Formula Fertilization Promotes Phoebe bournei Robust Seedling Cultivation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Materials
2.3. Experimental Design
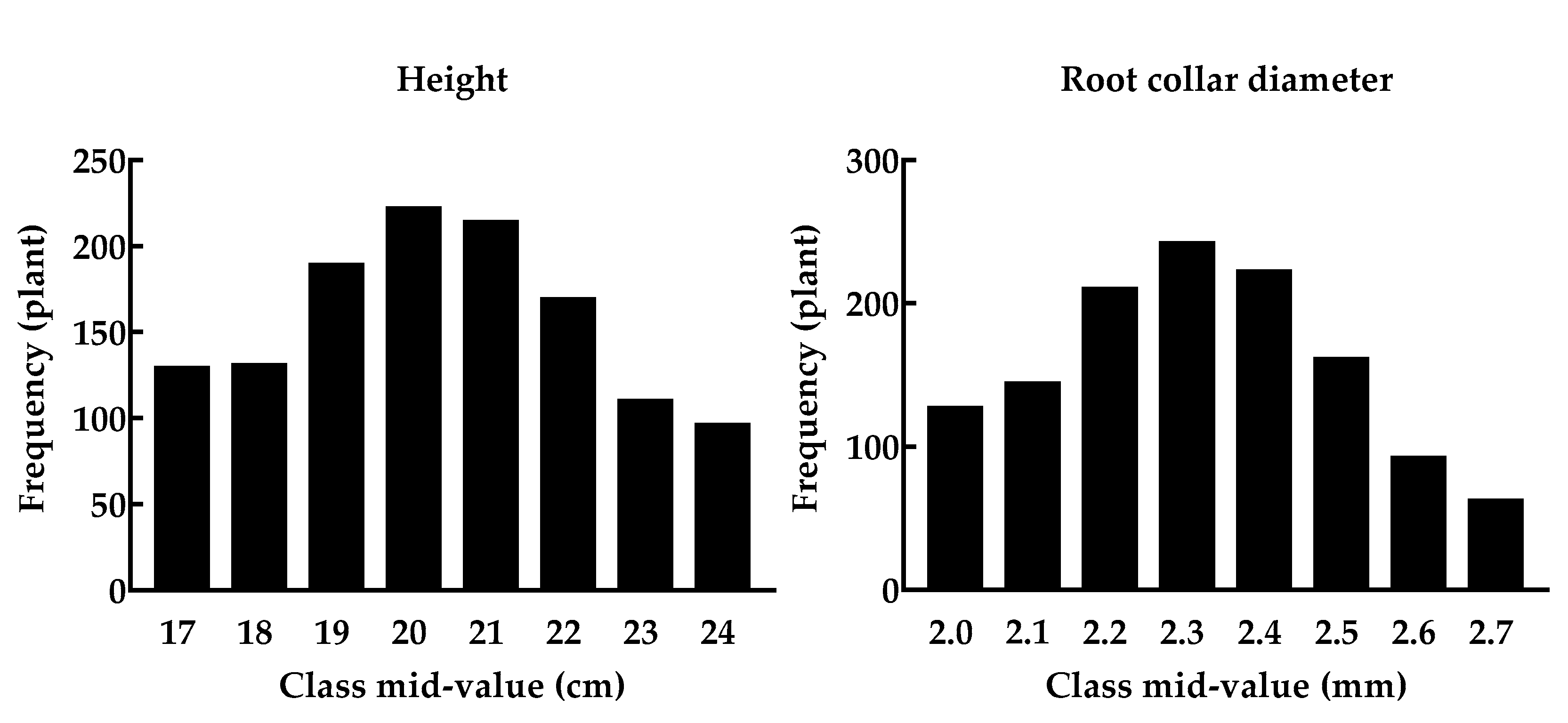
2.4. Measured Seedling Attributes
2.5. Data Processing and Analysis
2.5.1. Calculation of Comprehensive Index
2.5.2. Data Analysis
3. Results
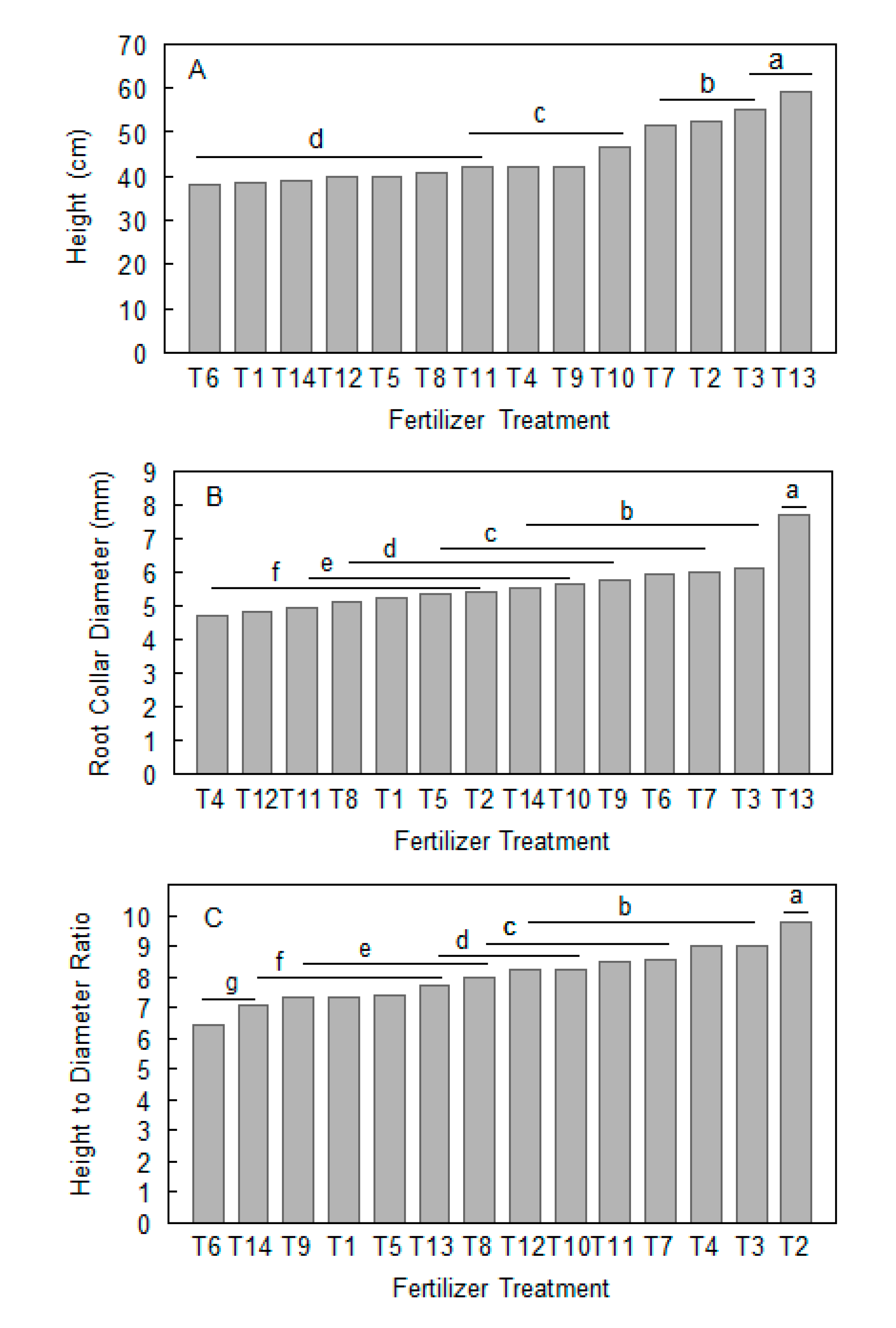
3.1. Effects of Different Fertilization Treatments on Seedling Growth
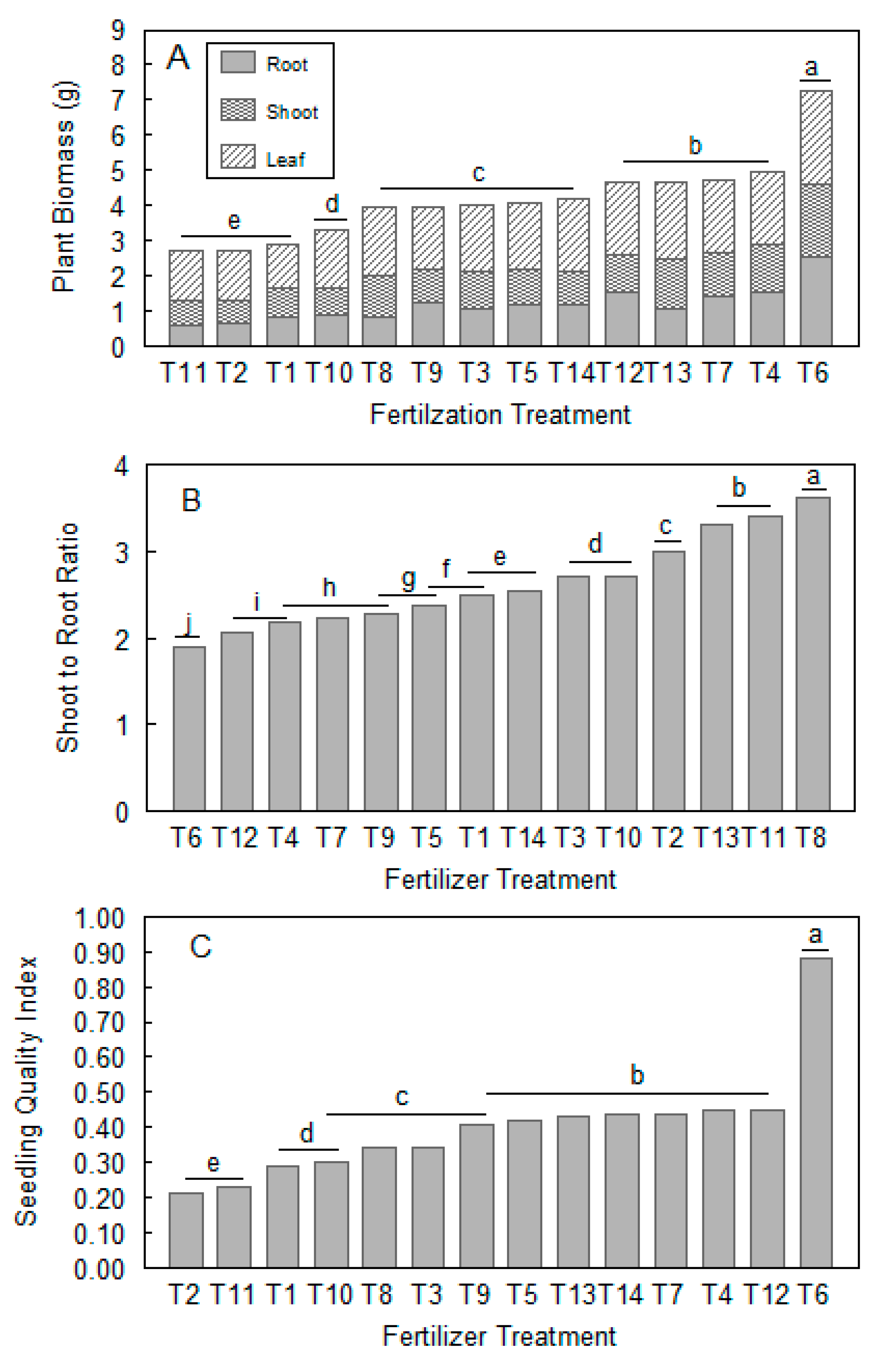
3.2. Effects of Different Fertilization Treatments on Seedling Biomass
3.3. Effects of Different Fertilization Treatments on Seedling Quality Index (QI)
3.4. Correlations among N–P–K Application Rates with Seedling Attributes and QI Index
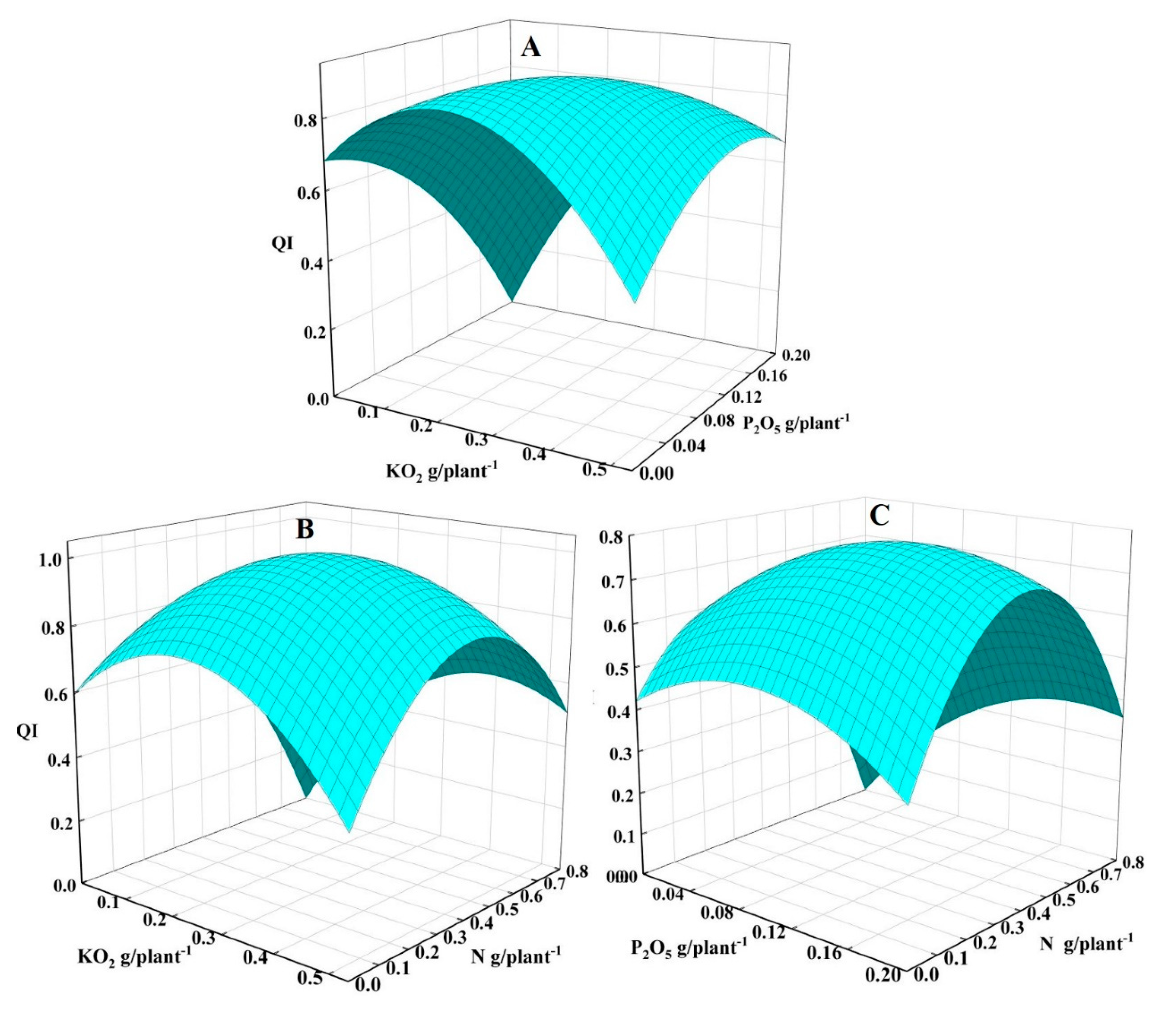
3.5. Interaction Analysis of N–P–K Fertilizer Applications
3.6. Fertilizer Function Model Fitting and Fertilizer Rate Analysis
4. Discussions
4.1. Effects of N–P–K Fertilizer on Phoebe bournei Seedlings Growth
4.2. Effects of N, P, K Fertilizer on Phoebe bournei Seedlings Growth
4.3. Interaction of N–P–K Fertilizer on Phoebe bournei Seedlings Growth
4.4. Optimum Application Rate of N–P–K Fertilizer for Seedlings Production
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, Z.; Shang, H.; Cao, J.; Yu, H. Effects of ambient ozone concentrations on contents of nonstructural carbohydrates in Phoebe bournei and Pinus massoniana seedlings in subtropical China. Water Air Soil Pollut. 2015, 226, 310. [Google Scholar] [CrossRef]
- Ding, W.; Liping, N.; Xing, H.; Wei, Z.; Zhoua, Q.; Nong, R.; Chen, J. Essential oil extracted from leaf of Phoebe bournei (Hemsl.) Yang: Chemical constituents, antitumor, antibacterial, hypoglycemic activities. Nat. Prod. Res. 2018. [Google Scholar] [CrossRef]
- Cao, J.; Shang, H.; Chen, Z.; Tian, Y.; Yu, H. Effects of elevated ozone on stoichiometry and nutrient pools of Phoebe bournei (Hemsl.) Yang and Phoebe Zhennan S. Lee et F. N. Wei seedlings in subtropical China. Forests 2016, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Blanco, J.A. Significant increase in ecosystem C can be achieved with sustainable forest management in subtropical plantation forests. PLoS ONE 2014, 9, e89688. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wei, X.; Liao, W.; Blanco, J.A.; Liu, Y.; Liu, S.; Liu, G.; Lu, Z.; Guo, X.; Guo, S. Evaluation of the effects of forest management strategies on carbon sequestration in evergreen broad-leaved (Phoebe bournei) plantation forests using FORECAST ecosystem model. For. Ecol. Manag. 2013, 300, 21–32. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Drouza, C.; Tzortzakis, N. Optimization of potassium fertilization/nutrition for growth, physiological development, essential oil composition and antioxidant activity of Lavandula angustifolia Mill. J. Soil Sci. Plant Nutr. 2017, 17, 291–306. [Google Scholar] [CrossRef] [Green Version]
- Sloan, J.L.; Uscola, M.; Jacobs, D.F. Nitrogen recovery in planted seedlings, competing vegetation, and soil in response to fertilization on a boreal mine reclamation site. For. Ecol. Manag. 2016, 360, 60–68. [Google Scholar] [CrossRef]
- Tinus, R.W.; McDonald, S.E. How to Grow Tree Seedlings in Containers in Greenhouses; US Department of Agriculture, Forest Service: Portland, OR, USA, 1979; Volume 60.
- Landis, T.D.; Tinus, R.W.; McDonald, S.E.; Barnett, J.P. Seedling nutrition and irrigation. In The Container Tree Nursery Manual; USDA Forest Service Agriculture Handbook 674; USDA Forest Service: Washington, DC, USA, 1989; Volume 4. [Google Scholar]
- Burdett, A.N. Quality control in the production of forest planting stock. For. Chron. 1983, 59, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Landis, T.D. Mineral nutrition as an index of seedling quality. In Evaluating Seedling Quality: Principles, Procedures, and Predictive Ability of Major Tests; Duryea, M.L., Ed.; Oregon State University, Forest Research Laboratory: Corvallis, OR, USA, 1985; pp. 29–48. [Google Scholar]
- Puttonen, P. Criteria for using seedling performance potential tests. New For. 1989, 3, 67–87. [Google Scholar] [CrossRef]
- Toumey, J.W. Seeding and Planting; Wiley: New York, NY, USA, 1916. [Google Scholar]
- Wakeley, P.C. Planting the Southern Pines; USDA Agricultural Monograph 18; US Forest Service: Portland, OR, USA, 1954; p. 233.
- Puttonen, P. Looking for the ‘‘silver’’ bullet-can one test do it all? New For. 1997, 13, 9–27. [Google Scholar] [CrossRef]
- Thompson, B.E. Seedling morphological evaluation: What you can tell by looking. In Evaluating Seedling Quality: Principles, Procedures, and Predictive Ability of Major Tests; Duryea, M.L., Ed.; Oregon State University, Forest Research Laboratory: Corvallis, OR, USA, 1985; pp. 59–72. [Google Scholar]
- Mexal, J.G.; Landis, T.D. Target seedling concepts: Height and diameter. In Target Seedling Symposium: Proceedings: Combined Meeting of the Western Forest Nursery Associations; Rose, R., Campbell, S.J., Landis, T.D., Eds.; RM-GTR-200; USDA Forest Service: Roseburg, OR, USA, 1990; pp. 17–36. [Google Scholar]
- Grossnickle, S.C. Why seedlings survive: Importance of plant attributes. New For. 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Why seedlings grow: Influence of plant attributes. New For. 2018, 49, 1–34. [Google Scholar] [CrossRef]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Quality appraisal of white spruce and white pine seedling stock in nurseries. For. Chron. 1960, 36, 10–13. [Google Scholar] [CrossRef]
- Mattsson, A. Predicting field performance using seedling quality assessment. New For. 1997, 13, 227–252. [Google Scholar] [CrossRef]
- Dong, L.; Zhu, X.; Lin, X.; Xu, Z. Effects of fertilization on the growth of three Lauraceae seedlings in containers. North. Hortic. 2011, 13, 73–77. [Google Scholar]
- Wang, Y.; Wang, X.; Wu, X.; Zhang, L.; Wu, L.; Xu, Y.; Zhou, Z. Effects of slow-release fertilizer loading on growth and construction of nutrients reserves of Phoebe chekiangensis and Phoebe bournei container seedlings. Sci. Silvae Sin. 2013, 49, 57–63. [Google Scholar]
- Ou, J. N, P, K formulas for fertilization with young Phoebe bournei plantations in Fujian province. J. Zhejiang A F Univ. 2015, 32, 92–97. [Google Scholar]
- Li, J.; Zhang, M.; Chen, F.; Yao, B. Improving rice modeling success rate with ternary non-structural fertilizer response model. Sci. Rep. 2018, 8, 9071. [Google Scholar] [CrossRef]
- Ren, T.; Zou, J.; Lu, J.; Chen, F.; Wu, J.; Li, X. On-farm trials of optimal fertilizer recommendations for the maintenance of high seed yields in winter oilseed rape (Brassica napus L.) production. Soil Sci. Plant Nutr. 2015, 61, 528–540. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, J.; Ma, Y.; Jia, Y.; Liang, Q. Apparent accumulated nitrogen fertilizer recovery in long-term wheat–maize cropping systems in China. Agronomy 2018, 8, 293. [Google Scholar] [CrossRef] [Green Version]
- Shi, B.; Wang, Y.; Meng, B.; Zhong, S.; Sun, W. Effects of nitrogen addition on the drought susceptibility of the Leymus chinensis meadow ecosystem vary with drought duration. Front. Plant Sci. 2018, 9, 254. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Chen, M.; Wu, J.; Dong, C.; Huang, X.; Huang, S.; Tang, A.; Huang, M. Nutritional properties and fertilizer demand rules of young Phoebe bournei forest. Agric. Sci. Technol. 2015, 16, 2758. [Google Scholar]
- Magomya, A.M.; Kubmarawa, D.; Ndahi, J.A.; Yebpella, G.G. Determination of plant proteins via the Kjeldahl method and amino acid analysis: A comparative study. Int. J. Sci. Res. 2014, 3, 68–72. [Google Scholar]
- Zhang, S.; Guo, X.; Yun, W.; Xia, Y.; You, Z.; Rillig, M.C. Arbuscular mycorrhiza contributes to the control of phosphorus loss in paddy fields. Plant Soil 2020, 447, 623–636. [Google Scholar] [CrossRef]
- Ishikawa, J.; Fujimura, S.; Kondo, M.; Murai-Hatano, M.; Goto, A.; Shinano, T. Dynamic changes in the Cs distribution throughout rice plants during the ripening period, and effects of the soil-K level. Plant Soil 2018, 429, 503–518. [Google Scholar] [CrossRef]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Seedling quality—Soil fertility relationships of white spruce and, red and white pine in nurseries. For. Chron. 1960, 36, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, G.A.; Landis, T.D.; Dumroese, R.K.; Haase, D.L. Assessing plant quality. In Seedling Processing, Storage, and Outplanting: The Container Tree Nursery Manual; Landis, T.D., Dumroese, R.K., Haase, D.L., Eds.; Agriculture handbook 674; USDA Forest Service: Washington, DC, USA, 2010; Volume 7, pp. 17–82. [Google Scholar]
- Liu, D.P.; Yang, S.Q.; Shi, H.B.; Zheng, X.B.; Sun, L.G.; Chang, C.L. Effect of combined nitrogen and phosphorus fertilizer application of wheat-maize inter-cropping system. Chin. J. Eco Agric. 2014, 22, 262–269. [Google Scholar] [CrossRef]
- Hawkins, B.J. Seedling mineral nutrition, the root of the matter. In National Proceedings: Forest and Conservation Nursery Associations-2010; Riley, L.E., Haase, D.L., Pinto, J.R., Eds.; USDA Forest Service RMRS-P-65; USDA Forest Service: Portland, OR, USA, 2011; p. 65. [Google Scholar]
- Pinto, J.R. Morphology targets: What do seedling morphological attributes tell us? In National Proceedings: Forest and Conservation Nursery Associations-2010; Riley, L.E., Haase, D.L., Pinto, J.R., Eds.; RMRSP-65; USDA Forest Service: Portland, OR, USA, 2011; pp. 74–79. [Google Scholar]
- Jin, G.; Yu, Q.; Jiao, Y.; Wang, Y.; Wang, H.; Zhou, Z. Effects of combined fertilization on young growth of Taxus chinensis var. Mairei. For. Res. 2007, 20, 251–256. [Google Scholar]
- Van den Driessche, R. Effects of nutrients on stock performance in the forest. In Mineral Nutrition of Conifer Seedlings; van den Driessche, R., Ed.; USDA Forest Service: Portland, OR, USA, 1990. [Google Scholar]
- Grossnickle, S.C. Ecophysiology of Northern Spruce Species: The Performance of Planted Seedlings; NRC Research Press: Ottawa, ON, Canada, 2000. [Google Scholar]
- Chapman, H.D. Plant analysis values suggestive of nutrient status of selected crops. In Soil Testing and Plant Analysis. II; Westerman, R., Ed.; Special Publication Series Number 2; Soil Science Society of America: Madison, WI, USA, 1967; pp. 77–92. [Google Scholar]
- Larcher, W. Physiological Plant Ecology, 3rd ed.; Springer: New York, NY, USA, 1995. [Google Scholar]
- Rook, D.A. Seedling development and physiology in relation to mineral nutrition. In Mineral Nutrition of Conifer Seedlings; van den Driessche, R., Ed.; Research Branch Ministry of Forests: Victoria, BC, Canada, 1990; pp. 86–105. [Google Scholar]
- Yuan, T.; Cheng, Y.; Huang, W.; Zhang, Z.; Lei, Z.; Shimizu, K.; Utsumi, M. Fertilizer potential of liquid product from hydrothermal treatment of swine manure. Waste Manag. 2018, 77, 166–171. [Google Scholar] [CrossRef]
- Filipescu, C.N.; Trofymow, J.A.; Koppenaal, R.S. Late-rotation nitrogen fertilization of Douglas-fir: Growth response and fibre properties. Can. J. For. Res. 2016, 47, 134–138. [Google Scholar] [CrossRef]
- Coyle, D.R.; Coleman, M.D.; Aubrey, D.P. Above-and below-ground biomass accumulation, production, and distribution of sweetgum and loblolly pine grown with irrigation and fertilization. Can. J. For. Res. 2008, 38, 1335–1348. [Google Scholar] [CrossRef] [Green Version]
- Pallardy, S.G. Physiology of Woody Plants; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Bulgarelli, R.G.; Marcos, F.C.C.; Ribeiro, R.V.; de Andrade, S.A.L. Mycorrhizae enhance nitrogen fixation and photosynthesis in phosphorus-starved soybean (Glycine max L. Merrill). Environ. Exp. Bot. 2017, 140, 26–33. [Google Scholar] [CrossRef]
- Dong, J.; Xu, Q.; Gruda, N.; Chu, W.; Li, X.; Duan, Z. Elevated and super-elevated CO2 differ in their interactive effects with nitrogen availability on fruit yield and quality of cucumber. J. Sci. Food Agric. 2018, 98, 4509–4516. [Google Scholar] [CrossRef]
- Proe, M.F.; Millard, P. Effect of supply upon seasonal growth and internal cycling of P in Sitka spruce (Picea sitchensis (Bong.) Carr.) seedlings. Plant Soil 1995, 168, 313–317. [Google Scholar] [CrossRef]
- Teng, Y.; Timmer, V.R. Modeling nitrogen and phosphorous interactions in intensively managed nursery soli-plant systems. Can. J. Soil Sci. 1996, 76, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Folk, R.S.; Grossnickle, S.C. Stock-type patterns of phosphorus uptake, retranslocation, net photosynthesis and morphological development in interior spruce. New For. 2000, 19, 27–49. [Google Scholar] [CrossRef]
- Zerihun, A.; Montagu, K.D. Belowground to aboveground biomass ratio and vertical root distribution responses of mature Pinus radiata stands to phosphorus fertilization at planting. Can. J. For. Res. 2004, 34, 1883–1894. [Google Scholar] [CrossRef]
- Lin, S.; Litaker, R.W.; Sunda, W.G. Phosphorus physiological ecology and molecular mechanisms in marine phytoplankton. J. Phycol. 2016, 52, 10–36. [Google Scholar] [CrossRef]
- Razaq, M.; Zhang, P.; Shen, H.L. Influence of nitrogen and phosphorous on the growth and root morphology of Acer mono. PLoS ONE 2017, 12, e0171321. [Google Scholar] [CrossRef] [Green Version]
- Frajese, G.V.; Benvenuto, M.; Fantini, M.; Ambrosin, E.; Sacchetti, P.; Masuelli, L.; Giganti, M.G.; Modesti, A.; Bei, R. Potassium increases the antitumor effects of ascorbic acid in breast cancer cell lines in vitro. Oncol. Lett. 2016, 11, 4224–4234. [Google Scholar] [CrossRef] [Green Version]
- de Melo, E.A.S.C.; de Gonçalves, J.L.M.; Rocha, J.H.T.; Hakamada, R.E.; Bazani, J.H.; Wenzel, A.V.A.; Arthur, J.C., Jr.; Borges, J.S.; Malheiros, R.; de Lemos, C.C.Z.; et al. Responses of clonal eucalypt plantations to N, P and K fertilizer application in different edaphoclimatic conditions. Forests 2016, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.; Wang, X.; Zhang, D.; Wu, X.; Zhou, Z. Responses of Taxus chinensis and Phoebe chekiangensis seedlings to controlled-release fertilizer in various formulations and application rates. iForest Biogeosci. For. 2019, 12, 254. [Google Scholar] [CrossRef]
- Snowdon, P. Modeling Type 1 and Type 2 growth responses in plantations after application of fertilizer or other silvicultural treatments. For. Ecol. Manag. 2002, 163, 229–244. [Google Scholar] [CrossRef]
- Yu, H.; Liu, D.; Chen, G.; Wan, B.; Wang, S.; Yang, B. A neural network ensemble method for precision fertilization modeling. Math. Comput. Model. 2010, 51, 1375–1382. [Google Scholar] [CrossRef]
- Narita, Y. Four-dimensional energy spectrum for space–time structure of plasma turbulence. Nonlinear Process. Geophys. 2014, 21, 41–47. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Average Temperature (°C) | Average Precipitation (mm) |
|---|---|---|
| 3 | 16.7 | 98.1 |
| 4 | 20.3 | 71.5 |
| 5 | 25.5 | 156.2 |
| 6 | 26.4 | 214.3 |
| 7 | 29.1 | 173.3 |
| 8 | 28.3 | 301.5 |
| 9 | 27.0 | 124.3 |
| 10 | 21.5 | 61.2 |
| 11 | 15.2 | 130.5 |
| 12 | 10.6 | 25.1 |
| Number | Fertilization Treatment | N (g·plant−1) | P2O5 (g·plant−1) | KO2 (g·plant−1) |
|---|---|---|---|---|
| T1 | N0P0K0 | 0.000 | 0.000 | 0.000 |
| T2 | N0P2K2 | 0.000 | 0.133 | 0.356 |
| T3 | N1P2K2 | 0.266 | 0.133 | 0.356 |
| T4 | N2P0K2 | 0.532 | 0.000 | 0.356 |
| T5 | N2P1K2 | 0.532 | 0.067 | 0.356 |
| T6 | N2P2K2 | 0.532 | 0.133 | 0.356 |
| T7 | N2P3K2 | 0.532 | 0.200 | 0.356 |
| T8 | N2P2K0 | 0.532 | 0.133 | 0.000 |
| T9 | N2P2K1 | 0.532 | 0.133 | 0.178 |
| T10 | N2P2K3 | 0.532 | 0.133 | 0.534 |
| T11 | N3P2K2 | 0.798 | 0.133 | 0.356 |
| T12 | N1P1K2 | 0.266 | 0.067 | 0.356 |
| T13 | N1P2K1 | 0.266 | 0.133 | 0.178 |
| T14 | N2P1K1 | 0.532 | 0.067 | 0.178 |
| Fertilization Treatment | N (g·kg−1) | P (g·kg−1) | K (g·kg−1) |
|---|---|---|---|
| T1 | 10.88 | 3.16 | 27.11 |
| T2 | 14.60 | 4.32 | 48.38 |
| T3 | 13.84 | 3.74 | 39.52 |
| T4 | 12.62 | 2.56 | 23.15 |
| T5 | 15.84 | 3.97 | 53.81 |
| T6 | 11.34 | 3.39 | 44.55 |
| T7 | 12.49 | 3.33 | 25.58 |
| T8 | 13.77 | 2.81 | 23.24 |
| T9 | 16.33 | 4.05 | 36.73 |
| T10 | 13.55 | 3.15 | 50.71 |
| T11 | 17.48 | 4.68 | 40.70 |
| T12 | 13.43 | 3.67 | 38.75 |
| T13 | 14.02 | 4.00 | 37.48 |
| T14 | 15.25 | 2.98 | 31.85 |
| Indicator 1 | N | P | K |
|---|---|---|---|
| Height (cm) | 17.12 | 13.55 | 8.58 |
| Root collar diameter (mm) | 1.16 | 1.31 | 0.8 |
| Rb (g) | 1.9 | 1.3 | 1.66 |
| Sb (g) | 1.44 | 1.07 | 1.26 |
| Lb (g) | 1.27 | 0.8 | 1.08 |
| Tm (g) | 4.55 | 3.17 | 3.97 |
| HD | 3.38 | 2.58 | 1.84 |
| Sr | 1.52 | 0.49 | 1.74 |
| QI | 0.66 | 0.45 | 0.57 |
| Source of Variation | df | SS | MS | F Value | p-Value |
|---|---|---|---|---|---|
| N | 3 | 0.88 | 0.29 | 386.88 | <0.001 |
| P | 3 | 0.44 | 0.15 | 162.70 | <0.001 |
| K | 3 | 0.64 | 0.21 | 289.15 | <0.001 |
| N×P | 7 | 0.89 | 0.13 | 176.96 | <0.001 |
| N×K | 7 | 0.92 | 0.13 | 196.83 | <0.001 |
| P×K | 7 | 0.66 | 0.09 | 157.31 | <0.001 |
| N×P×K | 13 | 0.44 | 0.05 | 122.15 | <0.001 |
| Index | N | P2O5 | KO2 |
|---|---|---|---|
| Root biomass | 0.20 | −0.02 | 0.22 |
| Stem biomass | 0.19 | 0.09 | 0.01 |
| Leaves biomass | 0.30 * | 0.17 | 0.10 |
| Total biomass | 0.24 | 0.07 | 0.13 |
| Shoot–root ratio | −0.02 | 0.29 | −0.32 * |
| Height | −0.31 * | 0.50 ** | 0.20 |
| Root collar diameter | −0.15 | 0.44 ** | −0.06 |
| Height–diameter ratio | −0.23 | 0.13 | 0.35 * |
| QI 1 | 0.23 | 0.04 | 0.09 |
| Model | Nutrient | Fertilizer Response Equation | p | R2 | Maximum Rate (g·Plant−1) |
|---|---|---|---|---|---|
| Unitary | N | Y = 0.134 + 2.409N − 2.742N2 | 0.023 | 0.686 | 0.439 |
| P2O5 | Y = 0.376 + 5.309P − 23.314P2 | 0.147 | 0.448 | 0.114 | |
| K2O | Y = 0.270 + 2.931K − 5.128K2 | 0.037 | 0.641 | 0.286 | |
| Binary | N | Y = −0.202 + 2.359N + 4.402P − 2.297N2 − 13.089P2 − 2.648NP | 0.030 | 0.570 | 0.442 |
| P2O5 | 0.123 | ||||
| N | Y = −0.421 + 2.555N + 2.938K − 2.269N2 − 3.724K2 − 1.611NK | 0.025 | 0.582 | 0.458 | |
| K2O | 0.295 | ||||
| P2O5 | Y = −0.296 + 6.376P + 3.385K − 14.800P2 − 4.025K2 − 8.413PK | 0.110 | 0.442 | 0.136 | |
| K2O | 0.278 | ||||
| Ternary | N | Y = 0.284 + 0.936N + 0.076P + 0.594K − 1.776N2 − 4.786P2 − 2.623K2 + 3.348NP + 0.575NK + 1.442PK | 0.016 | 0.529 | 0.361 |
| P2O5 | 0.061 | ||||
| K2O | 0.246 |
| Item | N | P2O5 | K2O |
|---|---|---|---|
| Coding weighted average | 0.396 | 0.096 | 0.262 |
| Standard error | 0.148 | 0.064 | 0.116 |
| 95% confidence interval | 0.373~0.420 | 0.086~0.106 | 0.243~0.280 |
| Optimal fertilizer (g·plant−1) | 0.373~0.420 | 0.086~0.106 | 0.243~0.280 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.-J.; Wu, X.-H.; Grossnickle, S.C.; Chen, L.-H.; Yu, X.-X.; El-Kassaby, Y.A.; Feng, J.-L. Formula Fertilization Promotes Phoebe bournei Robust Seedling Cultivation. Forests 2020, 11, 781. https://doi.org/10.3390/f11070781
Yang Z-J, Wu X-H, Grossnickle SC, Chen L-H, Yu X-X, El-Kassaby YA, Feng J-L. Formula Fertilization Promotes Phoebe bournei Robust Seedling Cultivation. Forests. 2020; 11(7):781. https://doi.org/10.3390/f11070781
Chicago/Turabian StyleYang, Zhi-Jian, Xiao-Hui Wu, Steven C. Grossnickle, Lai-He Chen, Xin-Xiao Yu, Yousry A. El-Kassaby, and Jin-Ling Feng. 2020. "Formula Fertilization Promotes Phoebe bournei Robust Seedling Cultivation" Forests 11, no. 7: 781. https://doi.org/10.3390/f11070781