Response of Soil Microbial Community to C:N:P Stoichiometry along a Caragana korshinskii Restoration Gradient on the Loess Plateau, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Experimental Design
2.2. Sampling
2.3. Laboratory Analysis
2.4. DNA Extraction and Amplification
2.5. Processing the Sequencing Data
2.6. Statistical Analyses
3. Results
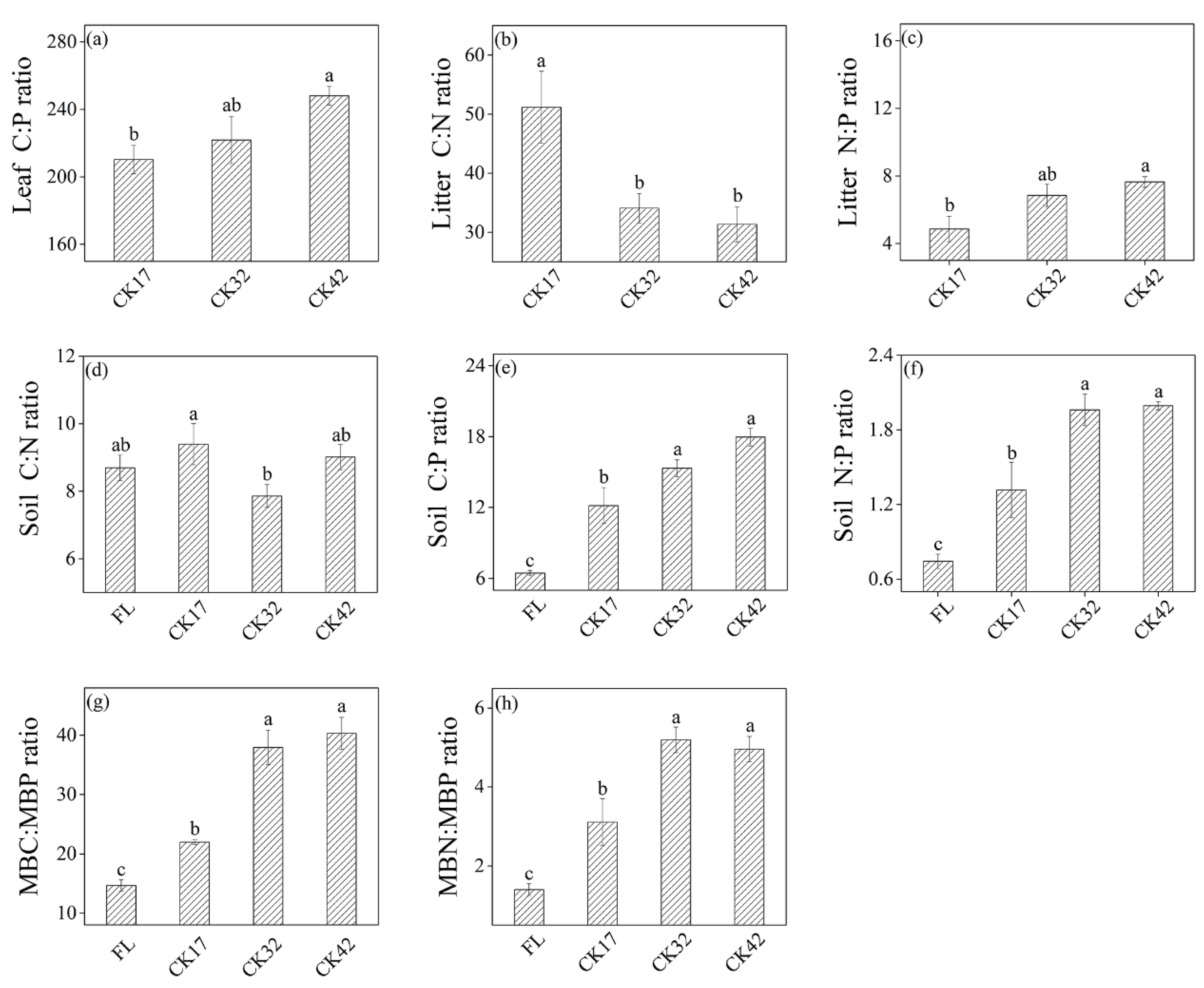
3.1. C, N, and P Contents and C:N:P Stoichiometry
3.1.1. C, N, and P Contents and C:N:P Stoichiometry in Plants
3.1.2. C, N, and P Contents and C:N:P Stoichiometry in the Soil–Microbe Continuum
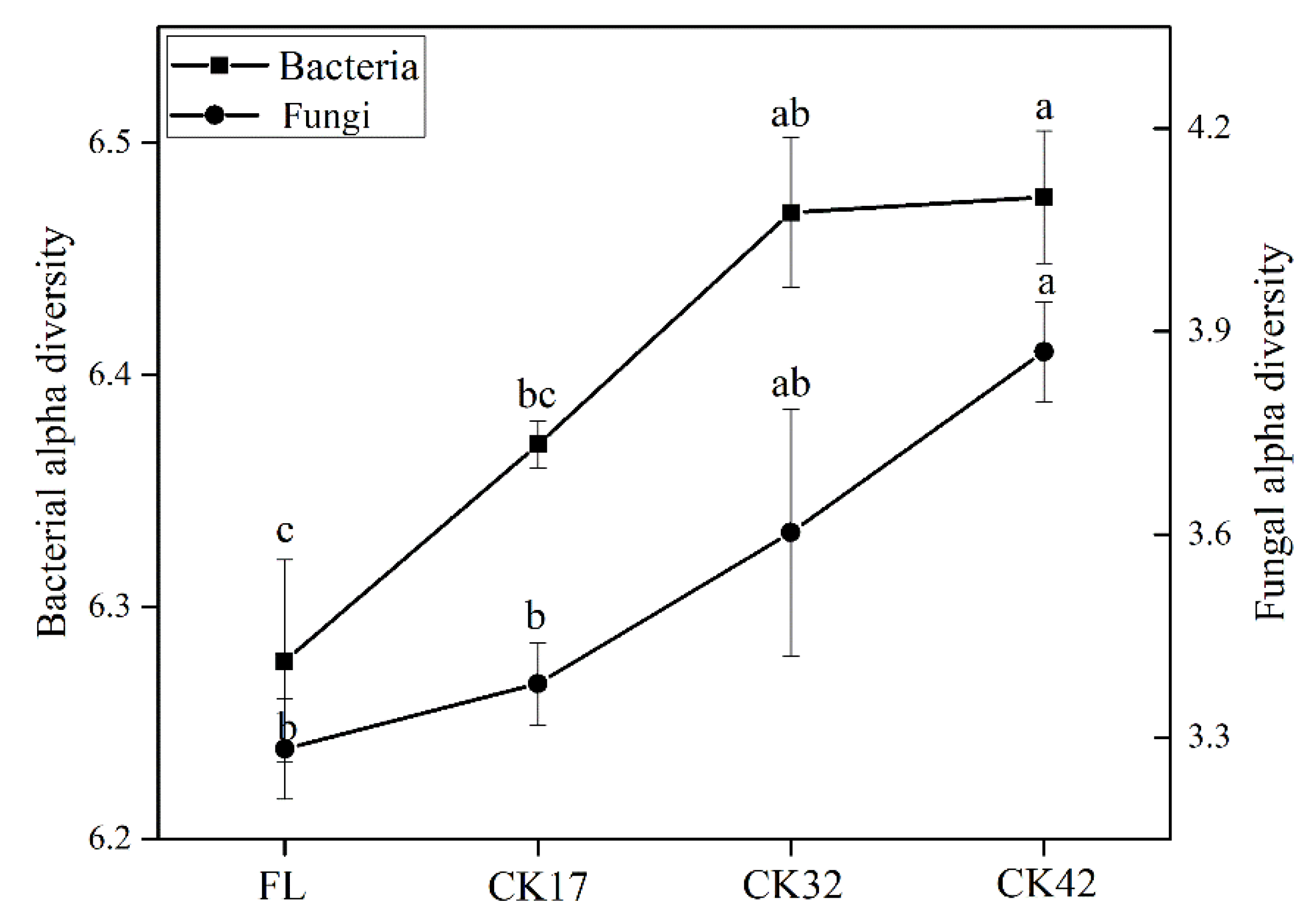
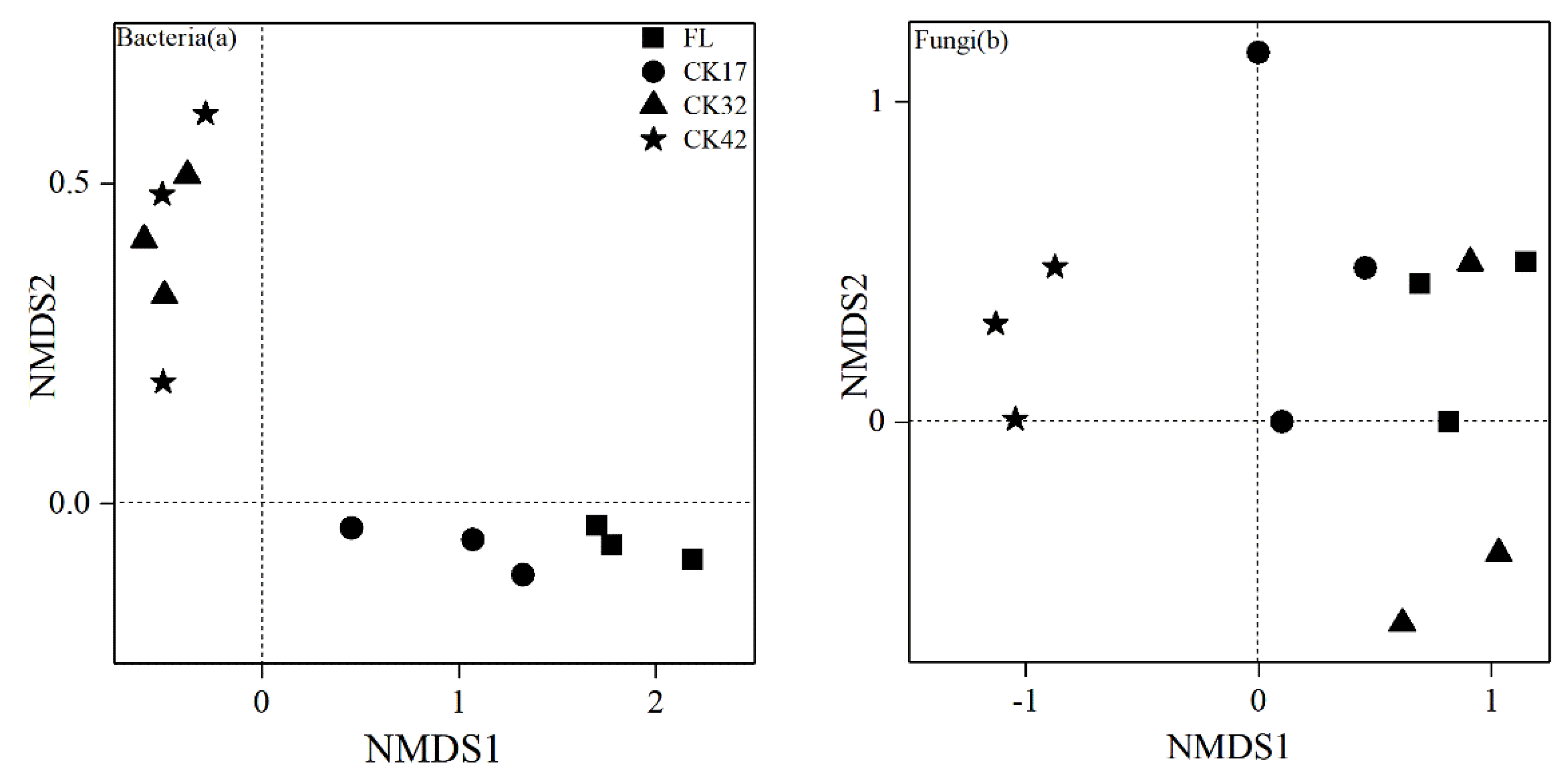
3.2. Changes in Diversity and Composition of Microbial Community
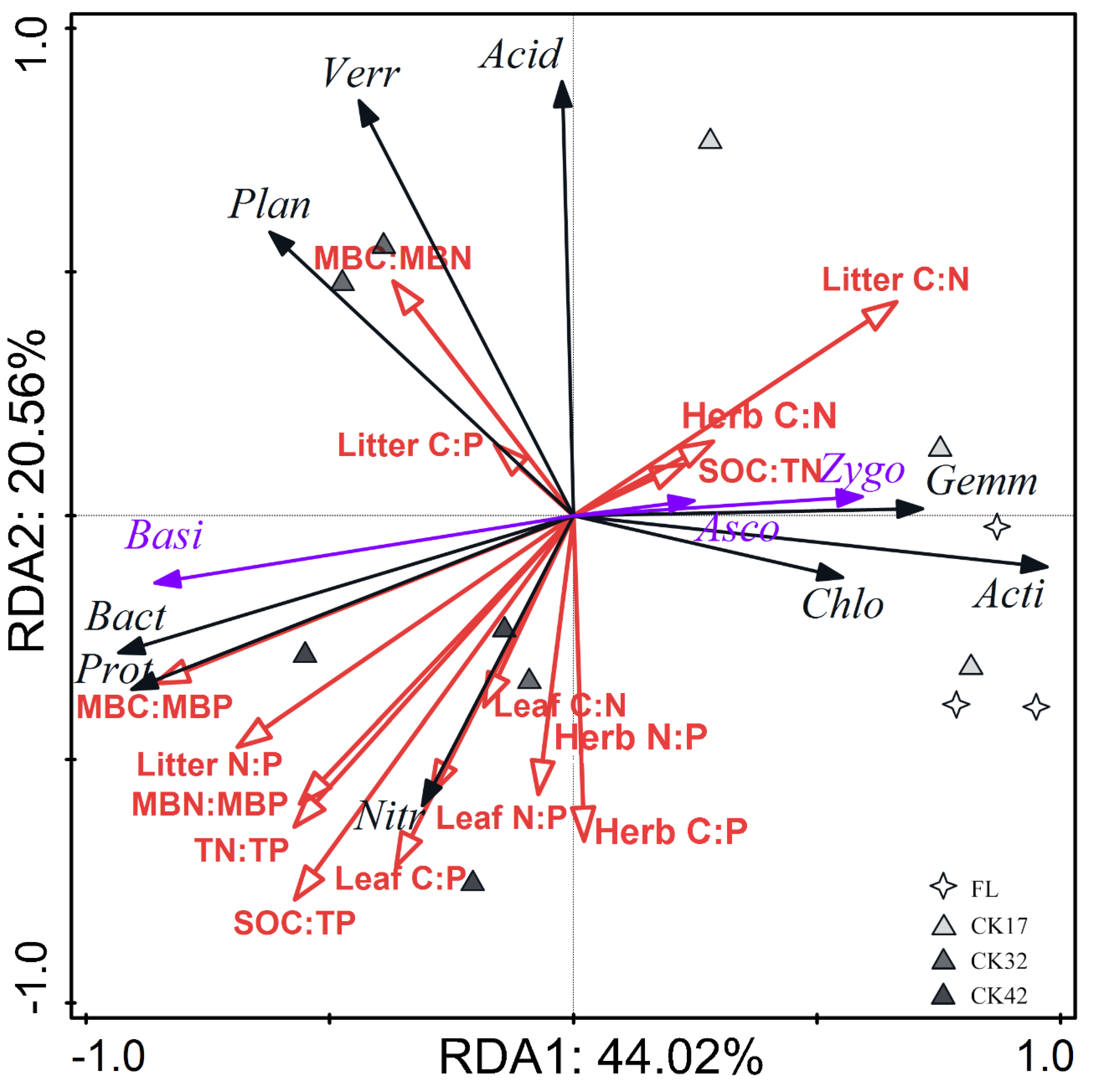
3.3. Correlation of Soil Microbial Composition and Diversity with C:N:P Stoichiometry
4. Discussion
4.1. Changes in the C:N:P Stoichiometry in the Plant–Soil–Microbe Continuum during Afforestation
4.2. Response of Microbial Community to the C:N:P Stoichiometry in the Plant–Soil–Microbe Continuum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deng, J.; Sun, P.S.; Zhao, F.Z.; Han, X.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Soil C, N, P and its stratification ratio affected by artificial vegetation in subsoil, Loess Plateau China. PLoS ONE 2016, 11, e0151446. [Google Scholar] [CrossRef]
- Ren, C.J.; Chen, J.; Deng, J.; Zhao, F.Z.; Han, X.H.; Yang, G.H.; Tong, X.G.; Feng, Y.Z.; Shelton, S.; Ren, G.X. Response of microbial diversity to C:N:P stoichiometry in fine root and microbial biomass following afforestation. Biol. Fert. Soils 2017, 53, 457–468. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.B.; Xue, S.; Song, Z.L. Rhizosphere soil microbial activity under different vegetation types on the Loess Plateau, China. Geoderma 2011, 161, 115–125. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, C.J.; Zhao, F.Z.; Kang, D.; Yang, G.H.; Han, X.H.; Tong, X.G.; Feng, Y.Z.; Ren, G.X. Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmland. For. Ecol. Manag. 2016, 376, 59–66. [Google Scholar] [CrossRef]
- Fanin, N.; Fromin, N.; Buatois, B.; Hattenschwiler, S. An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system. Ecol. Lett. 2013, 16, 764–772. [Google Scholar] [CrossRef]
- Bui, E.N.; Henderson, B.L. C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 2013, 373, 553–568. [Google Scholar] [CrossRef]
- Fan, H.B.; Wu, J.P.; Liu, W.F.; Yuan, Y.H.; Hu, L.; Cai, Q.K. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 2015, 392, 127–138. [Google Scholar] [CrossRef]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Richardson, S.J.; Allen, R.B.; Doherty, J.E. Shifts in leaf N:P ratio during resorption reflect soil P in temperate rainforest. Funct. Ecol. 2008, 22, 738–745. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, Y.M. Coupling of plant and soil C:N:P stoichiometry in black locust (Robinia pseudoacacia) plantations on the Loess Plateau, China. Trees Struct. Funct. 2017, 31, 1559–1570. [Google Scholar] [CrossRef]
- Liu, J.L.; Ha, V.N.; Shen, Z.; Dang, P.; Zhu, H.L.; Zhao, F.; Zhao, Z. Response of the rhizosphere microbial community to fine root and soil parameters following Robinia pseudoacacia L. afforestation. Appl. Soil Ecol. 2018, 132, 11–19. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chen, L.Y.; Peng, Y.F.; Ding, J.Z.; Li, F.; Yang, G.B.; Kou, D.; Liu, L.; Fang, K.; Zhang, B.B.; et al. Linking microbial C:N:P stoichiometry to microbial community and abiotic factors along a 3500-km grassland transect on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2016, 25, 1416–1427. [Google Scholar] [CrossRef]
- Mao, C.L.; Wang, Y.B.; Wang, X.J.; Ren, G.X.; Yuan, L.Y.; Feng, Y.Z. Correlations between microbial community and C:N:P stoichiometry during the anaerobic digestion process. Energy 2019, 174, 687–695. [Google Scholar] [CrossRef]
- Erlacher, A.; Cernava, T.; Cardinale, M.; Soh, J.; Sensen, C.W.; Grube, M.; Berg, G. Rhizobiales as functional and endosymbiontic members in the lichen symbiosis of Lobaria pulmonaria L. Front. Microbiol. 2015, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, G.B.; Xue, S.; Wang, G.L. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- McCaig, A.E.; Glover, L.A.; Prosser, J.I. Molecular analysis of bacterial community structure and diversity in unimproved and improved upland grass pastures. Appl. Environ. Microbiol. 1999, 65, 1721–1730. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Axelrood, P.E.; Chow, M.L.; Radomski, C.C.; McDermott, J.M.; Davies, J. Molecular characterization of bacterial diversity from British Columbia forest soils subjected to disturbance. Can. J. Microbiol. 2002, 48, 655–674. [Google Scholar] [CrossRef]
- Zhong, Z.K.; Wang, X.; Zhang, X.Y.; Zhang, W.; Xu, Y.D.; Ren, C.J.; Han, X.H.; Yang, G.H. Edaphic factors but not plant characteristics mainly alter soil microbial properties along a restoration chronosequence of Pinus tabulaeformis stands on Mt. Ziwuling, China. For. Ecol. Manag. 2019, 453. [Google Scholar] [CrossRef]
- Bai, Z.G.; Dent, D. Recent land degradation and improvement in China. Amibo 2009, 38, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.J.; Chen, L.D.; Ma, K.M.; Zhou, H.F.; Wang, J. The relationships between land use and soil conditions in the hilly area of the loess plateau in northern Shaanxi, China. Catena 2000, 39, 69–78. [Google Scholar] [CrossRef]
- Cao, C.Y.; Jiang, D.M.; Teng, X.H.; Jiang, Y.; Liang, W.J.; Cui, Z.B. Soil chemical and microbiological properties along a chronosequence of Caragana microphylla Lam. plantations in the Horqin sandy land of Northeast China. Appl. Soil Ecol. 2008, 40, 78–85. [Google Scholar] [CrossRef]
- Jiang, D.M.; Li, Q.; Liu, F.M.; Jiang, Y.; Liang, W.J. Vertical distribution of soil nematodes in an age sequence of Caragana microphylla plantations in the Horqin Sandy Land, Northeast China. Ecol. Res. 2006, 22, 49–56. [Google Scholar] [CrossRef]
- Fu, X.L.; Shao, M.A.; Wei, X.R.; Horton, R. Soil organic carbon and total nitrogen as affected by vegetation types in Northern Loess Plateau of China. Geoderma 2010, 155, 31–35. [Google Scholar] [CrossRef]
- Zhong, Z.K.; Han, X.H.; Xu, Y.; Zhang, W.; Fu, S.Y.; Liu, W.C.; Ren, C.J.; Yang, G.H.; Ren, G.X. Effects of land use change on organic carbon dynamics associated with soil aggregate fractions on the Loess Plateau, China. Land Degrad. Dev. 2019, 30, 1070–1082. [Google Scholar] [CrossRef]
- Zhang, W.; Qiao, W.J.; Gao, D.X.; Dai, Y.Y.; Deng, J.; Yang, G.H.; Han, X.H.; Ren, G.X. Relationship between soil nutrient properties and biological activities along a restoration chronosequence of Pinus tabulaeformis plantation forests in the Ziwuling Mountains, China. Catena 2018, 161, 85–95. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.B.; Xue, S.; Wang, G.L. Changes in rhizospheric microbial community structure and function during the natural recovery of abandoned cropland on the Loess Plateau, China. Ecol. Eng. 2015, 75, 161–171. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.B.; Song, Z.L.; Wang, J.; Guo, L. Interactions of soil bacteria and fungi with plants during long-term grazing exclusion in semiarid grasslands. Soil Biol. Biochem. 2018, 124, 47–58. [Google Scholar] [CrossRef]
- Kou, M.; Garcia-Fayos, P.; Hu, S.; Jiao, J.Y. The effect of Robinia pseudoacacia afforestation on soil and vegetation properties in the Loess Plateau (China): A chronosequence approach. For. Ecol. Manag. 2016, 375, 146–158. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Phosphorus in the soil microbial biomass. Soil Biol. Biochem. 1984, 16, 169–175. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biddle, J.F.; Fitz-Gibbon, S.; Schuster, S.C.; Brenchley, J.E.; House, C.H. Metagenomic signatures of the Peru Margin subseafloor biosphere show a genetically distinct environment. Proc. Natl. Acad. Sci. USA 2008, 105, 10583–10588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, P.K.; Chandra, J.; Retuerto, M.; Sikaroodi, M.; Brown, R.E.; Jurevic, R.; Salata, R.A.; Lederman, M.M.; Gillevet, P.M.; Ghannoum, M.A. Oral mycobiome analysis of HIV-infected patients: Identification of Pichia as an antagonist of opportunistic fungi. PLoS Pathog. 2014, 10, e1003996. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.K.; Wang, Y.P.; Wang, S.L.; He, T.X.; Liu, L. Fresh carbon and nitrogen inputs alter organic carbon mineralization and microbial community in forest deep soil layers. Soil Biol. Biochem. 2014, 72, 145–151. [Google Scholar] [CrossRef]
- Hu, Y.L.; Zeng, D.H.; Ma, X.Q.; Chang, S.X. Root rather than leaf litter input drives soil carbon sequestration after afforestation on a marginal cropland. For. Ecol. Manag. 2016, 362, 38–45. [Google Scholar] [CrossRef]
- Lai, Z.R.; Zhang, Y.Q.; Liu, J.B.; Wu, B.; Qin, S.G.; Fa, K.Y. Fine-root distribution, production, decomposition, and effect on soil organic carbon of three revegetation shrub species in northwest China. For. Ecol. Manag. 2016, 359, 381–388. [Google Scholar] [CrossRef]
- Fan, J.; Wang, J.Y.; Hu, X.F.; Chen, F.S. Seasonal dynamics of soil nitrogen availability and phosphorus fractions under urban forest remnants of different vegetation communities in Southern China. Urban For. Urban Green. 2014, 13, 576–585. [Google Scholar] [CrossRef]
- Zhu, H.H.; He, X.Y.; Wang, K.L.; Su, Y.R.; Wu, J.S. Interactions of vegetation succession, soil bio-chemical properties and microbial communities in a Karst ecosystem. Eur. J. Soil Biol. 2012, 51, 1–7. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Kang, D.; Han, X.H.; Yang, G.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Soil stoichiometry and carbon storage in long-term afforestation soil affected by understory vegetation diversity. Ecol. Eng. 2015, 74, 415–422. [Google Scholar] [CrossRef]
- Zhang, C.; Xue, S.; Liu, G.B.; Song, Z.L. A comparison of soil qualities of different revegetation types in the Loess Plateau, China. Plant Soil 2011, 347, 163–178. [Google Scholar] [CrossRef]
- O’Dea, J.K.; Jones, C.A.; Zabinski, C.A.; Miller, P.R.; Keren, I.N. Legume, cropping intensity, and N-fertilization effects on soil attributes and processes from an eight-year-old semiarid wheat system. Nutr. Cycl. Agroecosyst. 2015, 102, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.W.; Syers, J.K. The fate of phosphorus during pedogenesis. Geoderma 1976, 15, 1–19. [Google Scholar] [CrossRef]
- Vance, C.P. Symbiotic nitrogen fixation and phosphorus acquisition. Plant nutrition in a world of declining renewable resources. Plant Physiol. 2001, 127, 390–397. [Google Scholar] [CrossRef]
- Siles, J.A.; Margesin, R. Abundance and diversity of bacterial, archaeal, and fungal communities along an altitudinal gradient in alpine forest soils: What are the driving factors? Microb. Ecol. 2016, 72, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.Z.; Bai, L.; Wang, J.Y.; Deng, J.; Ren, C.J.; Han, X.H.; Yang, G.H.; Wang, J. Change in soil bacterial community during secondary succession depend on plant and soil characteristics. Catena 2019, 173, 246–252. [Google Scholar] [CrossRef]
- Liu, L.; Gundersen, P.; Zhang, W.; Zhang, T.; Chen, H.; Mo, J. Effects of nitrogen and phosphorus additions on soil microbial biomass and community structure in two reforested tropical forests. Sci. Rep. 2015, 5, 14378. [Google Scholar] [CrossRef]
- Xiang, X.J.; Shi, Y.; Yang, J.; Kong, J.J.; Lin, X.G.; Zhang, H.Y.; Zeng, J.; Chu, H.Y. Rapid recovery of soil bacterial communities after wildfire in a Chinese boreal forest. Sci. Rep. 2014, 4, 3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.M.; Wang, R.Q.; Liu, J.; Wang, M.C.; Zhou, J.; Guo, W.H. Effects of vegetation type on soil microbial community structure and catabolic diversity assessed by polyphasic methods in North China. J. Environ. Sci. 2007, 19, 1228–1234. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and associated fungi drive long-term carbon sequestration in boreal forest. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Stursova, M.; Zifcakova, L.; Leigh, M.B.; Burgess, R.; Baldrian, P. Cellulose utilization in forest litter and soil: Identification of bacterial and fungal decomposers. FEMS Microbiol. Ecol. 2012, 80, 735–746. [Google Scholar] [CrossRef]
- Elser, J.J.; Acharya, K.; Kyle, M.; Cotner, J.; Makino, W.; Markow, T.; Watts, T.; Hobbie, S.; Fagan, W.; Schade, J.; et al. Growth rate-stoichiometry couplings in diverse biota. Ecol. Lett. 2003, 6, 936–943. [Google Scholar] [CrossRef] [Green Version]
- Pii, Y.; Mimmo, T.; Tomasi, N.; Terzano, R.; Cesco, S.; Crecchio, C. Microbial interactions in the rhizosphere: Beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. A review. Biol. Fertil. Soils 2015, 51, 403–415. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Elser, J.J.; Dobberfuhl, D.R.; MacKay, N.A.; Schampel, J.H. Organism size, life history, and N:P stoichiometry. BioScience 1996, 46, 674–684. [Google Scholar] [CrossRef] [Green Version]
- Bastida, F.; Hernández, T.; Albaladejo, J.; García, C. Phylogenetic and functional changes in the microbial community of long-term restored soils under semiarid climate. Soil Biol. Biochem. 2013, 65, 12–21. [Google Scholar] [CrossRef]
- Hannula, S.E.; Boschker, H.T.; de Boer, W.; van Veen, J.A. 13C pulse-labeling assessment of the community structure of active fungi in the rhizosphere of a genetically starch-modified potato (Solanum tuberosum) cultivar and its parental isoline. New Phytol. 2012, 194, 784–799. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Restoration Stages A | Geographical Coordinates | Elevation (m) | Slope Gradient (°) | Slope Aspect (°) B | Crown Density (%) C | Understory Coverage (%) C | Clay Content (%) | Silt Content (%) | Sand Content (%) | SBD (g·m−3) | SWC (%) | pH |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FL | 36°51′54.6″ N, 109°21′4.9″ E | 1205.0 | 15.4 | NbyW75 | - | - | 20.49 (0.66) a | 45.59 (1.01) b | 33.92 (0.39) | 1.30 (0.00) a | 9.77 (0.05) c | 8.24 (0.05) |
| CK17 | 36°44′25.1″ N, 109°15′41.0″ E | 1280.0 | 38.5 | SbyW63 | 43 | 60 | 18.83 (0.45) b | 46.58 (0.85) b | 34.59 (0.43) | 1.21 (0.02) b | 10.30 (0.44) bc | 8.17 (0.01) |
| CK32 | 36°51′55.5″ N, 109°21′0.9″ E | 1259.8 | 35.7 | SbyW47 | 55 | 75 | 17.51 (0.30) b | 51.48 (2.04) ab | 31.01 (1.74) | 1.12 (0.02) c | 11.66 (0.43) ab | 8.18 (0.15) |
| CK42 | 36°52′11.7″ N, 109°21′0.2″ E | 1307.8 | 39.1 | NbyE85 | 62 | 70 | 15.74 (0.09) c | 55.27 (2.97) a | 29.00 (2.99) NS | 1.04 (0.02) d | 12.97 (0.61) a | 8.07 (0.01) NS |
| Variables | Restoration Stages A | |||
|---|---|---|---|---|
| FL B | CK17 | CK32 | CK42 | |
| Leaf OC (g·kg−1) | - | 427.63 (29.45) | 438.80 (13.31) | 475.61 (4.95) NS |
| Leaf TN (g·kg−1) | - | 34.93 (0.11) | 34.15 (0.08) | 35.48 (1.72) NS |
| Leaf TP (g·kg−1) | - | 2.03 (0.06) | 1.99 (0.09) | 1.92 (0.03) NS |
| Herb OC (g·kg−1) | - | 422.31 (22.27) | 432.40 (10.80) | 434.62 (7.33) NS |
| Herb TN (g·kg−1) | - | 11.91 (0.77) | 12.89 (0.99) | 13.67 (0.28) NS |
| Herb TP (g·kg−1) | - | 1.95 (0.37) | 1.78 (0.36) | 1.53 (0.03) NS |
| Litter OC (g·kg−1) | - | 389.36 (7.04) b | 413.03 (6.15) b | 457.72 (6.48) a |
| Litter TN (g·kg−1) | - | 7.82 (0.88) b | 12.24 (0.91) a | 14.83 (1.24) a |
| Litter TP (g·kg−1) | - | 1.66 (0.21) | 1.80 (0.14) | 1.94 (0.14) NS |
| SOC (g·kg−1) | 2.98 (0.05) d | 5.95 (0.33) c | 7.73 (0.19) b | 9.28 (0.31) a |
| TN (g·kg−1) | 0.34 (0.01) c | 0.64 (0.05) b | 0.99 (0.05) a | 1.03 (0.02) a |
| TP (g·kg−1) | 0.46 (0.02) | 0.50 (0.04) | 0.50 (0.01) | 0.52 (0.01) NS |
| MBC (mg·kg−1) | 71.69 (2.16) d | 176.81 (11.71) c | 323.38 (11.00) b | 427.29 (13.90) a |
| MBN (mg·kg−1) | 6.82 (0.49) c | 24.48 (3.47) b | 44.57 (3.23) a | 52.66 (2.44) a |
| MBP (mg·kg−1) | 4.90 (0.18) c | 8.05 (0.45) b | 8.60 (0.57) b | 10.65 (0.35) a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Li, W.; Zhong, Z.; Zhang, Q.; Wang, X.; Han, X.; Ren, C.; Yang, G. Response of Soil Microbial Community to C:N:P Stoichiometry along a Caragana korshinskii Restoration Gradient on the Loess Plateau, China. Forests 2020, 11, 823. https://doi.org/10.3390/f11080823
Zhang X, Li W, Zhong Z, Zhang Q, Wang X, Han X, Ren C, Yang G. Response of Soil Microbial Community to C:N:P Stoichiometry along a Caragana korshinskii Restoration Gradient on the Loess Plateau, China. Forests. 2020; 11(8):823. https://doi.org/10.3390/f11080823
Chicago/Turabian StyleZhang, Xinyi, Wenjie Li, Zekun Zhong, Qingyue Zhang, Xing Wang, Xinhui Han, Chengjie Ren, and Gaihe Yang. 2020. "Response of Soil Microbial Community to C:N:P Stoichiometry along a Caragana korshinskii Restoration Gradient on the Loess Plateau, China" Forests 11, no. 8: 823. https://doi.org/10.3390/f11080823