Species Ecological Envelopes under Climate Change Scenarios: A Case Study for the Main Two Wood-Production Forest Species in Portugal

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
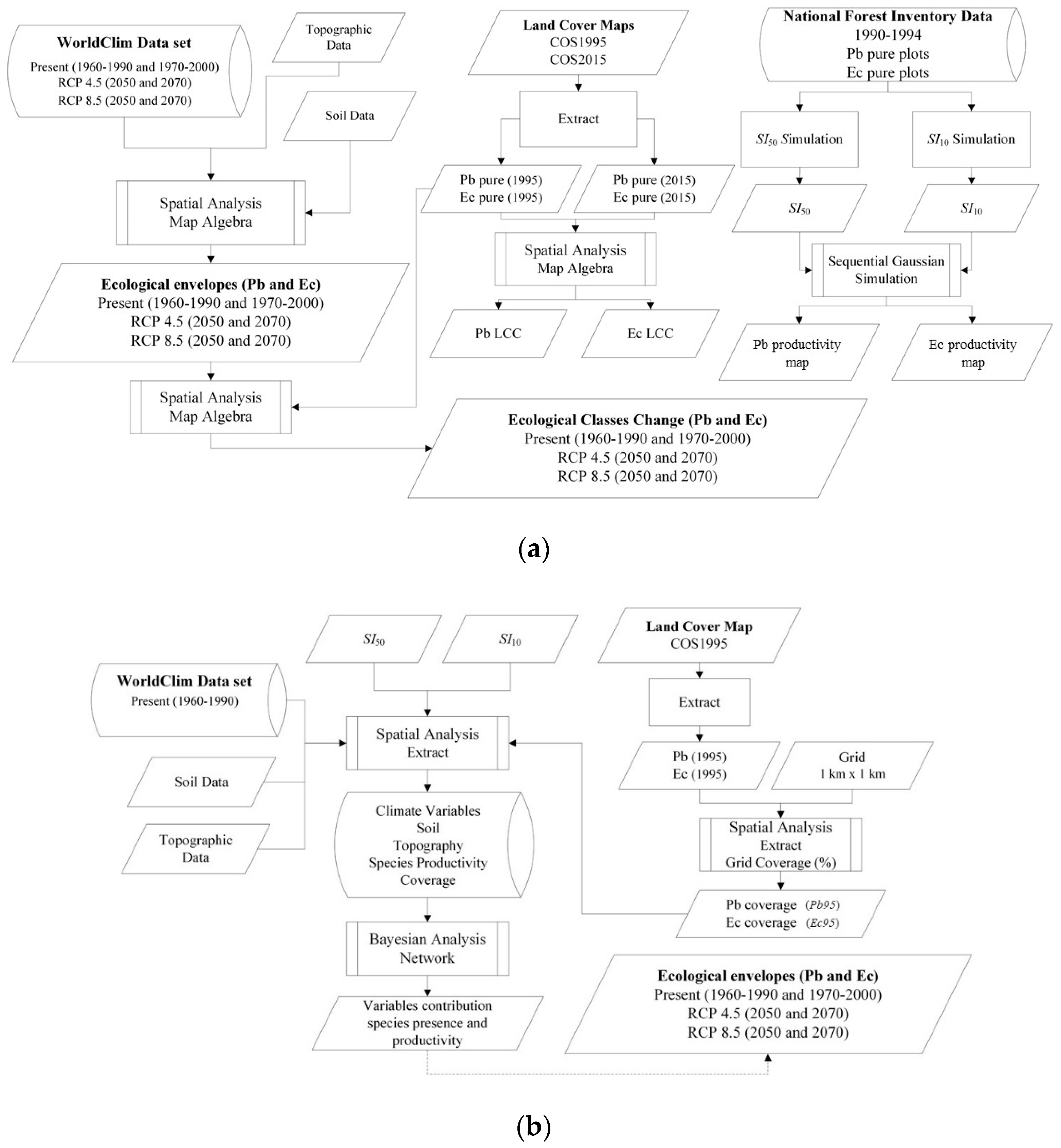
2. Materials and Methods
2.1. Data
2.1.1. Environmental Data—Climate, Topography, and Soil
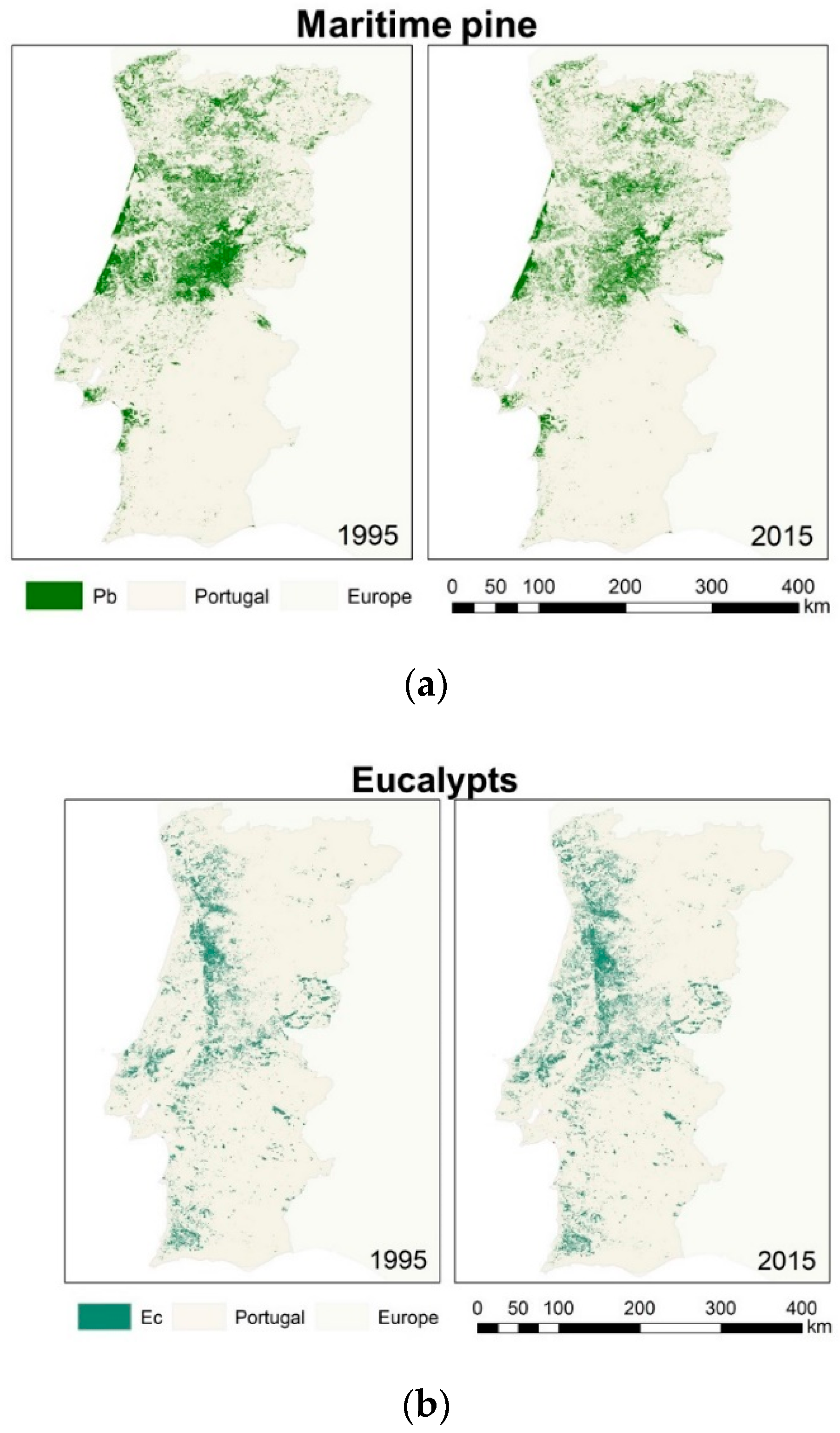
2.1.2. Species Forests Cover (1995 and 2015)
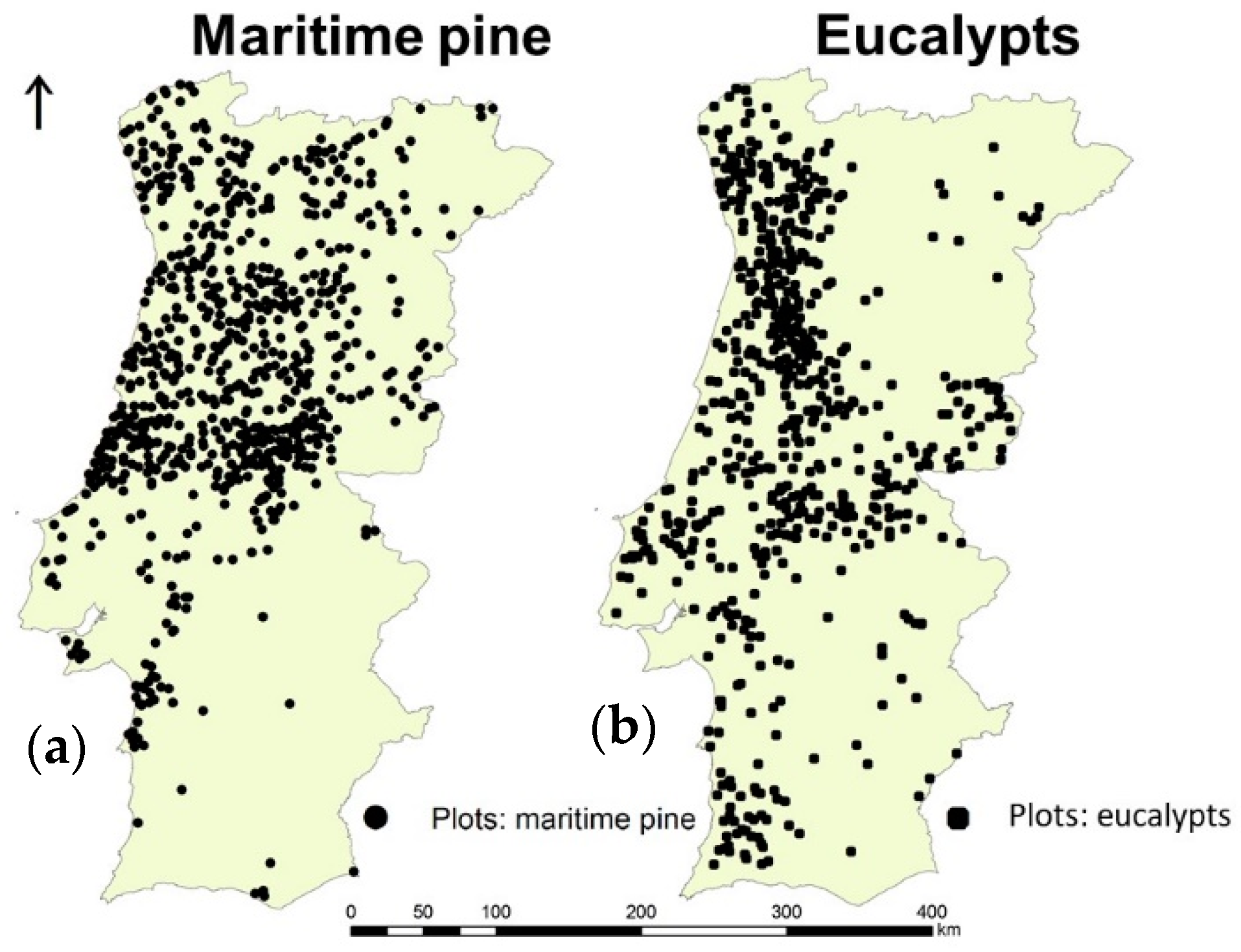
2.1.3. Species Forest Inventory (1990–1994)
2.2. Methods
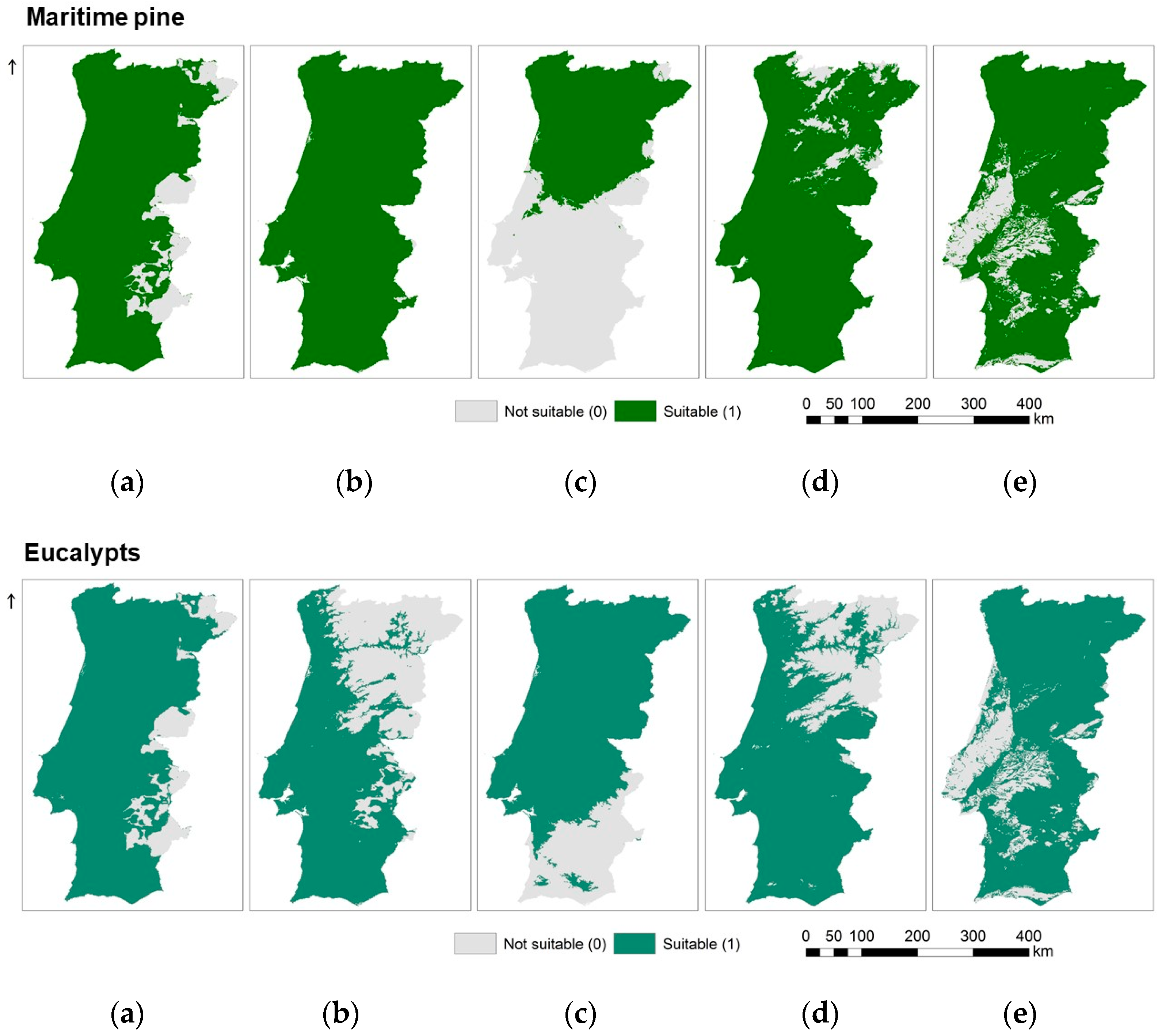
2.2.1. Species Ecological Envelopes Maps
2.2.2. Species Forests Cover Distributions (1995 and 2015)
2.2.3. Species Productivity Maps
2.2.4. Variables Influence Analysis in Species Distribution and Productivity
3. Results
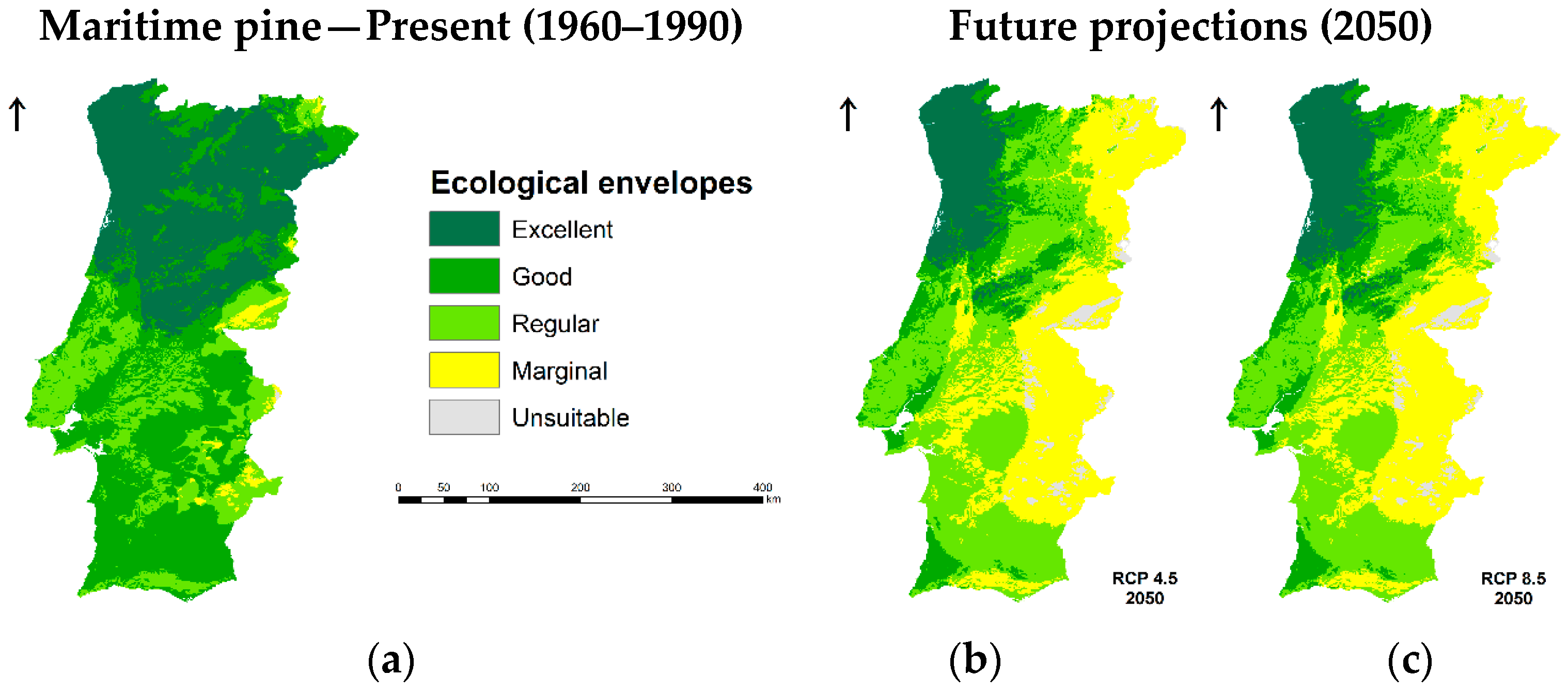
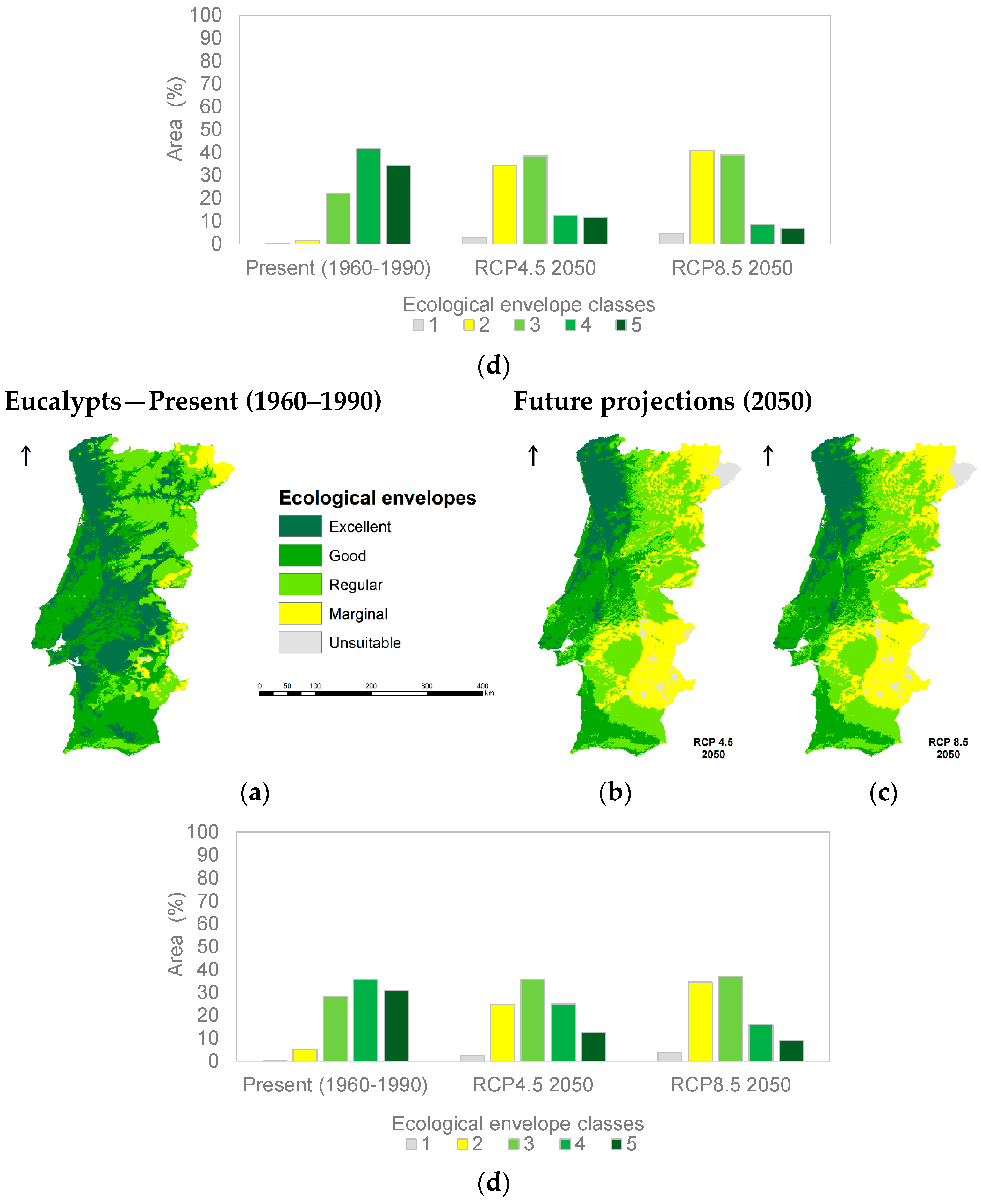
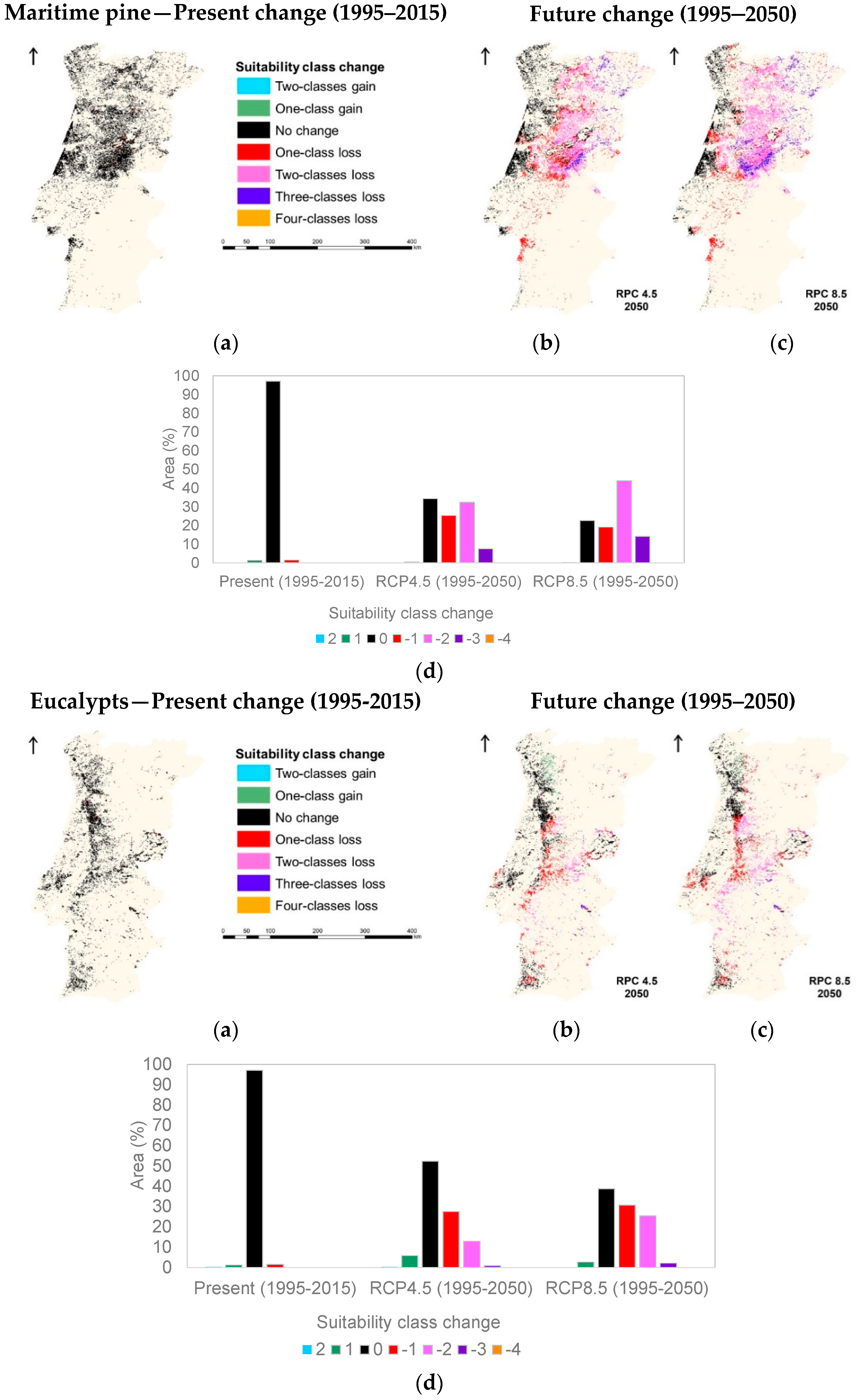
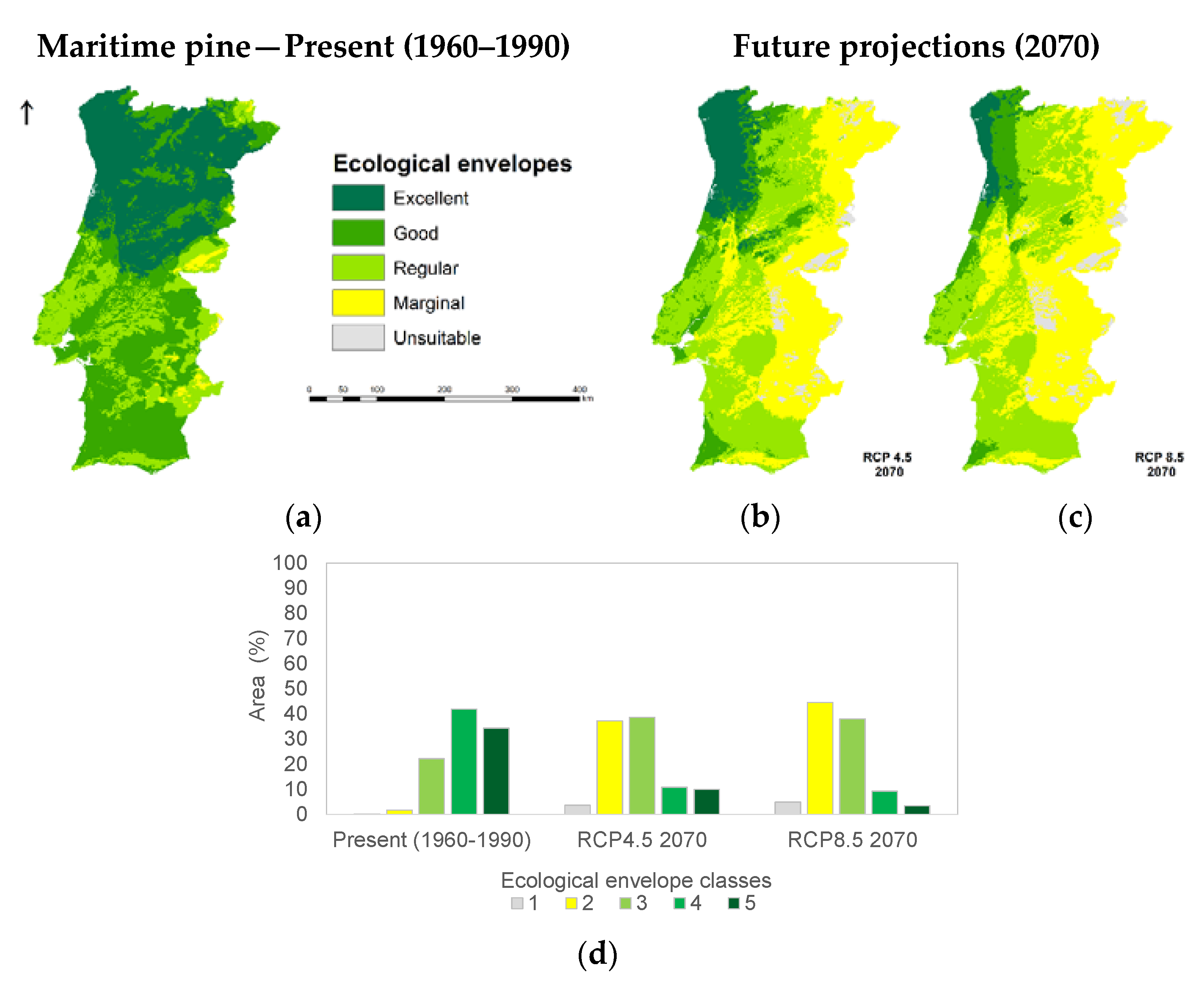
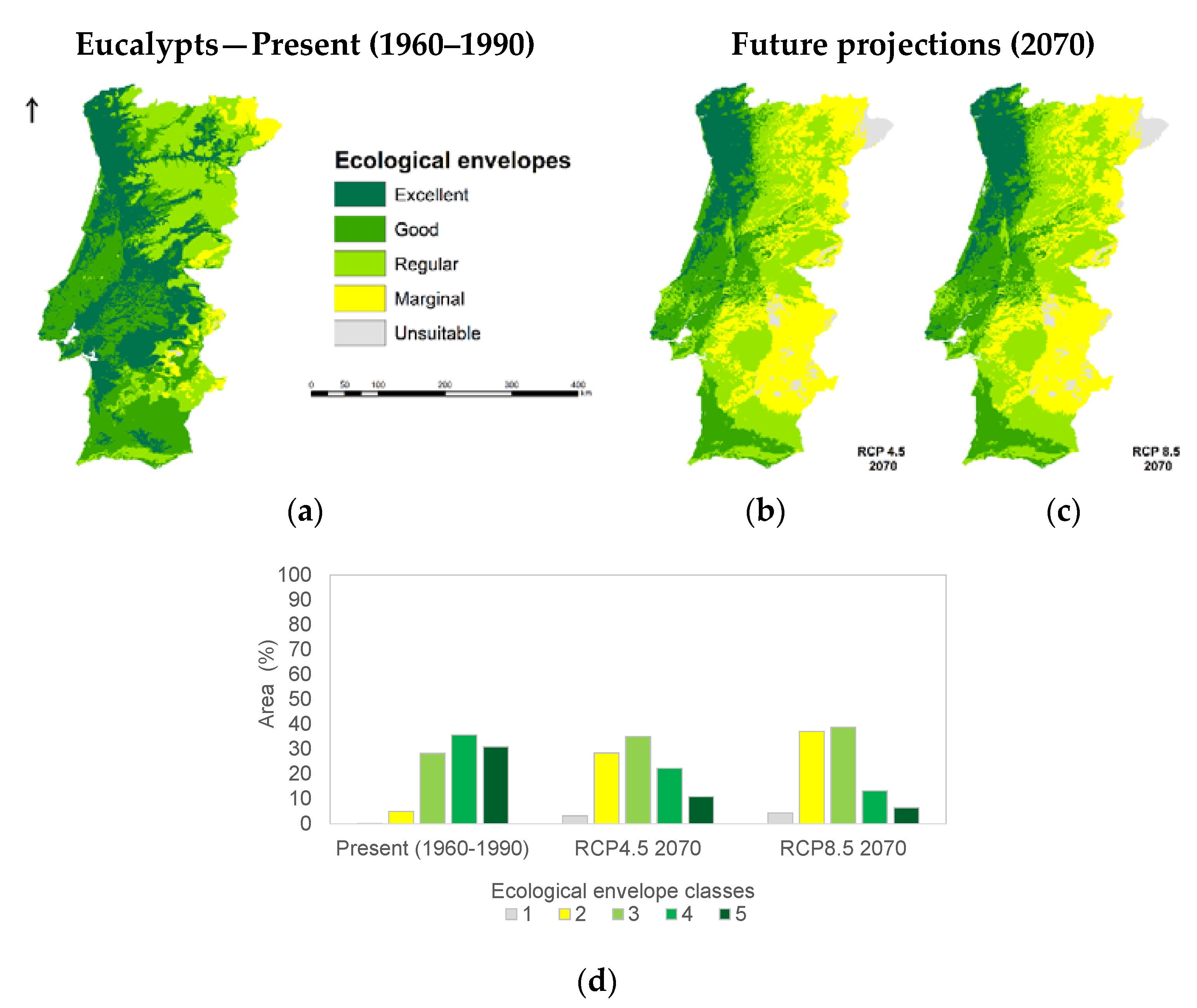
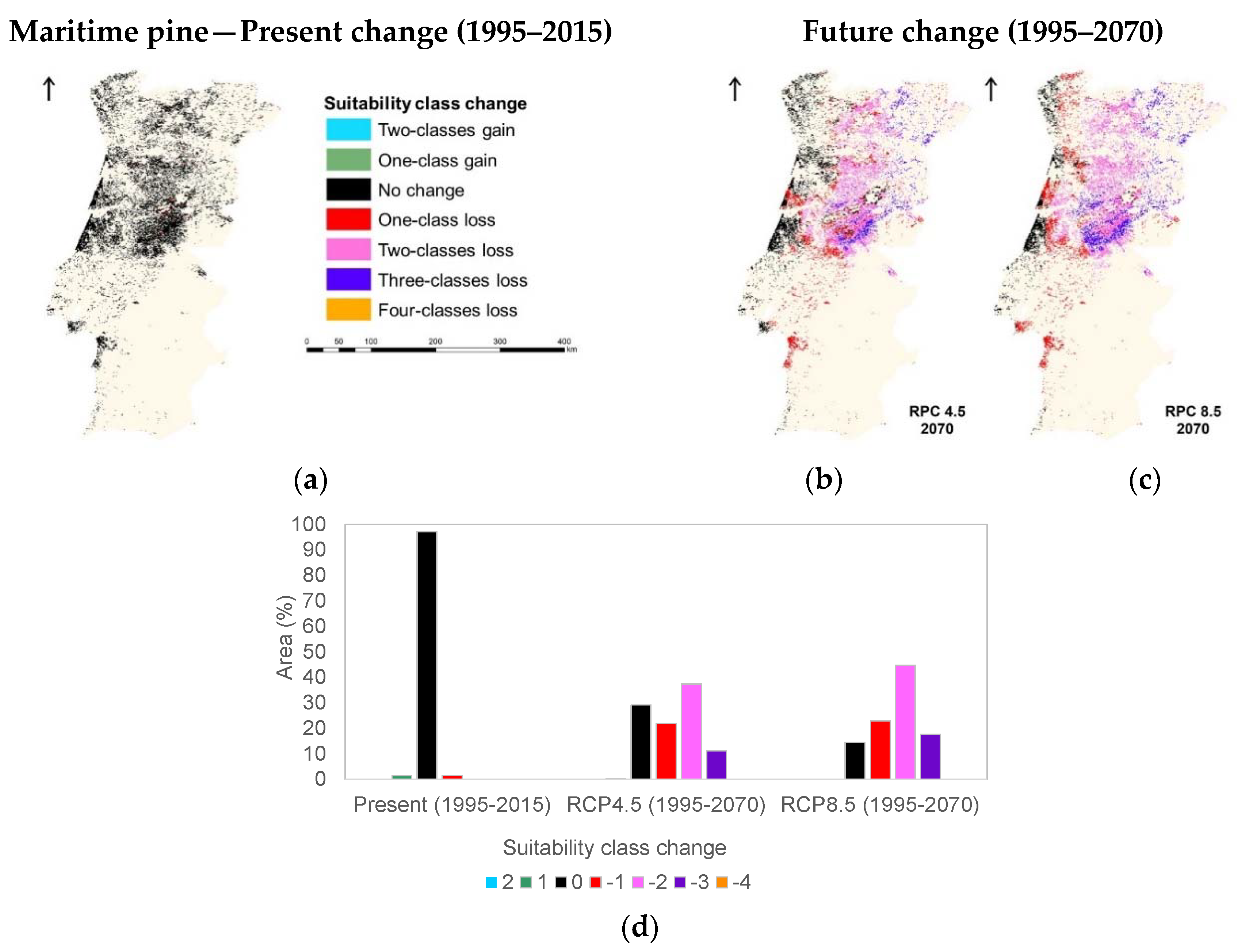
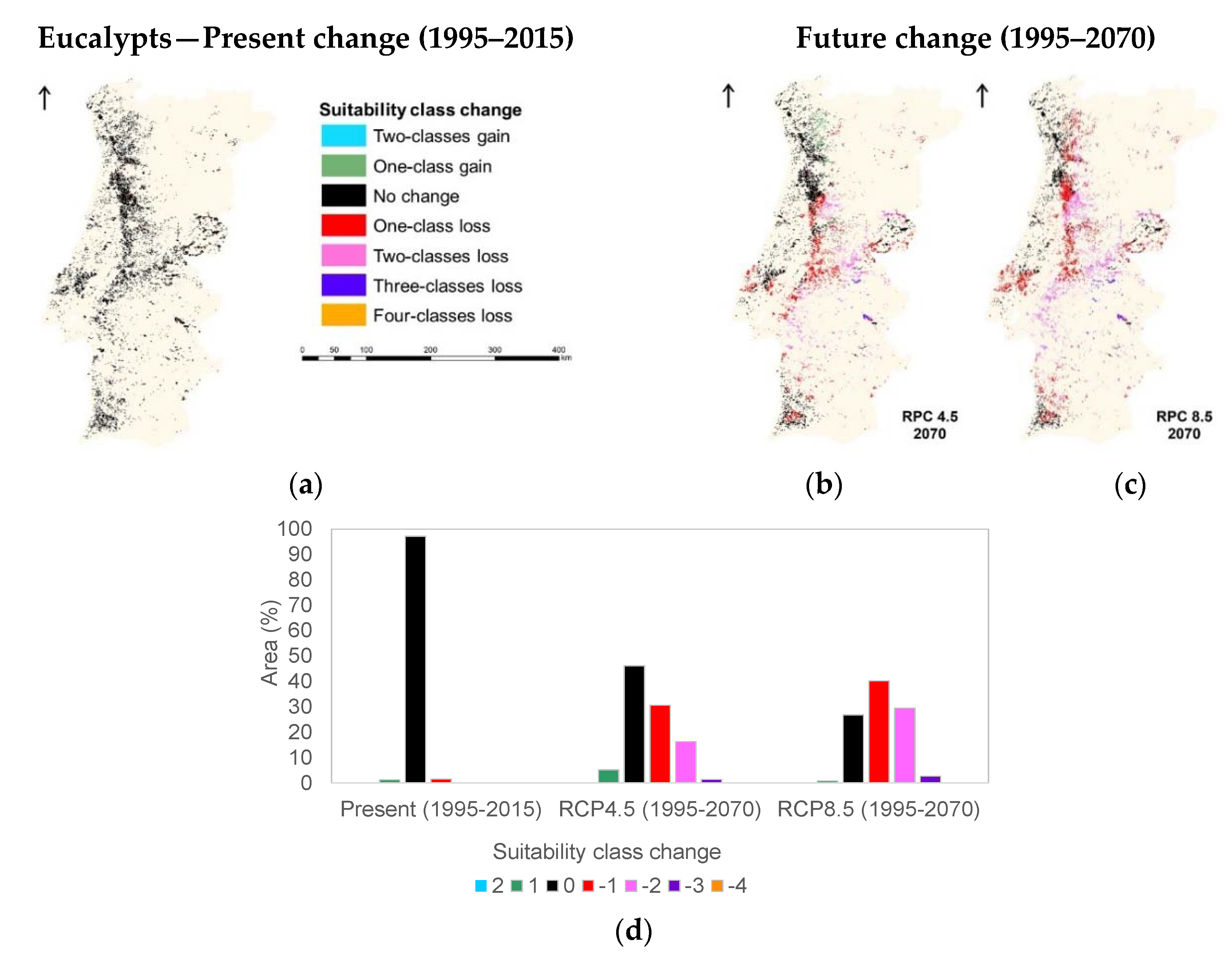
3.1. Species Ecological Envelopes under the Climate Scenarios
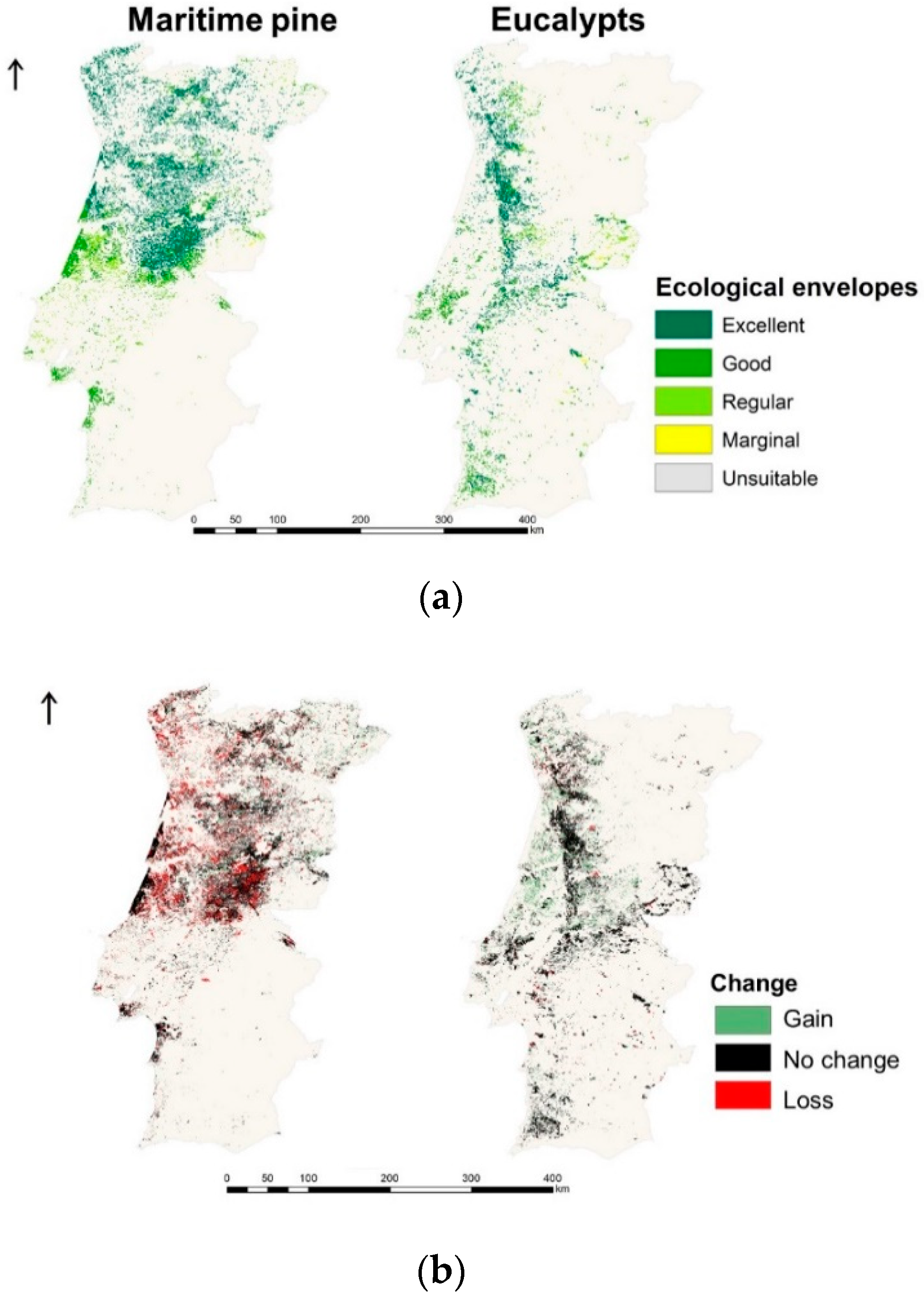
3.2. Species Forests Cover Distributions Maps
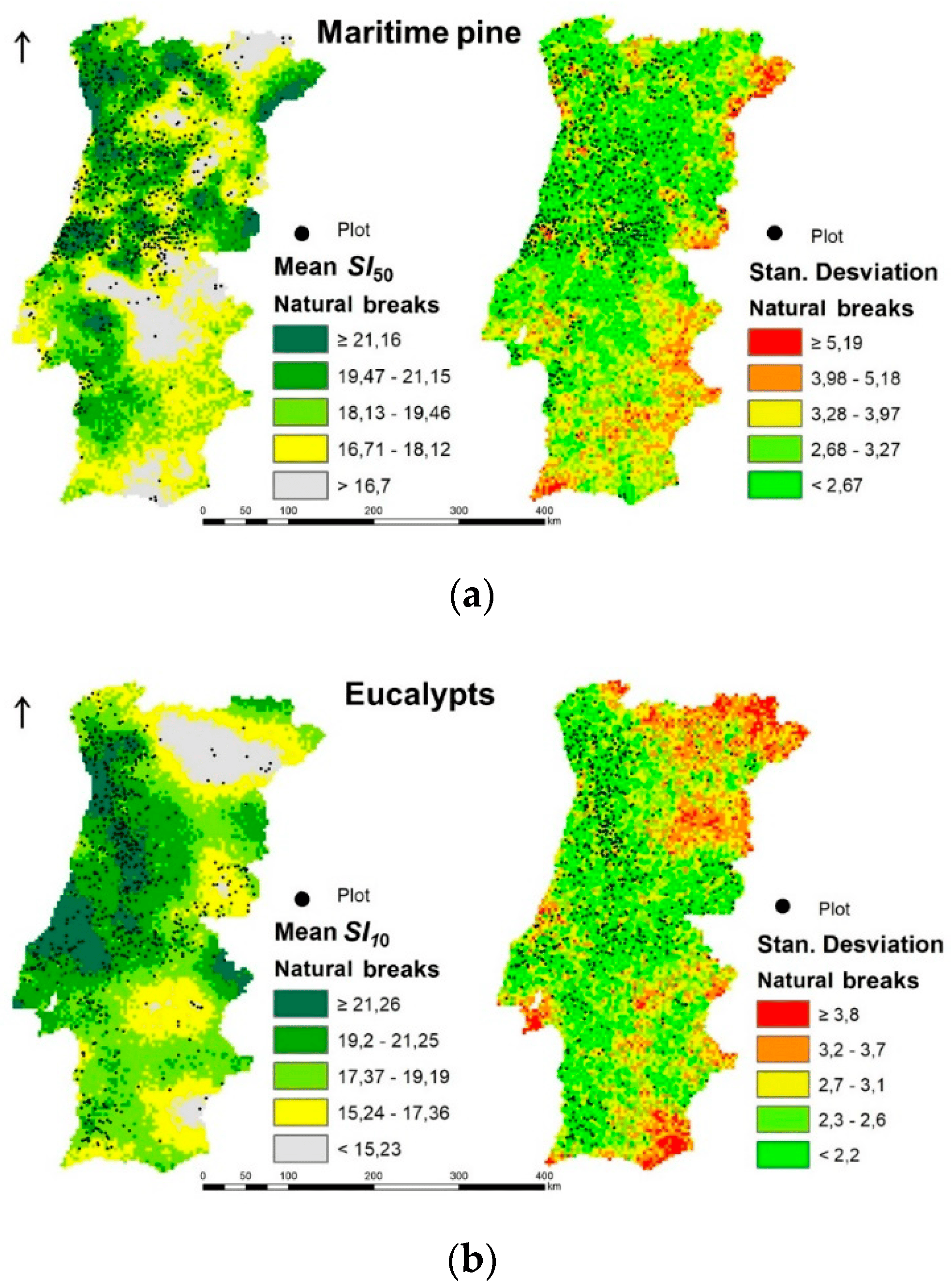
3.3. Current Climate Species Productivities Maps
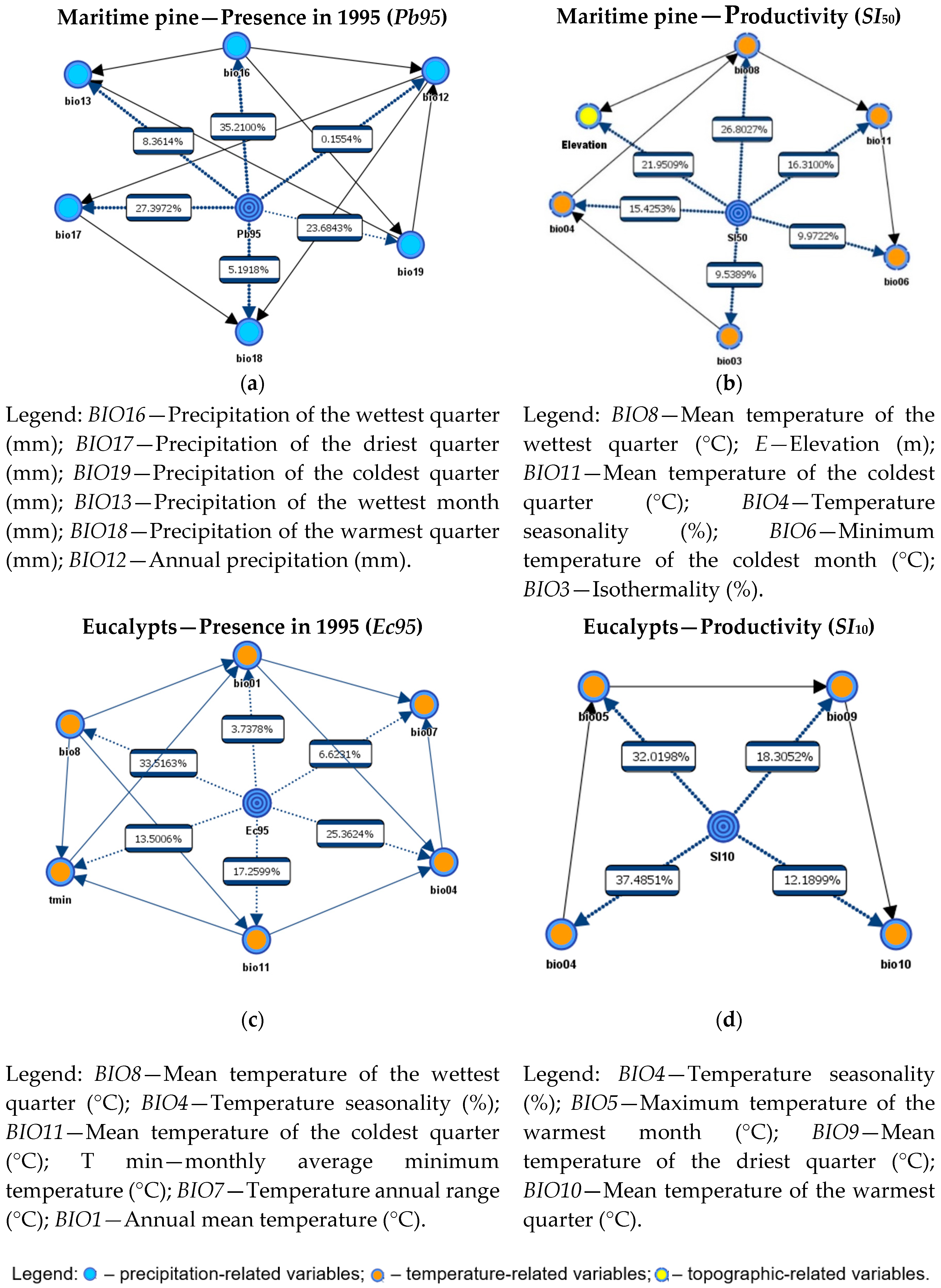
3.4. Variables Influence Analysis in Current Species Distribution and Productivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Units | Min. | Max. | Mean | Std. Dev. | Min. | Max. | Mean | Std. Dev. | Min. | Max. | Mean | Std. Dev. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Presence—Pb95 and Ec95 (n = 88,496) | Maritime pine—SI50 (n = 740) | Eucalypts—SI10 (n = 614) | |||||||||||
| Pb95 | % | 0.0 | 100.0 | 6.6 | 16.5 | ||||||||
| Ec95 | % | 0.0 | 100.0 | 9.6 | 20.0 | ||||||||
| SI50 | m | 5.8 | 48.6 | 18.4 | 4.4 | ||||||||
| SI10 | m | 4.7 | 37.4 | 19.1 | 4.0 | ||||||||
| T max | °C | 9.9 | 22.8 | 19.7 | 1.9 | 4.5 | 13.1 | 9.9 | 1.5 | 16.1 | 22.2 | 19.7 | 1.2 |
| T min | °C | 2.7 | 13.2 | 10.2 | 1.9 | 13.0 | 21.9 | 19.0 | 1.4 | 7.4 | 12.8 | 10.5 | 1.2 |
| BIO1 | °C | 6.3 | 17.6 | 15.0 | 1.8 | 9.5 | 17.5 | 14.5 | 1.4 | 11.8 | 17.1 | 15.1 | 1.1 |
| BIO2 | °C | 5.2 | 11.7 | 9.5 | 1.0 | 5.8 | 11.0 | 9.1 | 0.9 | 6.0 | 11.2 | 9.1 | 0.9 |
| BIO3 | % | 31.0 | 51.0 | 40.0 | 2.4 | 31.0 | 46.0 | 40.4 | 2.7 | 35.0 | 45.0 | 41.1 | 2.2 |
| BIO4 | % | 26.0 | 61.2 | 48.6 | 7.2 | 29.8 | 60.6 | 44.7 | 0.7 | 29.5 | 60.5 | 46.2 | 0.7 |
| BIO5 | °C | 19.7 | 34.0 | 28.6 | 2.5 | 22.1 | 31.9 | 27.3 | 2.1 | 22.4 | 32.7 | 27.6 | 2.4 |
| T max Aug | °C | 19.7 | 33.7 | 28.6 | 2.5 | 20.9 | 28.9 | 24.8 | 1.4 | 20.8 | 29.9 | 25.5 | 1.7 |
| BIO6 | °C | −2.8 | 9.2 | 5.0 | 2.3 | −1.1 | 8.3 | 4.8 | 1.9 | 1.4 | 8.4 | 5.6 | 1.5 |
| T min Jan | °C | −4.6 | 10.0 | 2.8 | 2.0 | −2.6 | 8.4 | 3.0 | 2.0 | 0.3 | 7.7 | 3.6 | 1.4 |
| BIO7 | °C | 12.2 | 29.3 | 23.6 | 2.9 | 14.1 | 27.9 | 22.5 | 2.9 | 14.4 | 27.8 | 22.0 | 2.7 |
| BIO8 | °C | 0.5 | 13.7 | 9.5 | 2.5 | 3.4 | 13.1 | 9.1 | 2.1 | 5.4 | 13.1 | 10.1 | 1.5 |
| BIO9 | °C | 13.6 | 24.9 | 21.2 | 1.7 | 16.8 | 23.8 | 20.4 | 1.3 | 17.4 | 24.4 | 20.8 | 1.5 |
| BIO10 | °C | 13.6 | 25.0 | 21.4 | 1.8 | 16.8 | 23.8 | 20.6 | 1.4 | 17.6 | 24.5 | 21.0 | 1.6 |
| BIO11 | °C | 0.5 | 13.0 | 9.0 | 2.3 | 2.8 | 12.3 | 8.8 | 1.9 | 5.4 | 12.2 | 9.6 | 1.4 |
| BIO12 | mm | 463.0 | 1792.0 | 840.7 | 269.9 | 483.0 | 1593.0 | 1015.2 | 211.7 | 503.0 | 1542.0 | 937.1 | 246.8 |
| BIO13 | mm | 64.0 | 272.0 | 121.5 | 38.5 | 76.0 | 248.0 | 146.7 | 30.0 | 75.0 | 240.0 | 134.0 | 32.4 |
| BIO14 | mm | 0.0 | 37.0 | 8.0 | 5.8 | 1.0 | 30.0 | 10.6 | 4.4 | 1.0 | 28.0 | 9.2 | 5.1 |
| BIO15 | % | 39.0 | 72.0 | 56.4 | 5.4 | 43.0 | 71.0 | 54.8 | 3.5 | 48.0 | 70.0 | 55.7 | 4.5 |
| BIO16 | mm | 180.0 | 719.0 | 345.4 | 102.6 | 220.0 | 657.0 | 413.0 | 79.9 | 220.0 | 619.0 | 383.5 | 91.4 |
| BIO17 | mm | 13.0 | 157.0 | 51.7 | 25.5 | 14.0 | 132.0 | 65.2 | 19.5 | 16.0 | 123.0 | 58.1 | 23.0 |
| BIO18 | mm | 15.0 | 161.0 | 54.8 | 27.6 | 17.0 | 143.0 | 70.2 | 22.6 | 18.0 | 137.0 | 64.1 | 27.8 |
| BIO19 | mm | 168.0 | 719.0 | 341.0 | 104.3 | 202.0 | 657.0 | 409.5 | 81.3 | 207.0 | 619.0 | 378.3 | 91.9 |
| E | m | 0.0 | 1921.0 | 321.0 | 262.6 | 0.0 | 1275.0 | 329.1 | 248.7 | 8.0 | 752.0 | 231.2 | 155.3 |
| S | % | 0.0 | 25.4 | 2.8 | 3.0 | 0.1 | 20.2 | 3.6 | 3.4 | 0.0 | 17.4 | 2.9 | 2.5 |
| A | ° | 0.0 | 360.0 | 188.7 | 103.4 | 0.0 | 360.0 | 197.7 | 97.9 | 0.0 | 359.7 | 198.4 | 99.7 |
| WRBFU | 2.0 | 130.0 | 86.2 | 33.8 | 27.0 | 124.0 | 83.8 | 38.1 | 27.0 | 127.0 | 85.3 | 36.1 | |




| Variable | Pb95 | Ec95 | SI50 | SI10 | ||||
|---|---|---|---|---|---|---|---|---|
| RMI (%) | RS (%) | RMI (%) | RS (%) | RMI (%) | RS (%) | RMI (%) | RS (%) | |
| T max | 5.95 | 57.61 | 3.91 | 62.16 | 0.95 | 44.09 | 2.14 | 60.02 |
| T min | 6.60 | 63.83 | 5.54 | 88.01 | 1.43 | 66.24 | 1.54 | 43.26 |
| BIO1 | 6.02 | 58.24 | 4.60 | 73.06 | 1.44 | 66.36 | 2.12 | 59.30 |
| BIO2 | 3.75 | 36.27 | 3.73 | 59.31 | 0.81 | 37.15 | 0.85 | 23.94 |
| BIO3 | 1.94 | 19.18 | 3.86 | 61.38 | 1.63 | 75.38 | 1.67 | 46.75 |
| BIO4 | 2.97 | 28.74 | 5.32 | 84.45 | 1.62 | 74.88 | 3.07 | 85.79 |
| BIO5 | 3.42 | 33.15 | 2.60 | 41.35 | 1.09 | 50.38 | 3.58 | 100.00 |
| BIO6 | 2.54 | 24.58 | 0.53 | 8.41 | 1.77 | 81.97 | 0.63 | 17.61 |
| BIO7 | 3.19 | 30.88 | 5.38 | 85.45 | 1.36 | 62.99 | 2.27 | 63.63 |
| BIO8 | 5.88 | 59.88 | 6.30 | 100.00 | 2.16 | 100.00 | 0.58 | 16.22 |
| BIO9 | 3.80 | 36.79 | 2.32 | 36.83 | 0.72 | 41.50 | 2.98 | 83.37 |
| BIO10 | 4.41 | 42.66 | 2.28 | 36.25 | 0.71 | 33.16 | 3.02 | 84.50 |
| BIO11 | 5.57 | 53.89 | 5.50 | 87.35 | 1.96 | 90.52 | 1.01 | 28.03 |
| BIO12 | 10.27 | 99.40 | 2.64 | 41.99 | 0.59 | 27.52 | 1.59 | 44.62 |
| BIO13 | 9.91 | 95.89 | 4.15 | 65.99 | 1.14 | 52.71 | 1.35 | 37.92 |
| BIO14 | 5.91 | 57.22 | 0.97 | 15.46 | 0.33 | 15.45 | 1.26 | 35.36 |
| BIO15 | 5.78 | 55.54 | 2.40 | 38.24 | 0.96 | 44.76 | 1.34 | 37.87 |
| BIO16 | 10.21 | 98.73 | 3.08 | 48.99 | 0.55 | 25.69 | 1.65 | 46.29 |
| BIO17 | 8.32 | 80.45 | 1.77 | 28.18 | 0.72 | 32.90 | 1.02 | 28.51 |
| BIO18 | 8.80 | 85.12 | 1.19 | 18.95 | 0.81 | 37.45 | 1.09 | 30.70 |
| BIO19 | 10.34 | 100.00 | 3.12 | 49.57 | 0.87 | 26.33 | 1.60 | 44.44 |
| Elevation | 1.99 | 19.27 | 2.91 | 46.31 | 1.95 | 90.34 | 0.56 | 15.64 |
| Slope | 0.92 | 8.99 | 0.40 | 6.36 | 1.25 | 58.07 | 0.25 | 7.23 |
| Aspect | 0.11 | 1.12 | 0.10 | 1.60 | 0.34 | 15.88 | 0.46 | 13.11 |
| WRBFU | 1.02 | 9.95 | 0.60 | 9.64 | 1.15 | 53.45 | 1.55 | 43.48 |
References
- Miranda, P.M.A.; Coelho, F.E.S.; Tomé, A.R.; Valente, M.A.; Carvalho, A.; Pires, C.; Pires, H.O.; Pires, V.C.; Ramalho, C. 20th century Portuguese climate and climate scenarios. In Climate Change in Portugal: Scenarios, Impacts and Adaptation Measures (SIAM Project); Santos, F.D., Forbes, K., Moita, R., Eds.; Gradiva: Lisbon, Portugal, 2002; pp. 23–83. [Google Scholar]
- DR. Estratégia Nacional para as Florestas, Resolução de Conselho de Ministros nº 114/2006, Diário da República, I Série—nº 179 de 15 de Setembro. 2006. Available online: https://dre.pt/application/file/539887 (accessed on 9 March 2018).
- DR. Estratégia Nacional para as Florestas, Resolução de Conselho de Ministros nº 6-B/2015, Diário da República, I Série—nº 24 de 4 de Fevereiro. 2015. Available online: https://dre.pt/application/file/66432612 (accessed on 9 March 2018).
- Pereira, J.S.; Correia, A.V.; Correia, C.V.; Ferreira, M.T.; Onofre, N.; Freitas, H.; Godinho, F. Florestas e biodiversidade. In Alterações Climáticas em Portugal. Cenários, Impactos e Medidas de Adaptação (Projecto SIAM II); Santos, F.D., Miranda, P.M.A., Eds.; Gradiva: Lisbon, Portugal, 2006; pp. 301–344. [Google Scholar]
- Leite, A.; Martins, L. Forestry resources. In Forests of Portugal; Direcção Geral das Florestas (DGF): Lisbon, Portugal, 2000; pp. 139–143. [Google Scholar]
- Jones, N.; de Graaff, J.; Rodrigo, I.; Duarte, F. Historical review of land use changes in Portugal (before and after EU integration in 1986) and their implications for land degradation and conservation, with a focus on Centro and Alentejo regions. Appl. Geogr. 2011, 31, 1036–1048. [Google Scholar] [CrossRef]
- Teixeira, J.S.; Matos, J. The eucalyptus sector. In Forests of Portugal; Direcção Geral das Florestas (DGF): Lisbon, Portugal, 2000; pp. 203–207. [Google Scholar]
- Alves, A.M.; Pereira, J.S.; Silva, J.M.N. A introdução e expansão do eucalipto em Portugal. In O Eucaliptal em Portugal: Impactes Ambientais e Investigação Científica; Alves, A.M., Pereira, J.S., Silva, J.M.N., Eds.; ISA Press: Lisbon, Portugal, 2007; pp. 13–24. [Google Scholar]
- Catry, F.X.; Moreira, F.; Deus, E.; Silva, J.S.; Águas, A. Assessing the extent and the environmental drivers of Eucalyptus globulus wildling establishment in Portugal: Results from a countrywide survey. Biol. Invasions 2015, 17, 3163–3181. [Google Scholar] [CrossRef]
- Borralho, N.M.G.; Almeida, M.H.; Potts, B.M. O melhoramento do eucalipto em Portugal. In O eucaliptal em Portugal: Impactes Ambientais e Investigação Científica; Alves, A.M., Pereira, J.S., Silva, J.M.N., Eds.; ISA Press: Lisbon, Portugal, 2007; pp. 61–110. [Google Scholar]
- ICNF. 6º Inventário Florestal Nacional–IFN6. Áreas dos usos do solo e das espécies florestais de Portugal continental. 1995 | 2005 | 2010. Resultados Preliminaries; Instituto da Conservação da Natureza e das Florestas: Lisboa, Portugal, 2013; Available online: http://www2.icnf.pt/portal/florestas/ifn/resource/doc/ifn/ifn6-res-prelimv1-1 (accessed on 20 February 2020).
- INE. Estatísticas Agrícolas; Instituto Nacional de Estatística: Lisbon, Portugal, 2011.
- IEFC. Formodels; Institut Européen De La Forêt Cultivée: Bordeaux-Pierrot, France, 2020; Available online: http://www.iefc.net/formodels_database_forest_modeles_liste/ (accessed on 11 August 2020).
- Meason, D.; Mason, W. Evaluating the deployment of alternative species in planted conifer forests as a means of adaptation to climate change—Case studies in New Zealand and Scotland. Ann. For. Sci. 2014, 71. [Google Scholar] [CrossRef] [Green Version]
- Fourcade, Y.; Engler, J.O.; Rodder, D.; Secondi, J. Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 2019, 9, e97122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrio-Anta, M.; Castedo-Dorado, F.; Cámara-Obregón, A.; López-Sánchez, C. Predicting current and future suitable habitat and productivity for Atlantic populations of maritime pine (Pinus pinaster Aiton) in Spain. Ann. For. Sci. 2020, 77, 41. [Google Scholar] [CrossRef]
- Bede-Fazekas, A.; Levente Horváth, K.M. Impact of climate change on the potential distribution of Mediterranean pines. Q. J. Hung. Meteorol. Serv. 2014, 118, 41–52. [Google Scholar]
- Deus, E.; Silva, J.S.; Castro-Díez, P.; Lomba, A.; Ortiz, M.L.; Vicente, J. Current and future conflicts between eucalypt plantations and high biodiversity areas in the Iberian Peninsula. J. Nat. Conserv. 2018, 45, 107–117. [Google Scholar] [CrossRef]
- Costa, R.; Fraga, H.; Fernandes, P.M.; Santos, J.A. Implications of future bioclimatic shifts on Portuguese forests. Reg. Environ. Change 2017, 17, 117–127. [Google Scholar] [CrossRef]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Krey, T.K.V.; Lamarque, J.; Masui, T.; et al. The representative concentration pathways: An overview. Clim. Change 2011, 109, 5. [Google Scholar] [CrossRef]
- Ribeiro, M.M.; Roque, N.; Ribeiro, S.; Gavinhos, C.; Castanheira, I.; Quinta-Nova, L.; Albuquerque, T.; Gerassis, S. Bioclimatic modeling in the Last Glacial Maximum, Mid-Holocene and facing future climatic changes in the strawberry tree (Arbutus unedo L.). PLoS ONE 2019, 14, e0210062. [Google Scholar] [CrossRef] [Green Version]
- ICNF. Planos Regionais de Ordenamento Florestal; Instituto da Conservação da Natureza e das Florestas: Lisboa, Portugal, 2019; Available online: http://www2.icnf.pt/portal/florestas/profs/prof-em-vigor (accessed on 20 February 2020).
- Navalho, I.; Alegria, C.; Quinta-Nova, L.; Fernandez, P. Integrated planning for landscape diversity enhancement, fire hazard mitigation and forest production regulation: A case study in central Portugal. Land Policy 2017, 61, 398–412. [Google Scholar] [CrossRef]
- Mestre, S.; Alegria, C.; Teresa, M.; Albuquerque, D.; Goovaerts, P. Developing an index for forest productivity mapping—A case study for maritime pine production regulation in Portugal. Rev. Árvore 2017, 41, e410306. [Google Scholar] [CrossRef]
- Quinta-Nova, L.; Roque, N.; Navalho, I.; Alegria, C.; Albuquerque, T. Using geostatistics and multicriteria spatial analysis to map forest species biogeophysical suitability: A study case for the Centro region of Portugal. In Information and Communication Technologies in Modern Agricultural Development. HAICTA 2017; Salampasis, M., Bournaris, T., Eds.; Springer: Cham, Switzerland, 2019; Volume 953, pp. 64–83. [Google Scholar] [CrossRef]
- DGRF. Plano Regional de Ordenamento Florestal do Pinhal Interior Sul. Documento estratégico; Direção Geral dos Recursos Florestais: Lisbon, Portugal, 2005. [Google Scholar]
- Alves, A.A.M. Técnicas de Produção Florestal; Instituto Nacional de Investigação Científica: Lisbon, Portugal, 1988. [Google Scholar]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The community climate system model version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high-resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. BIOCLIM: The first species distribution modelling package, its early applications and relevance to most current MAXENT studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- STRM. Shuttle Radar Topography Mission 1 Arc-Second Global: SRTM1N22W016V3; U.S. Geological Survey (USGS); National Geospatial-Intelligence Agency (NGA); National Aeronautics and Space Administration (NASA): Sioux Falls, SD, USA, 2017. Available online: http://www2.jpl.nasa.gov/srtm/ (accessed on 9 March 2018).
- Panagos, P. The European soil database. GEO Connex. 2006, 5, 32–33. [Google Scholar]
- Van Liedekerke, M.; Jones, A.; Panagos, P. ESDBv2 Raster Library—A Set of Rasters Derived from the European Soil Database Distribution v2.0 (CD-ROM, EUR 19945 EN); European Commission and the European Soil Bureau Network. 2006. Available online: https://esdac.jrc.ec.europa.eu/content/european-soil-database-v2-raster-library-1kmx1km (accessed on 29 July 2018).
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- DGT. Catálogo de Serviços de Dados Geográficos; Direção Geral do Território: Lisboa, Portugal, 2020. Available online: https://snig.dgterritorio.gov.pt/rndg/srv/por/catalog.search#/search?anysnig=COS&fast=index (accessed on 20 February 2020).
- DGT. Especificações Técnicas da Carta de Uso e Ocupação do Solo de Portugal Continental para 1995, 2007, 2010 e 2015. Relatório Técnico; Direção-Geral do Território: Lisboa, Portugal, 2018. Available online: http://www.dgterritorio.pt/cartografia_e_geodesia/cartografia/cartografia_tematica/cartografia_de_uso_e_ocupacao_do_solo__cos_clc_e_copernicus_/ (accessed on 27 July 2019).
- ICNF. 6º Inventário Florestal Nacional—IFN6. 2015. Relatório Final; Instituto da Conservação da Natureza e das Florestas: Lisboa, Portugal, 2019; Available online: http://www2.icnf.pt/portal/florestas/ifn/resource/doc/ifn/ifn6/IFN6_Relatorio_completo-2019-11-28.pdf (accessed on 20 February 2020).
- ICNF. Inventário Florestal Nacional. IFN4. Dados de base de 1995-98; ICNF: Lisboa, Portugal, 2020; Available online: http://www2.icnf.pt/portal/florestas/ifn/ifn4/dad-base95-98 (accessed on 11 August 2020).
- Oliveira, A. Boas Práticas Florestais para o Pinheiro Bravo. Manual; Centro Pinus: Porto, Portugal, 1999; Available online: https://centropinus.org/files/2018/04/manual01.pdf (accessed on 20 January 2017).
- Alves, A.M.; Pereira, J.S.; Correia, V.A. Silvicultura. A Gestão dos Ecossistemas Florestais; Fundação Calouste Gulbenkian: Lisbon, Portugal, 2012. [Google Scholar]
- Santos, C.; Almeida, J.A. Spatial characterization of maritime pine productivity in Portugal. In Forest Context and Policies in Portugal. World Forests; Reboredo, F., Ed.; Springer: Cham, Switzerland, 2014; Volume 19, pp. 185–217. [Google Scholar] [CrossRef]
- Oliveira, A.C.; Pereira, J.S.; Correia, A.V. A Silvicultura do Pinheiro Bravo. Manual; Centro Pinus: Porto, Portugal, 2000. [Google Scholar]
- Borralho, N.; Silva, M. O Imperativo da Renovação da Floresta de Eucalipto; RAIZ, Instituto de Investigação da Floresta e Papel: Eixo, Portugal, 2006. [Google Scholar]
- Vanclay, J. Modeling Forest Growth and Yield. Applications to Mixed Tropical Forests; CAB International: Wallingford, UK, 1994. [Google Scholar]
- Nunes, L.; Patrício, M.; Tomé, J.; Tomé, M. Modeling dominant height growth of maritime pine in Portugal using GADA methodology with parameters depending on soil and climate variables. Ann. For. Sci. 2011, 68, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Tomé, M.; Oliveira, T.; Soares, P. O modelo Globulus 3.0. Publicações GIMREF—RC2/2006; Centro de Estudos Florestais, Instituto Superior de Agronomia, Universidade Técnica de Lisboa: Lisboa, Portugal, 2006; Available online: https://www.repository.utl.pt/handle/10400.5/1760 (accessed on 9 March 2018).
- Tomé, M.; Ribeiro, F.; Soares, P. O modelo Globulus 2.1. Relatórios Técnico-Científicos do GIMREF, n° 1/2001; Departamento Engenharia Florestal, Instituto Superior de Agronomia: Lisboa, Portugal, 2001; Available online: https://www.repository.utl.pt/bitstream/10400.5/.../REP-DEF-Relatorio-Globulus_2.1.pdf (accessed on 29 July 2018).
- Cieszewski, C.J. GADA derivation of dynamic site equations with polymorphism and variable asymptotes from Richards, Weibull and other Exponential Functions. In Second International Conference on Forest Measurements and Qualitative Methods and Management; Cieszewski, C.J., Strub, M., Eds.; University of Georgia: Athens, GA, USA, 2004; pp. 248–261. [Google Scholar]
- SMN. O Clima de Portugal. Normais Climatológicas do Continente, Açores e Madeira correspondentes a 1931-1980. Fascículo XLIII; Serviço Meteorológico Nacional: Lisbon, Portugal, 1970.
- INMG. O Clima de Portugal. Normais Climatológicas da Região de “Entre Douro e Minho” e “Beira Litoral” correspondentes a 1951–1980. Fascículo XLIX; Instituto Nacional Meteorológico e Geofísica: Lisbon, Portugal, 1990; Volume 1–1ª região. [Google Scholar]
- INMG. O Clima de Portugal. Normais Climatológicas da Região de “Ribatejo e Oeste” Correspondentes a 1951–1980. Fascículo XLIX; Instituto Nacional Meteorológico e Geofísica: Lisbon, Portugal, 1991; Volume 2–2ª região. [Google Scholar]
- INMG. O Clima de Portugal. Normais Climatológicas da Região de “Trás-os-Montes e Alto Douro e Beira Interior” Correspondentes a 1951–1980. Fascículo XLIX; Instituto Nacional Meteorológico e Geofísica: Lisbon, Portugal, 1991; Volume 3–3ª região. [Google Scholar]
- INMG. O Clima de Portugal. Normais Climatológicas da Região de “Alentejo e Algarve” Correspondentes a 1951–1980. Fascículo XLIX; Instituto Nacional Meteorológico e Geofísica: Lisbon, Portugal, 1991; Volume 4–4ª região. [Google Scholar]
- IPMA. O Clima de Portugal. Normais Climatológicas. Fichas Climatológicas 1971–2000; Instituto Português do Mar e da Atmosfera: Lisboa, Portugal, 2020. Available online: https://www.ipma.pt/pt/oclima/normais.clima/1971-2000/normalclimate7100.jsp (accessed on 20 February 2020).
- Journel, A.; Huijbregts, C. Mining Geostatistics; Academic Press: San Diego, CA, USA, 1978. [Google Scholar]
- BioMedware. SpaceStat. BioMedware; Geospatial Research and Sofware: Ann Arbor, MI, USA, 2018; Available online: https://www.biomedware.com/software/spacestat/ (accessed on 11 August 2020).
- Goovaerts, P. Geostatistics for Natural Resources Evaluation; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Albuquerque, M.T.D.; Gerassis, S.; Sierra, C.; Taboada, J.; Martín, J.E.; Antunes, I.M.H.R.; Gallego, J.R. Developing a new Bayesian Risk Index for risk evaluation of soil contamination. Sci. Total Environ. 2017, 603, 167–177. [Google Scholar] [CrossRef]
- Jenks, G.F. The data model concept in statistical mapping. Int. Yearb. Cartogr. 1967, 7, 186–190. [Google Scholar]
- Pascual, M.; Pérez, E.; Giacomello, E. Integrating knowledge on biodiversity and ecosystem services: Mind-mapping and Bayesian Network modelling. Ecosyst. Serv. 2016, 17, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A mathematical theory of communication. Bell. Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Bayesia. BayesiaLab 9; Bayesia S.A.S.: Changé, France, 2020; Available online: https://www.bayesialab.com/ (accessed on 11 August 2020).
- Conrady, S.; Jouffe, L. Bayesian Networks and BayesiaLab—A Practical Introduction for Researchers; Franklin: Bayesia, USA: Laval, France, 2015. [Google Scholar]
- WEKA. WEKA software; Machine Learning Group at the University of Waikato: Waikato, New Zealand, 2020; Available online: https://www.cs.waikato.ac.nz/ml/weka/ (accessed on 11 August 2020).
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Witten, I.H.; Frank, E.; Hall, M.A. Data Mining: Practical Machine Learning Tools and Techniques; Morgan Kaufmann: Burlington, VT, USA, 2011. [Google Scholar]
- Alegria, C.; Pedro, N.; do Carmo Horta, M.; Roque, N.; Fernandez, P. Ecological envelope maps and stand production of eucalyptus plantations and naturally regenerated maritime pine stands in the central inland of Portugal. For. Ecol. Manag. 2019, 432, 327–344. [Google Scholar] [CrossRef]










| Variable | Units | Description |
|---|---|---|
| T max | °C 10−1 | Monthly average maximum temperature |
| T min | °C 10−1 | Monthly average minimum temperature |
| BIO1 | °C 10−1 | Annual mean temperature |
| BIO2 | °C 10−1 | Mean diurnal range (mean of monthly (max temp–min temp)) |
| BIO3 | % | Isothermality |
| BIO4 | % | Temperature seasonality (standard deviation * 100) |
| BIO5 | °C 10−1 | Maximum temperature of the warmest month |
| T max Aug | °C 10−1 | Maximum temperature in August |
| BIO6 | °C 10−1 | Minimum temperature of the coldest month (i.e., winter frost) |
| T min Jan | °C 10−1 | Minimum temperature in January |
| BIO7 | °C 10−1 | Temperature annual range |
| BIO8 | °C 10−1 | Mean temperature of the wettest quarter |
| BIO9 | °C 10−1 | Mean temperature of the driest quarter |
| BIO10 | °C 10−1 | Mean temperature of the warmest quarter |
| BIO11 | °C 10−1 | Mean temperature of the coldest quarter |
| BIO12 | mm | Annual precipitation |
| BIO13 | mm | Precipitation of the wettest month |
| BIO14 | mm | Precipitation of the driest month |
| BIO15 | % | Precipitation seasonality (coefficient of variation) |
| BIO16 | mm | Precipitation of the wettest quarter |
| BIO17 | mm | Precipitation of the driest quarter |
| BIO18 | mm | Precipitation of the warmest quarter |
| BIO19 | mm | Precipitation of the coldest quarter |
| E | m | Elevation—The vertical distance measured between a point and a datum (a reference surface) which is usually the mean sea level (MSL) |
| S | % | Slope—The rate of change of elevation for each digital elevation model (DEM) cell (i.e., the first derivative of a DEM) |
| A | ° | Aspect—The orientation of slope measured clockwise in degrees from 0 to 360, where 0 is north-facing, 90 is east-facing, 180 is south-facing, and 270 is west-facing. |
| WRBFU | Soil codes from the international soil classification system for naming soils and creating legends for soil maps |
| Variable | Units | Min. | Max. | Mean | Std. dev. | Min | Max. | Mean | Std. dev. |
|---|---|---|---|---|---|---|---|---|---|
| NFI4 plots in pure maritime pine stands (n = 744) | NFI4 plots in pure eucalypts stands (n = 615) | ||||||||
| N | trees ha−1 | 10 | 2510 | 399 | 371 | 10 | 2770 | 607 | 486 |
| G | m2 ha−1 | 0.2 | 57.5 | 13.1 | 11.2 | 0.2 | 59.3 | 8.4 | 6.8 |
| V | m3 ha−1 | 0.0 | 440.6 | 90.7 | 86.9 | 0.0 | 704.9 | 53.7 | 56.9 |
| ddom | cm | 7.6 | 61.6 | 28.1 | 10.4 | 7.5 | 92 | 19.8 | 11.6 |
| hdom | m | 3.2 | 31.0 | 15.3 | 5.6 | 6.0 | 43.6 | 17.6 | 6.1 |
| t | years | 6 | 80 | 41 | 16 | 2 | 60 | 10 | 6 |
| Temperature Range (°C) | Temperature Limits (°C) | Precipitation (mm) | Elevation (m) | Lithology | |
|---|---|---|---|---|---|
| Maritime pine | T max Aug–T min Jan < 26 | T max Aug < 29.9 | P > 850 | E < 800 | different of Limestone |
| Eucalypts | T max Aug–T min Jan < 26 | T max Aug <31 T min Jan > 2 | P > 600 | E < 500 | different of Limestone and Wind alluvial sands |
| Species | Site Index Model |
|---|---|
| Maritime pine SI50—Dominant height at the reference age of 50 years | with, where hdom—Dominant height (m); t—Stand age (y); P—Annual precipitation (mm; BIO12); T—Annual mean temperature (°C; BIO1); WINTER—Type of winter in a scale from 1 to 5 (1—warm (<7 °C); 2—temperate (3–7 °C); 3—fresh (0–3 °C); 4—cold (−3–0 °C); 5—very cold (<−3 °C; BIO6); ST1, ST2, ST3—Dummy variables for humic cambisols, rankers and calcic cambisols, respectively. |
| Eucalypts SI10—Dominant height at the reference age of 10 years | where pd—number of precipitation days per year with values greater than 0.1 mm; hdom—dominant height (m); t—stand age (y). |
| Class | Productivity | Maritime Pine SI50 (m) | Eucalypts SI10 (m) |
|---|---|---|---|
| 1 | High | ≥24 | ≥23 |
| 2 | Medium-High | 20–24 | 20–23 |
| 3 | Medium | 16–20 | 17–20 |
| 4 | Low-Medium | 12–16 | 14–17 |
| 5 | Low | <12 | <14 |
| Variable | Units | Min. | Max. | Mean | Std. Dev. | Min. | Max. | Mean | Std. dev. |
|---|---|---|---|---|---|---|---|---|---|
| Maritime pine—Pb95 > 0 (n = 23,752) | Maritime pine—SI50 (n = 740) | ||||||||
| BIO5 | °C | 21.8 | 32.7 | 27.5 | 2.2 | 22.1 | 31.9 | 27.3 | 2.1 |
| T max Aug | °C | 21.8 | 32.7 | 27.5 | 2.2 | 20.9 | 28.9 | 24.8 | 1.4 |
| BIO6 | °C | −1.4 | 9.0 | 5.4 | 1.7 | −1.1 | 8.3 | 4.8 | 1.9 |
| T min Jan | °C | −2.4 | 9.3 | 3.4 | 2.0 | −2.6 | 8.4 | 3.0 | 2.0 |
| BIO7 | °C | 13.3 | 28.2 | 22.1 | 2.9 | 14.1 | 27.9 | 22.5 | 2.9 |
| T max Aug–T min Jan | °C | 14.1 | 31.0 | 24.1 | 3.2 | 13.6 | 27.4 | 21.8 | 2.4 |
| BIO12 | mm | 468.0 | 1587.0 | 968.3 | 221.3 | 483.0 | 1593.0 | 1015.2 | 211.7 |
| E | m | 0.0 | 1192.0 | 264.9 | 200.7 | 0.0 | 1275.0 | 329.1 | 248.7 |
| WRBFU | 2.0 | 129.0 | 82.8 | 37.2 | 27.0 | 124.0 | 83.8 | 38.1 | |
| Eucalypts—Ec95 > 0 (n = 34,730) | Eucalypts—SI10 (n = 614) | ||||||||
| BIO5 | °C | 21.6 | 33.9 | 27.9 | 2.4 | 22.4 | 32.7 | 27.6 | 2.4 |
| T max Aug | °C | 21.6 | 33.5 | 27.9 | 2.4 | 20.8 | 29.9 | 25.5 | 1.7 |
| BIO6 | °C | −1.4 | 9.1 | 5.7 | 1.6 | 1.4 | 8.4 | 5.6 | 1.5 |
| T min Jan | °C | −2.4 | 9.4 | 3.5 | 1.6 | 0.3 | 7.7 | 3.6 | 1.4 |
| BIO7 | °C | 12.9 | 29.1 | 22.3 | 2.8 | 14.4 | 27.8 | 22.0 | 2.7 |
| T max Aug–Tmin Jan | °C | 13.0 | 33.0 | 24.4 | 3.3 | 14.7 | 28.0 | 21.9 | 2.3 |
| BIO12 | mm | 468.0 | 1594.0 | 900.3 | 248.2 | 503.0 | 1542.0 | 937.1 | 246.8 |
| E | m | 0.0 | 1192.0 | 245.4 | 184.7 | 8.0 | 752.0 | 231.2 | 155.3 |
| WRBFU | 2.0 | 129.0 | 82.9 | 36.3 | 27.0 | 127.0 | 85.3 | 36.1 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alegria, C.; Roque, N.; Albuquerque, T.; Gerassis, S.; Fernandez, P.; Ribeiro, M.M. Species Ecological Envelopes under Climate Change Scenarios: A Case Study for the Main Two Wood-Production Forest Species in Portugal. Forests 2020, 11, 880. https://doi.org/10.3390/f11080880
Alegria C, Roque N, Albuquerque T, Gerassis S, Fernandez P, Ribeiro MM. Species Ecological Envelopes under Climate Change Scenarios: A Case Study for the Main Two Wood-Production Forest Species in Portugal. Forests. 2020; 11(8):880. https://doi.org/10.3390/f11080880
Chicago/Turabian StyleAlegria, Cristina, Natália Roque, Teresa Albuquerque, Saki Gerassis, Paulo Fernandez, and Maria Margarida Ribeiro. 2020. "Species Ecological Envelopes under Climate Change Scenarios: A Case Study for the Main Two Wood-Production Forest Species in Portugal" Forests 11, no. 8: 880. https://doi.org/10.3390/f11080880