Litter Decomposition and Nutrient Dynamics in Fire-Affected Larch Forests in the Russian Far East


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Characteristics
2.2. Litterbag Experiment
2.3. Mass Loss
2.4. Litter Respiration
2.5. Chemical Analysis
2.6. Statistical Analysis
3. Results
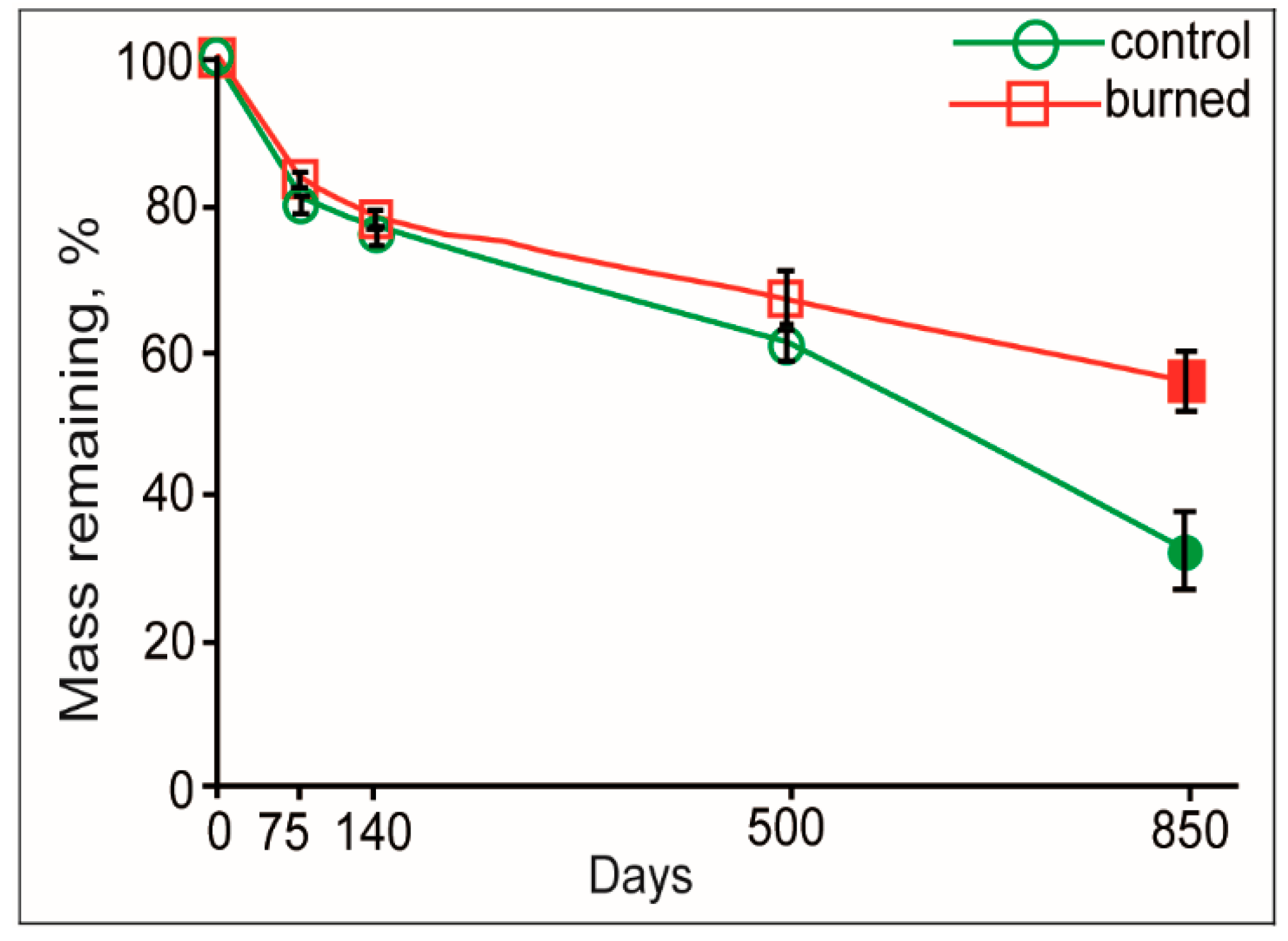
3.1. Mass Loss and Decomposition Constant
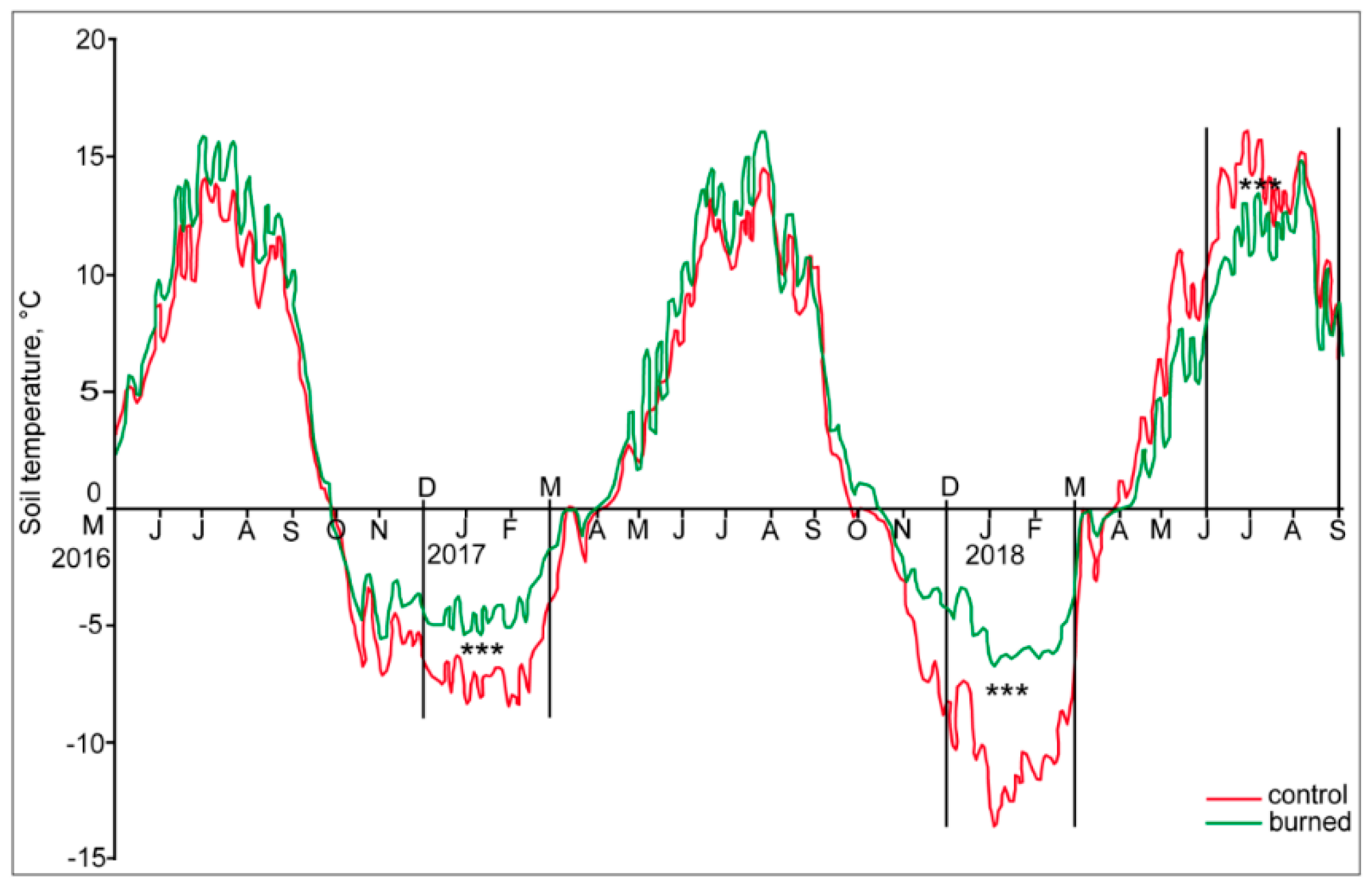
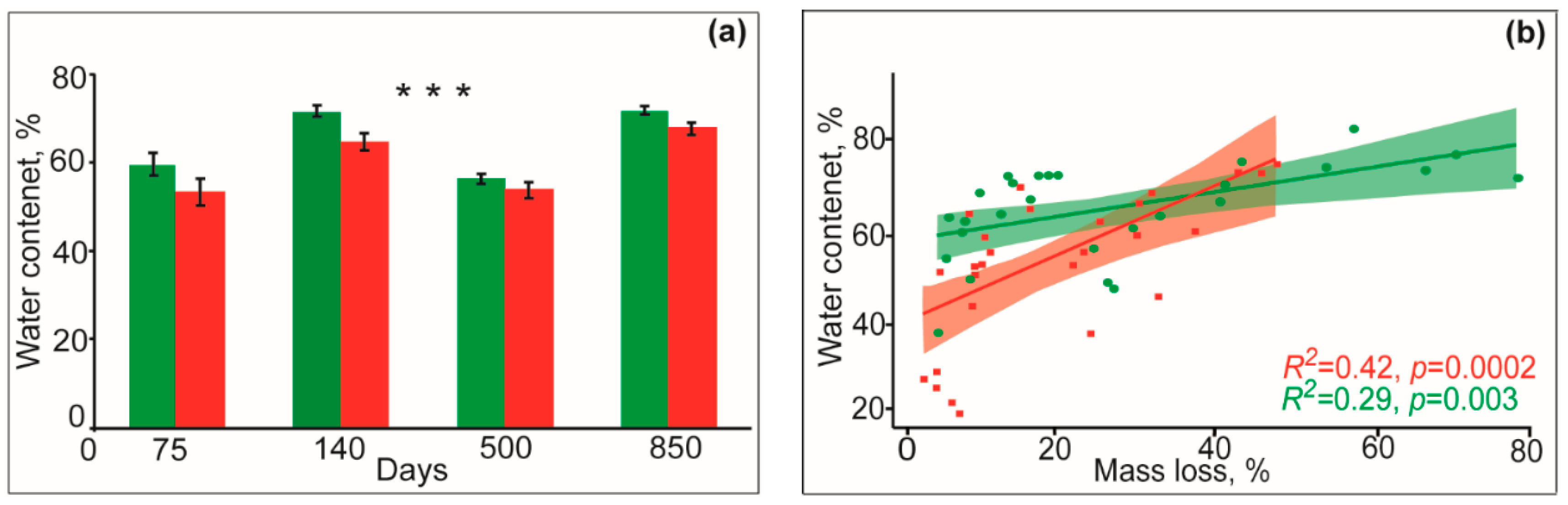
3.2. Soil Temperature and Moisture
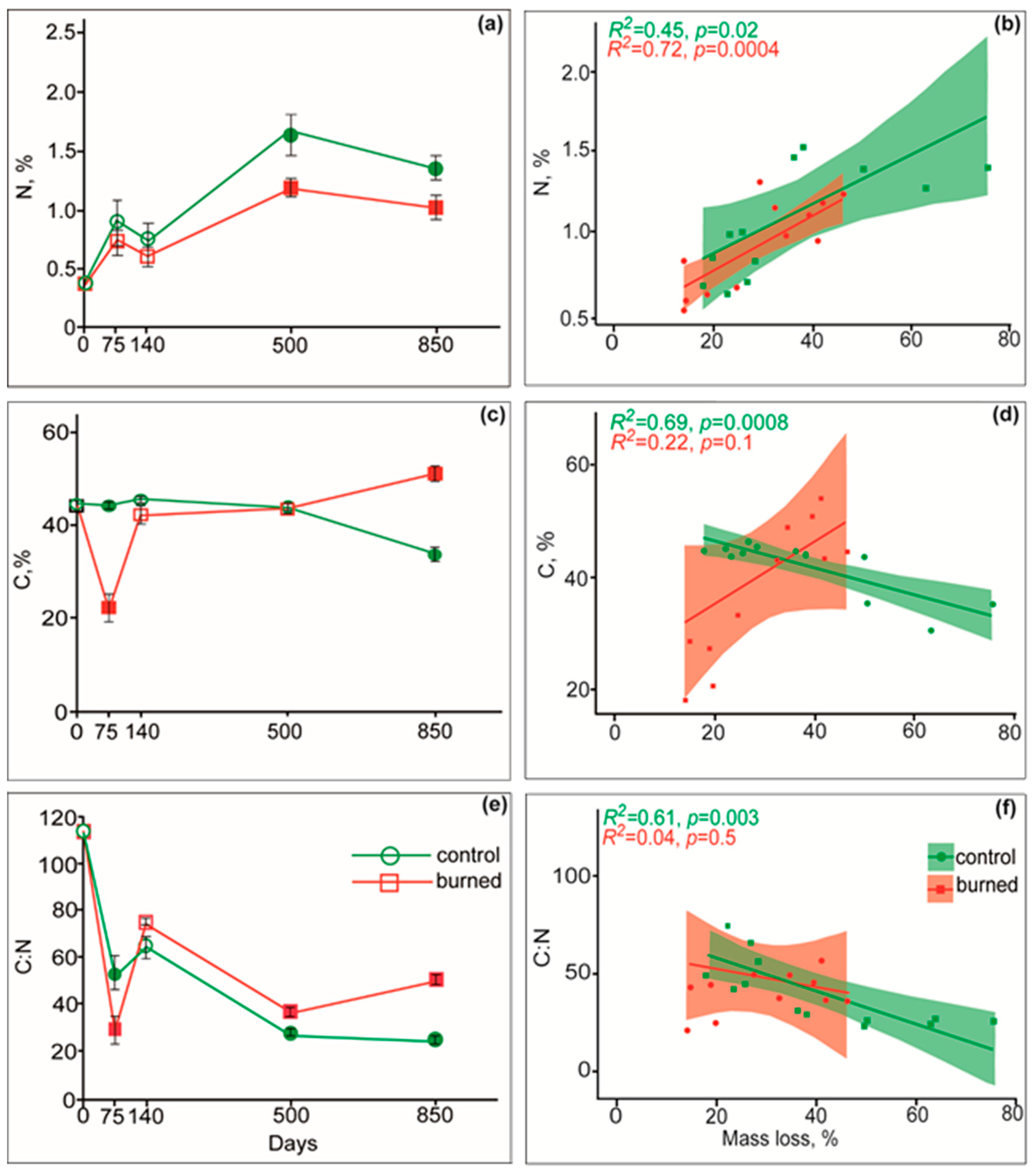
3.3. Dynamics of C, N, and C:N Ratio
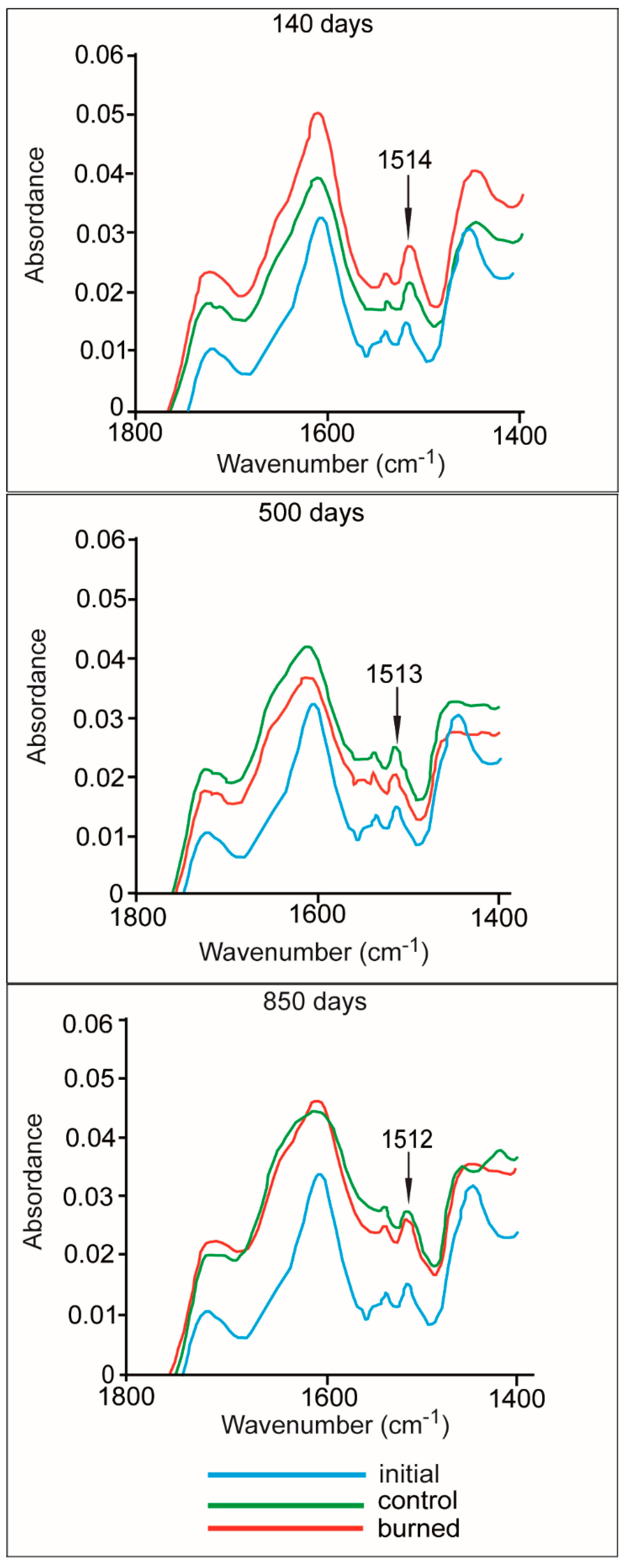
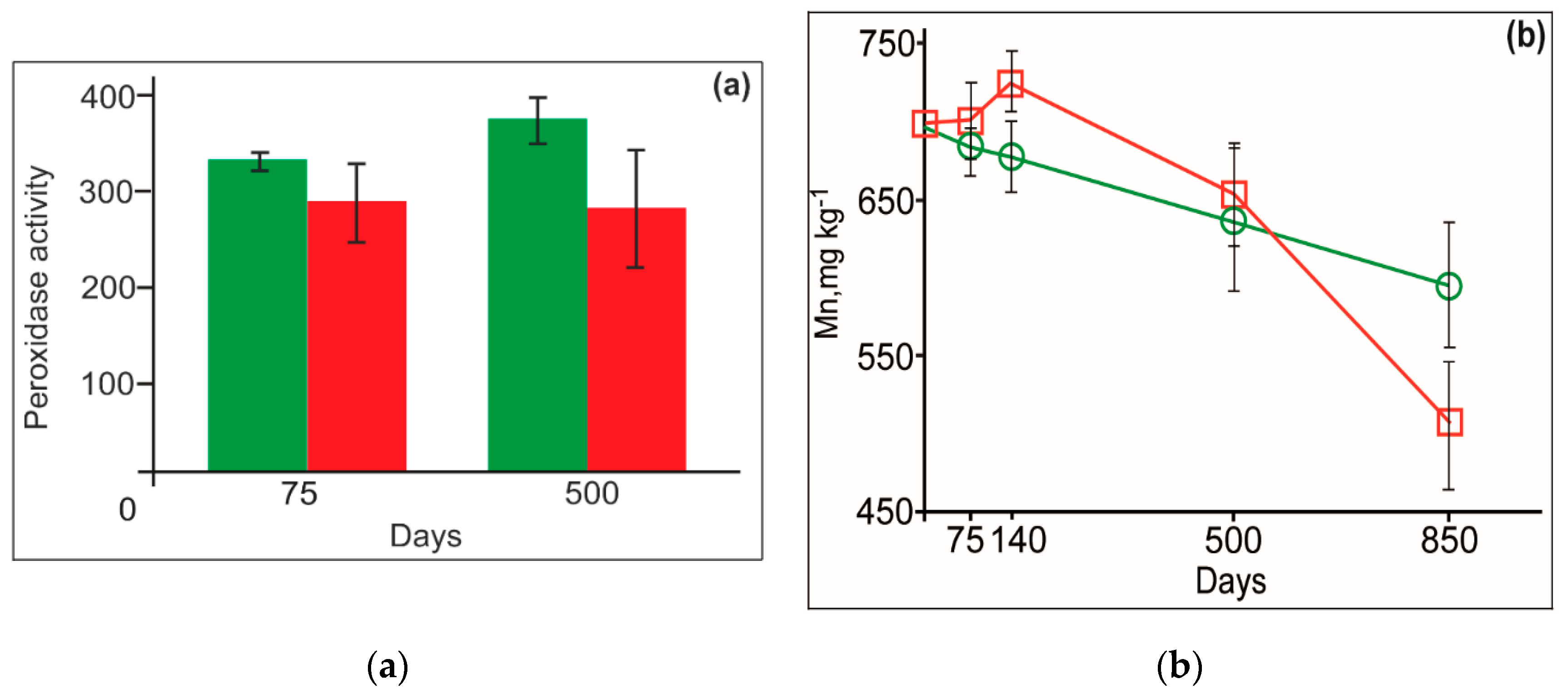
3.4. Lignin and Mn Dynamics
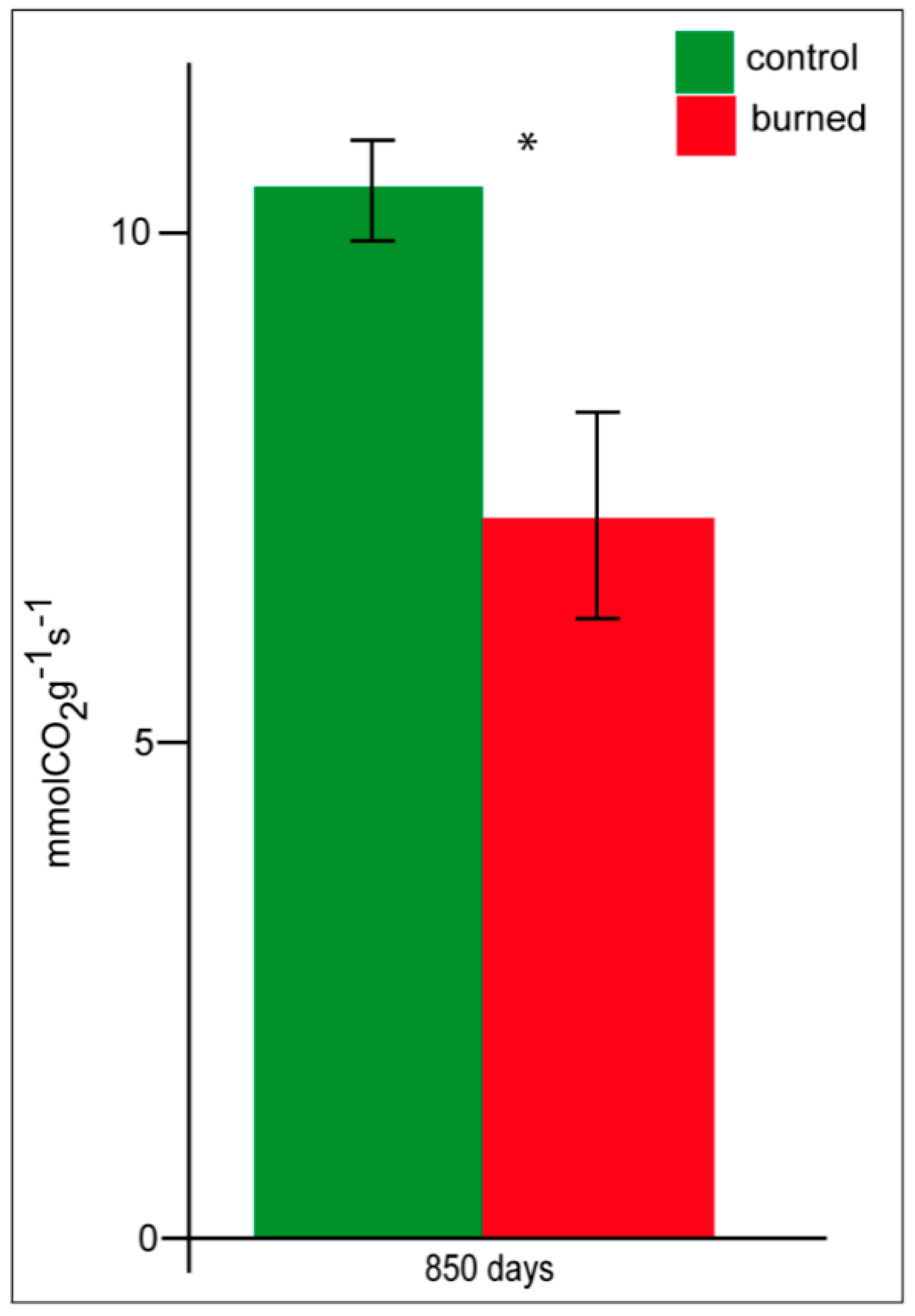
3.5. Litter Respiration and Enzyme Activities
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pimm, S.; Roulet, N.; Weaver, A. Boreal forests’ carbon stores need better management. Nature 2009, 462, 276. [Google Scholar] [CrossRef] [PubMed]
- Tarnocai, C.; Canadell, J.G.; Schuur, E.A.G.; Kuhry, P.; Mazhitova, G.; Zimov, S. Soil organic carbon pools in the northern circumpolar permafrost region. Global Biogeochem. Cycles 2009, 23. [Google Scholar] [CrossRef]
- DeLuca, T.H.; Boisvenue, C. Boreal forest soil carbon: Distribution, function and modelling. Forestry 2012, 85, 161–184. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef]
- Lawrence, D.M.; Slater, A.G.; Romanovsky, V.E.; Nicolsky, D.J. Sensitivity of a model projection of near-surface permafrost degradation to soil column depth and representation of soil organic matter. J. Geophys. Res. 2008, 113, F02011. [Google Scholar] [CrossRef] [Green Version]
- Kasischke, E.S.; Christensen, N.L.; Stocks, B.J. Fire, Global Warming, and the Carbon Balance of Boreal Forests. Ecol. Appl. 1995, 5, 437–451. [Google Scholar] [CrossRef]
- Schaphoff, S.; Reyer, C.P.O.; Schepaschenko, D.; Gerten, D.; Shvidenko, A. Tamm Review: Observed and projected climate change impacts on Russia’s forests and its carbon balance. For. Ecol. Manag. 2015, 361, 432–444. [Google Scholar] [CrossRef] [Green Version]
- Dove, N.C.; Safford, H.D.; Bohlman, G.N.; Estes, B.L.; Hart, S.C. High-severity wildfire leads to multi-decadal impacts on soil biogeochemistry in mixed-conifer forests. Ecol. Appl. 2020. [Google Scholar] [CrossRef]
- Wardle, D.A.; Nilsson, M.-C.; Zackrisson, O.; Gallet, C. Determinants of litter mixing effects in a Swedish boreal forest. Soil Biol. Biochem. 2003, 35, 827–835. [Google Scholar] [CrossRef]
- Bani, A.; Pioli, S.; Ventura, M.; Panzacchi, P.; Borruso, L.; Tognetti, R.; Tonon, G.; Brusetti, L. The role of microbial community in the decomposition of leaf litter and deadwood. Appl. Soil Ecol. 2018, 126, 75–84. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-3-642-38820-0. [Google Scholar]
- Perez, J.; Jeffries, T.W. Roles of manganese and organic acid chelators in regulating lignin degradation and biosynthesis of peroxidases by Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1992, 58, 2402–2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valášková, V.; Šnajdr, J.; Bittner, B.; Cajthaml, T.; Merhautová, V.; Hofrichter, M.; Baldrian, P. Production of lignocellulose-degrading enzymes and degradation of leaf litter by saprotrophic basidiomycetes isolated from a Quercus petraea forest. Soil Biol. Biochem. 2007, 39, 2651–2660. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Gough, L. Litter decomposition in moist acidic and non-acidic tundra with different glacial histories. Oecologia 2004, 140, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Averill, C.; Waring, B. Nitrogen limitation of decomposition and decay: How can it occur? Glob. Chang. Biol. 2018, 24, 1417–1427. [Google Scholar] [CrossRef] [Green Version]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Bradford, M.A.; Berg, B.; Maynard, D.S.; Wieder, W.R.; Wood, S.A. Understanding the dominant controls on litter decomposition. J. Ecol. 2016, 104, 229–238. [Google Scholar] [CrossRef]
- Moore, T.R.; Trofymow, J.A.; Siltanen, M.; Prescott, C.; Group, C.W. Patterns of decomposition and carbon, nitrogen, and phosphorus dynamics of litter in upland forest and peatland sites in central Canada. Can. J. For. Res. 2005, 35, 133–142. [Google Scholar] [CrossRef]
- Berg, B.; Söderström, B. Fungal biomass and nitrogen in decomposing scots pine needle litter. Soil Biol. Biochem. 1979, 11, 339–341. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. NITROGEN MINERALIZATION: CHALLENGES OF A CHANGING PARADIGM. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- García-Palacios, P.; Shaw, E.A.; Wall, D.H.; Hättenschwiler, S. Contrasting mass-ratio vs. niche complementarity effects on litter C and N loss during decomposition along a regional climatic gradient. J. Ecol. 2017, 105, 968–978. [Google Scholar] [CrossRef] [Green Version]
- Wan, S.; Hui, D.; Luo, Y. Fire Effects on Nitrogen Pools and Dynamics in Terrestrial Ecosystems: A Meta-Analysis. Ecol. Appl. 2001, 11, 1349. [Google Scholar] [CrossRef]
- Stirling, E.; Smernik, R.J.; Macdonald, L.M.; Cavagnaro, T.R. Fire influences needle decomposition: Tipping point in Pinus radiata carbon chemistry and soil nitrogen transformations. Soil Biol. Biochem. 2019, 135, 361–368. [Google Scholar] [CrossRef]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on belowground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Switzer, J.M.; Hope, G.D.; Grayston, S.J.; Prescott, C.E. Changes in soil chemical and biological properties after thinning and prescribed fire for ecosystem restoration in a Rocky Mountain Douglas-fir forest. For. Ecol. Manag. 2012, 275, 1–13. [Google Scholar] [CrossRef]
- Hart, S.C.; DeLuca, T.H.; Newman, G.S.; MacKenzie, M.D.; Boyle, S.I. Post-fire vegetative dynamics as drivers of microbial community structure and function in forest soils. For. Ecol. Manag. 2005, 220, 166–184. [Google Scholar] [CrossRef]
- Berg, B. Litter decomposition and organic matter turnover in northern forest soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Holden, S.R.; Gutierrez, A.; Treseder, K.K. Changes in Soil Fungal Communities, Extracellular Enzyme Activities, and Litter Decomposition Across a Fire Chronosequence in Alaskan Boreal Forests. Ecosystems 2013, 16, 34–46. [Google Scholar] [CrossRef] [Green Version]
- Köster, K.; Berninger, F.; Heinonsalo, J.; Lindén, A.; Köster, E.; Ilvesniemi, H.; Pumpanen, J. The long-term impact of low-intensity surface fires on litter decomposition and enzyme activities in boreal coniferous forests. Int. J. Wildl. Fire 2016, 25, 213. [Google Scholar] [CrossRef]
- Liechty, H.O.; Reinke, M. The influence of repeated prescribed fire on decomposition and nutrient release in uneven-aged loblolly–shortleaf pine stands. Fire Ecol. 2020, 16, 6. [Google Scholar] [CrossRef]
- Throop, H.L.; Abu Salem, M.; Whitford, W.G. Fire enhances litter decomposition and reduces vegetation cover influences on decomposition in a dry woodland. Plant Ecol. 2017, 218, 799–811. [Google Scholar] [CrossRef]
- Preston, C.M.; Schmidt, M.W.I. Black (pyrogenic) carbon: A synthesis of current knowledge and uncertainties with special consideration of boreal regions. Biogeosciences 2006, 3, 397–420. [Google Scholar] [CrossRef] [Green Version]
- Makoto, K.; Hirobe, M.; DeLuca, T.H.; Bryanin, S.V.; Procopchuk, V.F.; Koike, T. Effects of fire-derived charcoal on soil properties and seedling regeneration in a recently burned Larix gmelinii/Pinus sylvestris forest. J. Soils Sediments 2011, 11, 1317–1322. [Google Scholar] [CrossRef]
- Bryanin, S.; Abramova, E.; Makoto, K. Fire-derived charcoal might promote fine root decomposition in boreal forests. Soil Biol. Biochem. 2018, 116, 1–3. [Google Scholar] [CrossRef]
- Permafrost Ecosystems; Osawa, A.; Zyryanova, O.A.; Matsuura, Y.; Kajimoto, T.; Wein, R.W. (Eds.) Ecological Studies; Springer: Dordrecht, The Netherlands, 2010; Volume 209, ISBN 978-1-4020-9692-1. [Google Scholar]
- Kobak, K.I.; Turchinovich, I.Y.; Kondrasheva, N.Y.; Schulze, E.D.; Schulze, W.; Koch, H.; Vygodskaya, N.N. Vulnerability and adaptation of the larch forest in Eastern Siberia to climate change. Water. Air. Soil Pollut. 1996, 92, 119–127. [Google Scholar] [CrossRef]
- Kajimoto, T.; Matsuura, Y.; Sofronov, M.a.; Volokitina, a.V.; Mori, S.; Osawa, a.; Abaimov, a.P. Above- and belowground biomass and primary productivity of a Larix gmelinii stand near Tura, central Siberia. Tree Physiol. 1999, 19, 815–822. [Google Scholar] [CrossRef]
- Goldammer, J.G.; Furyaev, V.V. Fire in Ecosystems of Boreal Eurasia: Ecological Impacts and Links to the Global System; Springer: Dordrecht, The Netherlands, 1996; pp. 1–20. [Google Scholar]
- Zhang, W.; Yang, K.; Lyu, Z.; Zhu, J. Microbial groups and their functions control the decomposition of coniferous litter: A comparison with broadleaved tree litters. Soil Biol. Biochem. 2019, 133, 196–207. [Google Scholar] [CrossRef]
- Ludwig, S.M.; Alexander, H.D.; Kielland, K.; Mann, P.J.; Natali, S.M.; Ruess, R.W. Fire severity effects on soil carbon and nutrients and microbial processes in a Siberian larch forest. Glob. Chang. Biol. 2018, 24, 5841–5852. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Hu, H.; Li, F.; Zhao, B.; Wu, S.; Zhu, G.; Sun, L. Long-term effects of post-fire restoration types on nitrogen mineralisation in a Dahurian larch (Larix gmelinii) forest in boreal China. Sci. Total Environ. 2019, 679, 237–247. [Google Scholar] [CrossRef]
- Kong, J.; Yang, J.; Cai, W. Topography controls post-fire changes in soil properties in a Chinese boreal forest. Sci. Total Environ. 2019, 651, 2662–2670. [Google Scholar] [CrossRef]
- Amur Center for Hydrometeorology and Environmental Monitoring. Meteorological Characteristics of Zeya Meteorological Station between 1989 and 2011; Amur Center for Hydrometeorology and Environmental Monitoring: Blagoveschensk, Russia, 2015. [Google Scholar]
- WRB, I.W.G. World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2014; Volume 106, ISBN 978-92-5-108369-7. [Google Scholar]
- Procopchuk, V.F.; Bryanin, S.V. Ecological Stability of Brown Raw-humus Taiga Soils in Relation to Anthropogenic Influences in Northern Part of Amur Region. Eurasian J. For. Res. 2007, 10, 85–88. [Google Scholar]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Gartner, T.B.; Cardon, Z.G. Decomposition dynamics in mixed-species leaf litter. Oikos 2004, 104, 230–246. [Google Scholar] [CrossRef]
- Pumpanen, J.; Longdoz, B.L.; Kutsch, W. Field measurements of soil respiration: Principles and constraints, potentials and limitations of different methods. In Soil Carbon Dynamics; Kutsch, W.L., Bahn, M., Heinemeyer, A., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 16–33. ISBN 9780511711794. [Google Scholar]
- Pandey, K.K. A study of chemical structure of soft and hardwood and wood polymers by FTIR spectroscopy. J. Appl. Polym. Sci. 1999, 71, 1969–1975. [Google Scholar] [CrossRef]
- Ouatmane, A.; Provenzano, M.R.; Hafidi, M.; Senesi, N. Compost Maturity Assessment Using Calorimetry, Spectroscopy and Chemical Analysis. Compost Sci. Util. 2000, 8, 124–134. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017.
- Zhang, W.; Gao, D.; Chen, Z.; Li, H.; Deng, J.; Qiao, W.; Han, X.; Yang, G.; Feng, Y.; Huang, J. Substrate quality and soil environmental conditions predict litter decomposition and drive soil nutrient dynamics following afforestation on the Loess Plateau of China. Geoderma 2018, 325, 152–161. [Google Scholar] [CrossRef]
- Prokushkin, S.G.; Prokushkin, A.S.; Sorokin, N.D. The intensity of phytodetrite decomposition in larch forest of the permafrost zone in Central Siberia. Biol. Bull. 2014, 41, 89–97. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, Q.; Sun, X.; Chen, D.; Insam, H.; Koide, R.T.; Zhang, S. Effects of mixed-species litter on bacterial and fungal lignocellulose degradation functions during litter decomposition. Soil Biol. Biochem. 2020, 141, 107690. [Google Scholar] [CrossRef]
- Johnsen, A.R.; Jacobsen, O.S. A quick and sensitive method for the quantification of peroxidase activity of organic surface soil from forests. Soil Biol. Biochem. 2008, 40, 814–821. [Google Scholar] [CrossRef]
- Dove, N.C.; Hart, S.C. Fire Reduces Fungal Species Richness and In Situ Mycorrhizal Colonization: A Meta-Analysis. Fire Ecol. 2017, 13, 37–65. [Google Scholar] [CrossRef]
- Heim, A.; Frey, B. Early stage litter decomposition rates for Swiss forests. Biogeochemistry 2004, 70, 299–313. [Google Scholar] [CrossRef]
- Archibald, F.; Roy, B. Production of manganic chelates by laccase from the lignin-degrading fungus Trametes (Coriolus) versicolor. Appl. Environ. Microbiol. 1992, 58, 1496–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgensen, J.R.; Wells, C.G.; Metz, L.J. Nutrient Changes in Decomposing Loblolly Pine Forest Floor. Soil Sci. Soc. Am. J. 1980, 44, 1307–1314. [Google Scholar] [CrossRef]
- Horodecki, P.; Jagodziński, A.M. Site Type Effect on Litter Decomposition Rates: A Three-Year Comparison of Decomposition Process between Spoil Heap and Forest Sites. Forests 2019, 10, 353. [Google Scholar] [CrossRef] [Green Version]
- Petrillo, M.; Cherubini, P.; Sartori, G.; Abiven, S.; Ascher, J.; Bertoldi, D.; Camin, F.; Barbero, A.; Larcher, R.; Egli, M. Decomposition of norway spruce and european larch coarse woody debris (CWD) in relation to different elevation and exposure in an alpine setting. IForest 2016, 9, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Strukelj, M.; Brais, S.; Quideau, S.A.; Angers, V.A.; Kebli, H.; Drapeau, P.; Oh, S.W. Chemical transformations in downed logs and snags of mixed boreal species during decomposition. Can. J. For. Res. 2013, 43, 785–798. [Google Scholar] [CrossRef]
- Virzo De Santo, A.; De Marco, A.; Fierro, A.; Berg, B.; Rutigliano, F.A. Factors regulating litter mass loss and lignin degradation in late decomposition stages. Plant Soil 2009, 318, 217–228. [Google Scholar] [CrossRef]
- Berg, B.; Ekbohm, G. Nitrogen Immobilization in Decomposing Needle Litter at Variable Carbon: Nitrogen Ratios. Ecology 1983, 64, 63–67. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Control | Burned | ||
|---|---|---|---|---|
| Larix gmelinii (Rupr.) | ||||
| Stem number ha−1 | 436 | 124 | ||
| Basal area m2 ha−1 | 23.1 | 7.9 | ||
| Mean D1.3, cm | 26 | 28 | ||
| Mean height, m | 19 | 19 | ||
| Betula platyphylla Sukacz. | ||||
| Stem number ha−1 | 484 | 64 | ||
| Basal area m2 ha−1 | 21.9 | 1.1 | ||
| Mean D1.3, cm | 24 | 26 | ||
| Mean height, m | 21 | 18 | ||
| Soil Characteristics 0–10 cm | ||||
| Total litter stock: | g m−2 | 251 ± 20.4 | 94.4 ± 11.2 | |
| Water pH | 4.8 ± 0.1 | 5.4 ± 0.1 | ||
| Total organic C, % | 20.1 ± 2.0 | 21.1 ± 2.9 | ||
| Total N, % | 0.77 ± 0.08 | 0.86 ± 0.11 | ||
| Dissolved organic C, mg L−1 | 43.9 ± 4.35 | 24.1 ± 2.04 | ||
| Dissolved organic N, mg L−1 | 1.11 ± 0.17 | 0.77 ± 0.09 | ||
| Average counts of microorganisms, CFU 106g−1dry litter | ||||
| Micromycetes | 2.1 | 0.8 | ||
| Ammonifying bacteria | 4.2 | 1.2 | ||
| Oligotrophic bacteria | 6.4 | 2.0 | ||
| Actinomycetes | 0.2 | 1.4 | ||
| Period (days) | Control | Burned | ||||
|---|---|---|---|---|---|---|
| Mass Loss (%) | k (year−1) | r2 | Mass Loss (%) | k (year−1) | r2 | |
| 140 | 23.4 ± 2.02 | 0.714 | 0.92 | 21.9 ± 1.49 | 0.658 | 0.96 |
| 850 | 67.5 ± 5.18 | 0.435 | 0.96 | 44.2 ± 3.83 | 0.219 | 0.92 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryanin, S.; Kondratova, A.; Abramova, E. Litter Decomposition and Nutrient Dynamics in Fire-Affected Larch Forests in the Russian Far East. Forests 2020, 11, 882. https://doi.org/10.3390/f11080882
Bryanin S, Kondratova A, Abramova E. Litter Decomposition and Nutrient Dynamics in Fire-Affected Larch Forests in the Russian Far East. Forests. 2020; 11(8):882. https://doi.org/10.3390/f11080882
Chicago/Turabian StyleBryanin, Semyon, Anjelica Kondratova, and Evgeniya Abramova. 2020. "Litter Decomposition and Nutrient Dynamics in Fire-Affected Larch Forests in the Russian Far East" Forests 11, no. 8: 882. https://doi.org/10.3390/f11080882