119 | 103.4 | 0.993 |
| ACCAGAAAACAGACCACCTAAG | ||||||
| comp52434_c0 | g22 | AAGGAAGGTAAAGCAGGGAA | 177 | 98.4 | 0.995 | |
| GCATGGGCAGGGATATAAAC | ||||||
| comp87976_c0 | g23 | CACGCAGCATAGCCAAAC | 157 | 104.5 | 0.991 | |
| AGGCAGTTCTCTGATTCTTTTG | ||||||
| RPS | comp65909_c1 | g24 | GCTATGGTAGTCTCCCGAAAG | 182 | 104.4 | 0.992 |
| GGGGGAACAAGACTAAGGGT | ||||||
| comp67276_c0 | g25 | TTTTGTTTCCCCTCTTTGC | 97 | 102.6 | 0.991 | |
| AACCTTGAACAACCTGTGTAGAA | ||||||
| comp71526_c0 | g26 | CGGTTACACAAGGTTGAATGA | 117 | 104.5 | 0.996 | |
| AGAGGGTCTGGATTTGAGTGA | ||||||
| RuBP | comp47386_c0 | g27 | CAGCACCGTAATCCATAAAAC | 226 | 104.6 | 0.993 |
| CAAGCAGCCCAGCAAGTC | ||||||
| comp88001_c0 | g28 | ACAGGATGGGTAGAAAGAGGC | 210 | 104.9 | 0.996 | |
| AGGATTGAGCCGAATACAACG | ||||||
| SAMDC | comp44802_c0 | g29 | TCTTCGTGGCACTTCTCTCC | 133 | 101.5 | 0.993 |
| ACAGGGTGTTGACTTGTTTCC | ||||||
| comp71874_c0 | g30 | ATAAGGTCTCTTCTTGTTCGTGTAG | 178 | 103.5 | 0.994 | |
| GACTGAACAGCAACAGGAATAAT | ||||||
| comp80075_c0 | g31 | GCTGCCTGTGGGTCTCCTA | 85 | 104.8 | 0.998 | |
| GTAAACCCCAATGCTACTCCT | ||||||
| TEF | comp65909_c1 | g32 | GCTATGGTAGTCTCCCGAAAG | 184 | 100.5 | 0.995 |
| CTGGGGGAACAAGACTAAGG | ||||||
| comp70791_c0 | g33 | TCAACCAACCGTTCCTACC | 195 | 105.2 | 0.99 | |
| ACAACAGTCCTTTGCCACC | ||||||
| Tub-α | comp70323_c2 | g34 | GGTGGTGGAACTGGCTCTG | 217 | 103.3 | 0.993 |
| GGCAAATGTCATAGATGGCTT | ||||||
| comp76448_c4 | g35 | AAGGAGGGAATGAGTGGAG | 107 | 103.4 | 0.99 | |
| ACTATGGCAAGAAGTCAAAGC | ||||||
| Tub-β | comp66056_c0 | g36 | GCAAGAAAGCCTTCCTCCTAA | 153 | 103.9 | 0.999 |
| TTCCCAACAATGTCAAATCAA | ||||||
| comp79707_c1 | g37 | TTCAGGAGAGTCAGCGAGC | 187 | 100.4 | 0.999 | |
| CATCGTCTTCATATTCCCCTT | ||||||
| UBCE | comp79182_c1 | g38 | TCCTTGCTTGTGGCGTCA | 213 | 105.8 | 0.999 |
| CACGGGTGTCAAATCTGGC | ||||||
| UBQ | comp67366_c0 | g39 | GACGGGAGGACCTTAGCA | 298 | 105.5 | 0.993 |
| CTCGGAGACGGAGAACAA | ||||||
| comp82561_c0 | g40 | GCATTTGTGTCTTGCCTCTTTAT | 186 | 105.5 | 0.993 | |
| GCGATGAGCAACATTCCTTTA | ||||||
| comp75872_c1 | g41 | TCTTGAAGGGAATGGTGTTTTG | 267 | 105.3 | 0.993 | |
| AGATGTTAGGAGGACTGAGGAT | ||||||
| UPL | comp87122_c0 | g42 | GGTTGGTGGTAGAGTTGTGACTC | 182 | 105.8 | 0.995 |
| CGAGCACTACCACGACACG | ||||||
| comp88840_c0 | g43 | CTGCTCGTTGGTATGTAATGG | 128 | 104.3 | 0.99 | |
| TCAGGCAATCCAAAGACAACT |
| Treatments | Buds (Most Stable/Least Stable) | Leaves (Most Stable/Least Stable) | Cambium Region (Most Stable/Least Stable) | Total (Most Stable/Least Stable) |
|---|---|---|---|---|
| GA3 | g29/g16 | g13/g16 | g38/g3 | g20/g16 |
| ETH | g39/g3 | g38/g17 | g20/g15 | g20/g17 |
| BR | g42/g30 | g25/g8 | g22/g19 | g9/g27 |
| 6-BA | g20/g16 | g20/g16 | g12/g16 | g20/g39 |
| MeJA | g37/g9 | g38/g9 | g20/g9 | g38/g9 |
| ABA | g10/g9 | g23/g9 | g20/g9 | g38/g9 |
| IAA | g38/g18 | g7/g18 | g12/g18 | g7/g27 |
| Total (most stable/least stable) | g10/g17 | g23/g9 | g38/g9 | g10/g17 |
| Hormone | Tissue | |||
|---|---|---|---|---|
| Bud | Leaf | Cambium Region | Total | |
| 6-BA | g20, g23 | g20, g22, g10 | g26, g12 | g20, g12, g10 |
| ABA | g10, g37 | g23, g10, g24 | g20, g10 | g38, g20, g10 |
| BR | g38, g22 | g25, g42 | g9, g29 | g9, g38, g42 |
| ETH | g39, g10 | g38, g10 | g9, g20 | g20, g38, g10 |
| GA3 | g29, g22 | g13, g6 | g38, g20 | g20, g38, g29 |
| IAA | g38, g13 | g7, g13 | g12, g20 | g7, g13, g10 |
| MeJA | g37, g10 | g38, g24 | g20, g22 | g38, g20, g10 |
| Total | g10, g13, g29 | g23, g13, g29 | g38, g39, g23 | g10, g23, g12 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Li, J.; Li, B.; Li, C.; Chen, X.; Ouyang, K. Internal Reference Gene Selection under Different Hormone Stresses in Multipurpose Timber Yielding Tree Neolamarckia cadamba. Forests 2020, 11, 1014. https://doi.org/10.3390/f11091014
Zhang D, Li J, Li B, Li C, Chen X, Ouyang K. Internal Reference Gene Selection under Different Hormone Stresses in Multipurpose Timber Yielding Tree Neolamarckia cadamba. Forests. 2020; 11(9):1014. https://doi.org/10.3390/f11091014
Chicago/Turabian StyleZhang, Deng, Jingjian Li, Buye Li, Chunmei Li, Xiaoyang Chen, and Kunxi Ouyang. 2020. "Internal Reference Gene Selection under Different Hormone Stresses in Multipurpose Timber Yielding Tree Neolamarckia cadamba" Forests 11, no. 9: 1014. https://doi.org/10.3390/f11091014