An Assessment of Storability of Norway Spruce Container Seedlings in Freezer Storage as Affected by Short-Day Treatment


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
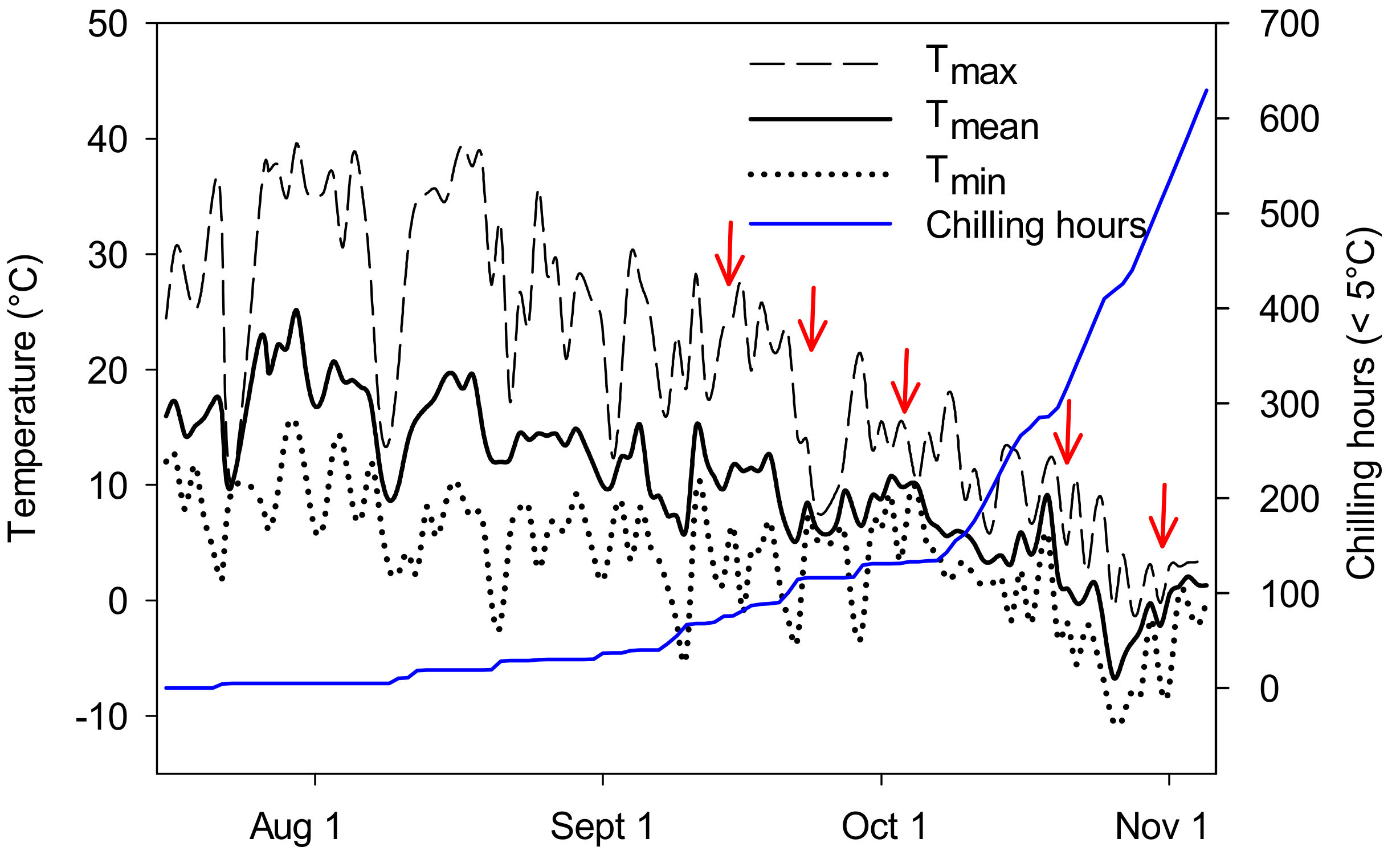
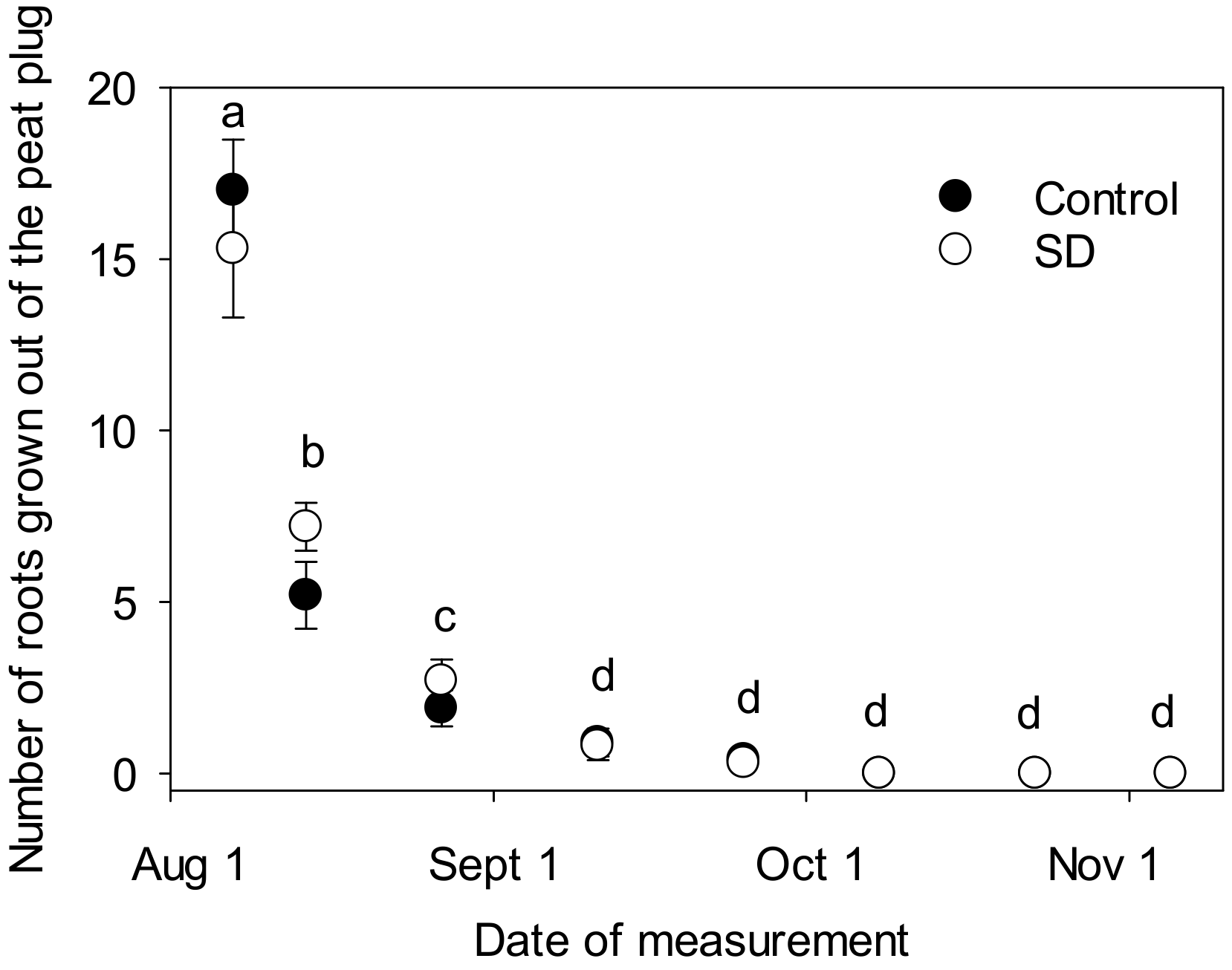
2.2. Determination of Root Growth in Autumn
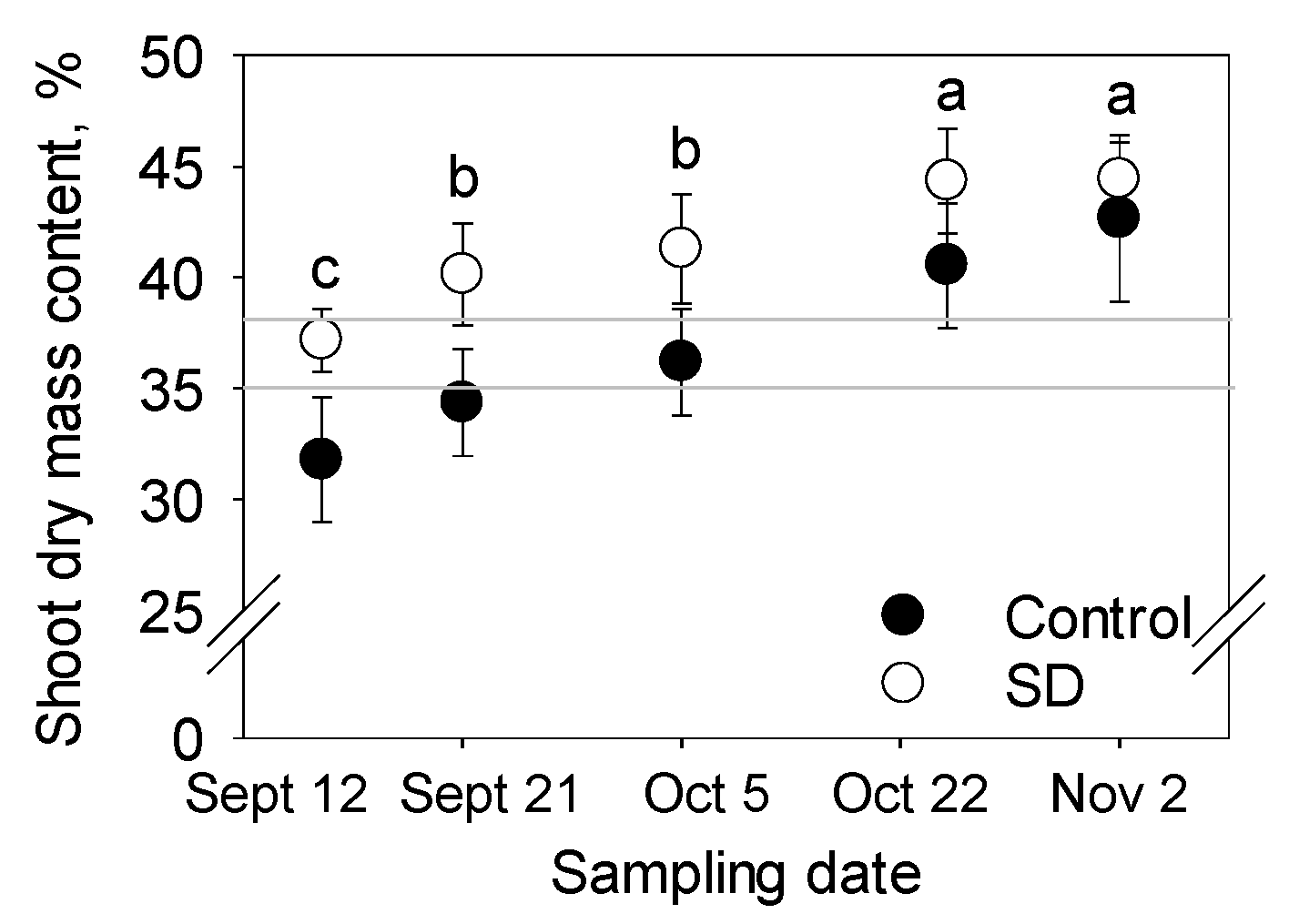
2.3. Storing of the Seedlings and Determination of Shoot Dry Matter Content
2.4. Frost Hardiness of Different Plant Parts
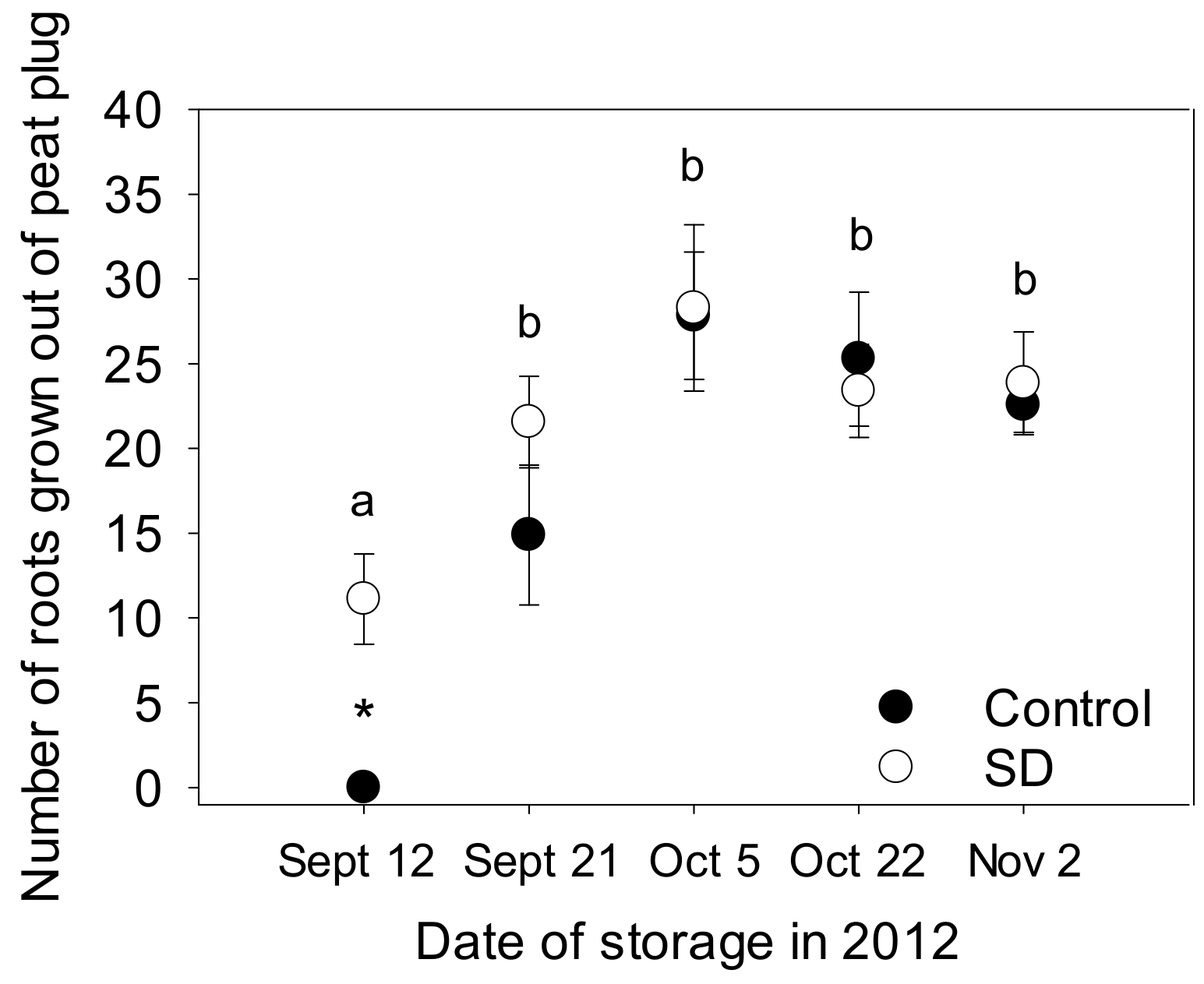
2.5. Vitality of Seedlings after Freezer Storage
2.6. Relationship between FH of Needles and Accumulation of Chilling Hours—Combined Data from Five Studies
2.7. Statistical Analysis
3. Results
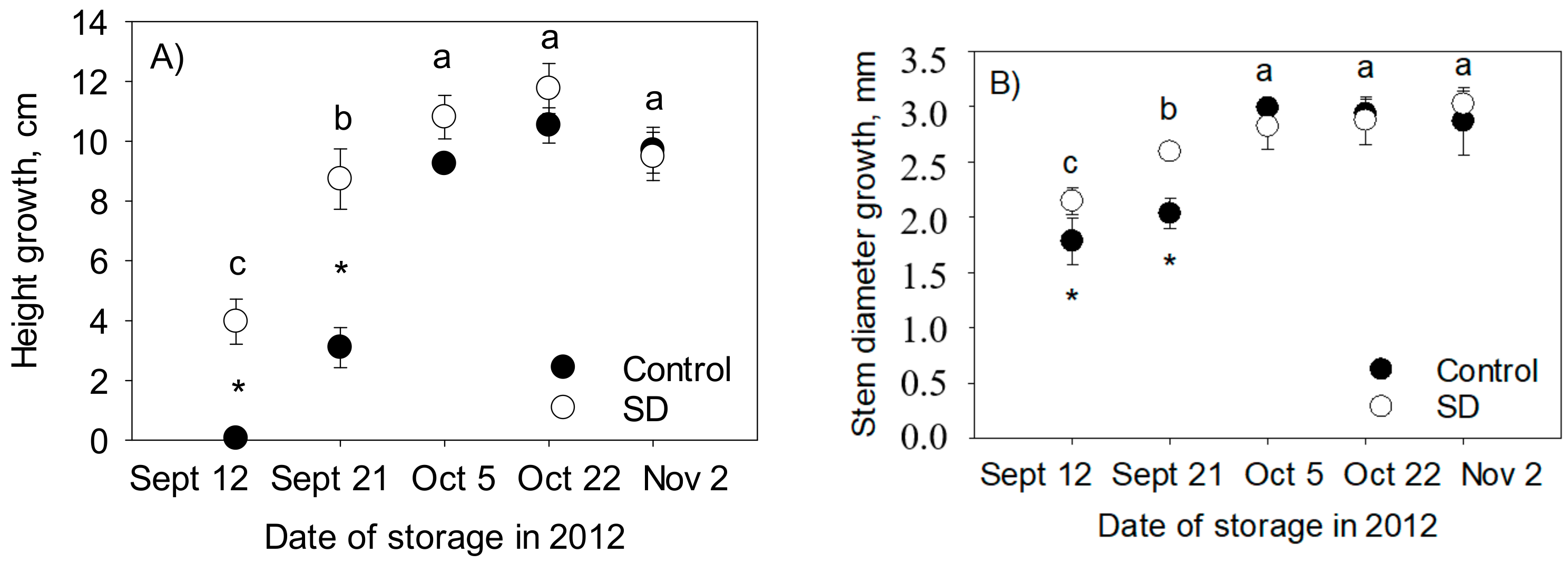
3.1. Effect of SD Treatment on Root Growth of Seedlings in Autumn
3.2. Dry Matter Content of Shoots and Frost Hardiness of Different Plant Parts
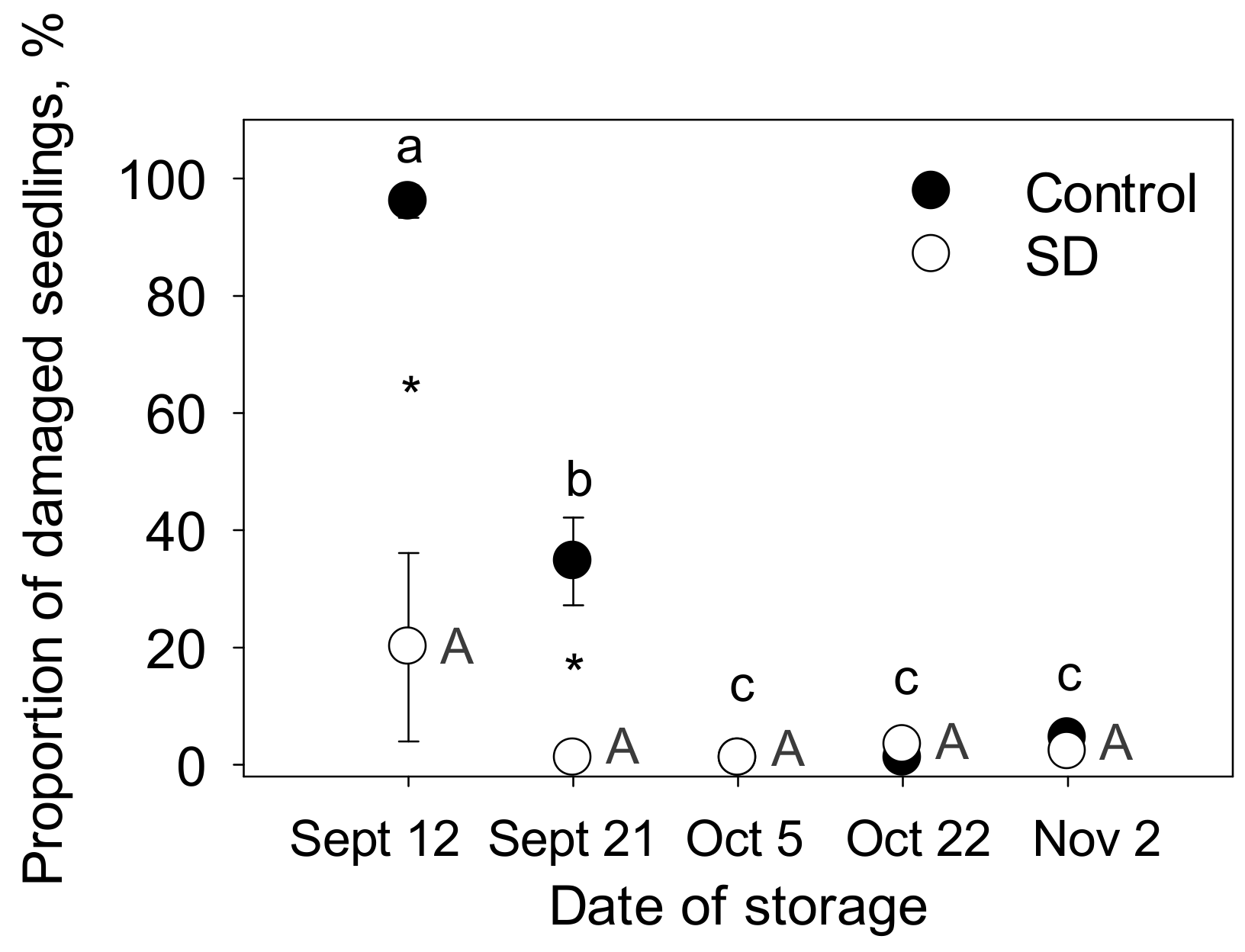
3.3. Vitality of the Seedlings after Freezer Storage
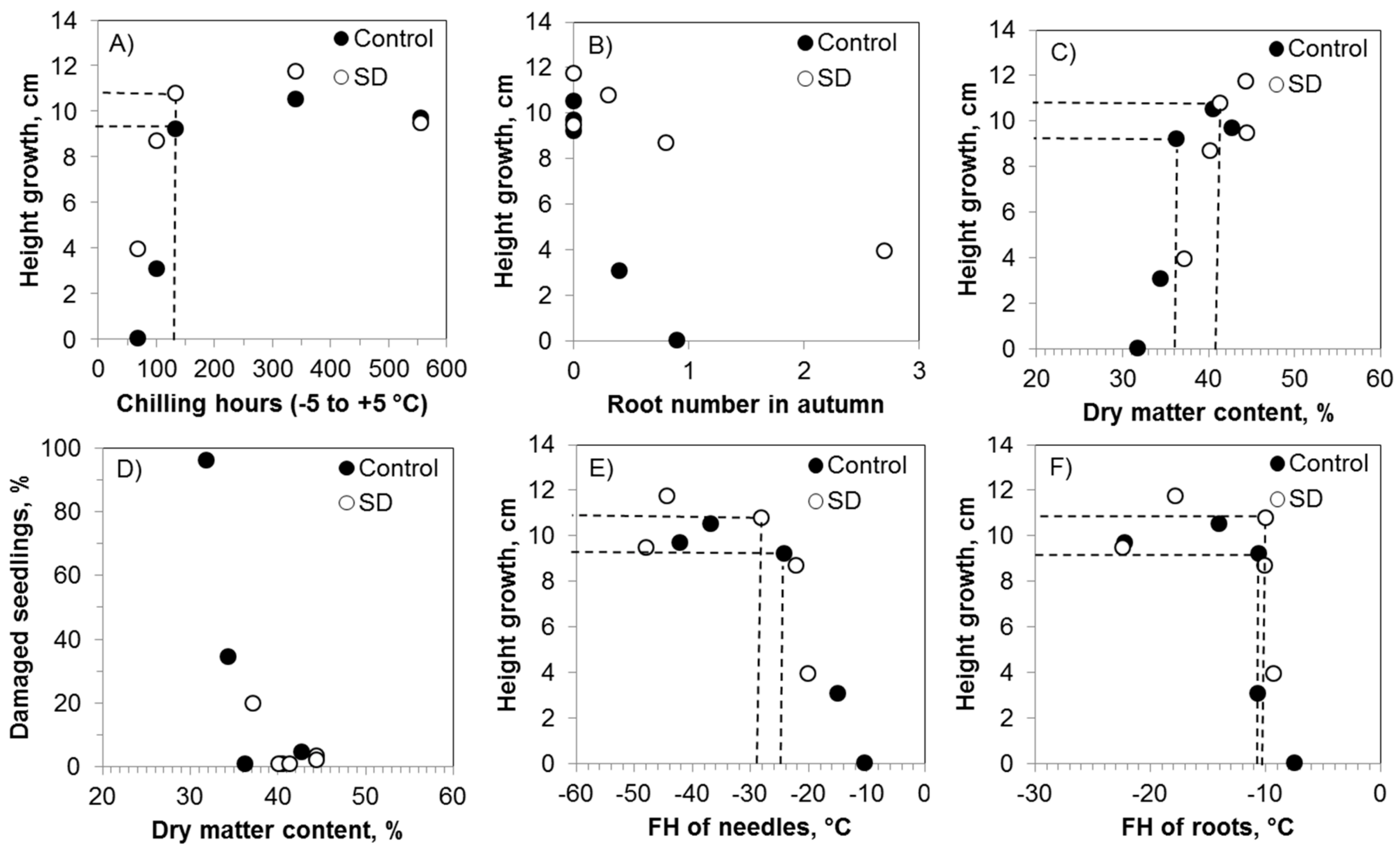
3.4. Variables Predicting Seedling Storability
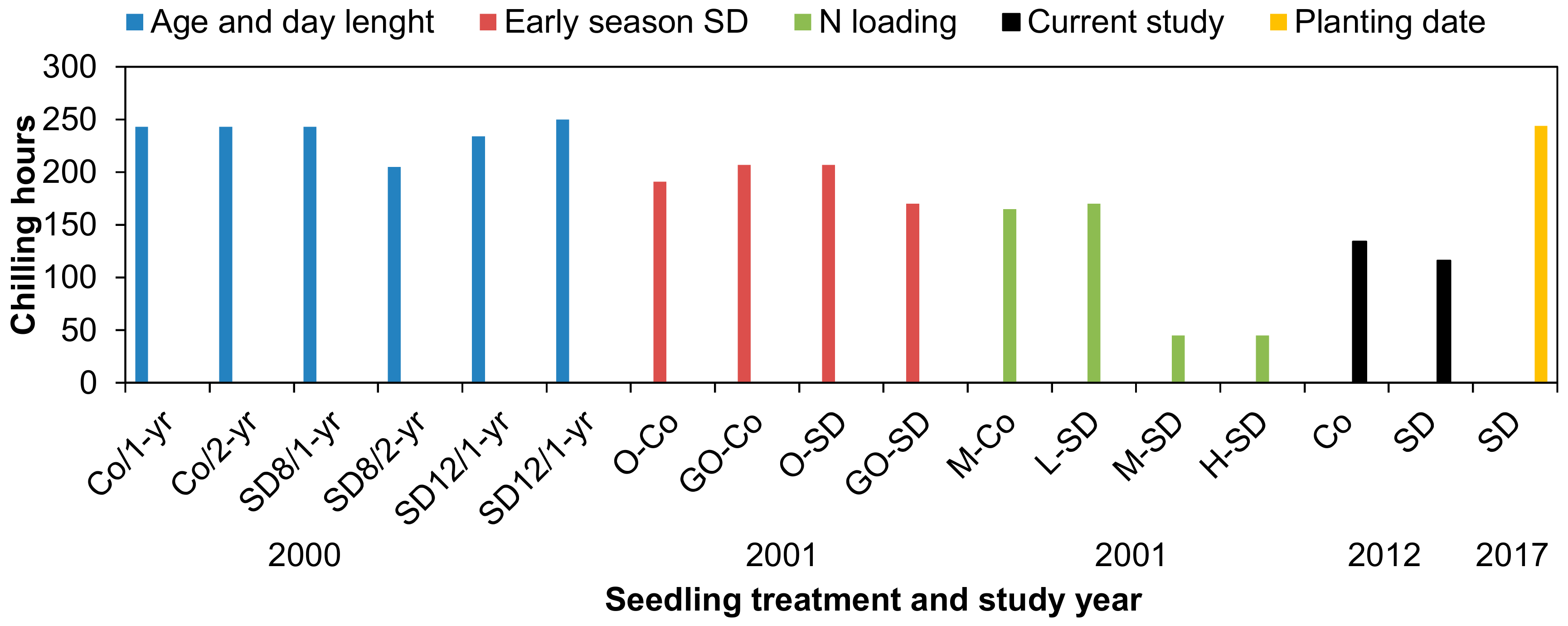
3.5. Relationship between Chilling Hours and FH of Needles
4. Discussion
4.1. Field Performance of Seedlings
4.2. Frost Hardiness of Seedlings
4.3. Shoot Dry Mass Content as an Indicator of Storability
4.4. Do Chilling Hours Indicate Storability of Seedlings?
4.5. Storage Duration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Landis, T.D.; Dumroese, R.K.; Haase, D.L. Seedling processing, storage, and outplanting. The container tree nursery manual. In Agricultural Handbook 674; Department of Agricultural Forest Service: Washington, DC, USA, 2010; Volume 7, p. 199. [Google Scholar]
- IPCC. Climate Change 2013: The Physical Science Basis; Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; p. 1535. [Google Scholar]
- Landis, T.D.; Tinus, R.W.; McDonald, S.E.; Barnett, J.P. Atmospheric environment. The container tree nursery manual. In Agricultural Handbook 674; Department of Agricultural Forest Service: Washington, DC, USA, 1992; Volume 3, p. 145. [Google Scholar]
- Bigras, F.J.; Ryyppö, A.; Lindström, A.; Stattin, E. Cold acclimation and deacclimation of shoots and roots of conifer seedlings. In Conifer Cold Hardiness; Bigras, F.J., Colombo, S.J., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2001; pp. 57–88. [Google Scholar]
- Lyr, H.; Hoffmann, G. Growth rates and growth periodicity of tree roots. Int. Rev. For. Res. 1967, 2, 181–206. [Google Scholar]
- Colombo, S.J.; Zhao, S.; Blumwald, E. Frost hardiness gradients in shoots and roots of Picea mariana seedlings. Scand. J. For. Res. 1995, 10, 32–36. [Google Scholar] [CrossRef]
- Lindström, A. Freezing temperatures in the root zone—Effects on growth of containerized Scots pine and Norway spruce seedlings. Scand. J. For. Res. 1986, 1, 371–377. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; South, D.B. Fall Acclimation and the Lift/Store Pathway: Effect on Reforestation. Open For. Sci. J. 2014, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Lindström, A.; Stattin, E.; Gräns, D.; Wallin, E. Storability measures of Norway spruce and Scots pine seedlings and assessment of post-storage vitality by measuring shoot electrolyte leakage. Scand. J. For. Res. 2014, 29, 717–724. [Google Scholar] [CrossRef]
- Lindström, A.; Håkansson, L. The EC-Method—A Way to Assess Storability of Forest Tree Seedlings; Research Report 95; Swedish University of Agricultural Sciences, Department of Forest Yield: Garpenberg, Sweden, 1996. [Google Scholar]
- Stattin, E.; Verhoef, N.; Balk, P.; van Wordragen, M.; Lindström, A. Development of a molecular test to determine the vitality status of Norway spruce (Picea abies) seedlings during frozen storage. New For. 2012, 43, 665–678. [Google Scholar] [CrossRef]
- Simpson, D.G. Frost hardiness, root growth capacity, and field performance relationships in interior spruce, lodgepole pine, Douglas-fir, and western hemlock seedlings. Can. J. For. Res. 1990, 20, 566–572. [Google Scholar] [CrossRef]
- L’Hirondelle, S.J.; Simpson, D.G.; Binder, W.D. Overwinter storability of conifer planting stock: Operational testing of fall frost hardiness. New For. 2006, 32, 307–321. [Google Scholar] [CrossRef]
- Mattsson, A. Seasonal variation in root growth capacity during cultivation of container grown Pinus sylvestris. Scand. J. For. Res. 1986, 1, 473–482. [Google Scholar] [CrossRef]
- Balk, P.A.; Brönnum, P.; Perks, M.; Stattin, E.; van der Geest, L.J.M.; van Wordragen, M. Innovative cold tolerance test for conifer seedlings. In Proceedings of the USDA Forest Service Proceedings RMRS-P-50, Fort Collins, CO, USA, 13–15 February 2007; pp. 9–14. [Google Scholar]
- Levitt, J. Responses of Plants to Environmental Stress: Chilling, Freezing, and High Temperature Stress; Academic Press: New York, NY, USA, 1980; Volume 1, p. 510. [Google Scholar]
- Colombo, S.J. Bud dormancy status, frost hardiness, shoot moisture content, and readiness of black spruce container seedlings for frozen storage. J. Am. Soc. Hortic. Sci. 1990, 115, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Stattin, E.; Hellqvist, C.; Lindström, A. 2000. Storability and root freezing tolerance of Norway spruce seedlings. Can. J. For. Res. 2000, 30, 964–970. [Google Scholar] [CrossRef]
- Lindström, A. How can we minimize storage damage. In Seedling Production and Seedling Damage—Conference Held in Uppsala, Sweden 30 January 1996; Hannerz, M., Ed.; The Forestry Research Institute of Sweden: Uppsala, Sweden, 1996; pp. 34–44. [Google Scholar]
- Rosvall-Åhnebrink, G. Preparing seedlings for autumn planting or winter storage. In Silviculture in Southern Sweden Jönköping 16–17 January 1985; Eriksson, A., Zimmerman, J., Eds.; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1985; pp. 33–37. [Google Scholar]
- Wallin, E.; Gräns, D.; Jacobs, D.F.; Lindström, A.; Verhoef, N. Short-day photoperiods affect expression of genes related to dormancy and freezing tolerance in Norway spruce seedlings. Ann. For. Sci. 2017, 74, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Juntunen, M.L.; Rikala, R. Fertilization practice in Finnish forest nurseries from the standpoint of environmental impact. New For. 2001, 21, 141–158. [Google Scholar] [CrossRef]
- Konttinen, K.; Luoranen, J.; Rikala, R. Growth and frost hardening of Picea abies seedlings after various night length treatments. Balt. For. 2007, 13, 140–148. [Google Scholar]
- Luoranen, J.; Lahti, M.; Rikala, R. Frost hardiness of nutrient-loaded two-year-old Picea abies seedlings in autumn and at the end of freezer storage. New For. 2008, 35, 207–220. [Google Scholar] [CrossRef]
- Luoranen, J.; Konttinen, K.; Rikala, R. Frost hardening and risk of a second flush in Norway spruce seedlings after an early-season short-day treatment. Silva. Fenn. 2009, 43, 209. [Google Scholar] [CrossRef] [Green Version]
- Luoranen, J.; Pikkarainen, L.; Poteri, M.; Peltola, H.; Riikonen, J. Duration limits on field storage in closed cardboard boxes before planting of Norway spruce and Scots pine container seedlings in different planting seasons. Forests 2019, 10, 1126. [Google Scholar] [CrossRef] [Green Version]
- SAS. Usage Note 56992: Modeling Continuous Proportions: Fractional and 4- (or 5-) Parameter Logit Models; SAS Institute Inc.: Cary, NC, USA, 2015; Available online: http://support.sas.com/kb/56/992.html (accessed on 20 May 2019).
- SAS. Usage Note 56549: Poisson Regression Using a Generalized Poisson Distribution for Overdispersed Data; SAS Institute Inc.: Cary, NC, USA, 2015; Available online: http://support.sas.com/kb/56/549.html (accessed on 28 November 2019).
- Lappi, J.; Luoranen, J. Testing the differences of LT50, LD50, or ED50. Can. J. For. Res. 2018, 48, 729–734. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Ecophysiology of Northern Spruce Species: The Performance of Planted Seedlings; NRC Research Press: Ottawa, ON, Canada, 2000; p. 409. [Google Scholar]
- Colombo, S.J.; Glerum, C.; Webb, D.P. Winter hardening in first-year black spruce (Picea mariana) seedlings. Physiol. Plant 1989, 76, 1–9. [Google Scholar] [CrossRef]
- Lindström, A.; Stattin, E. Root freezing tolerance and vitality of Norway spruce and Scots pine seedlings; influence of storage duration, storage temperature, and prestorage root freezing. Can. J. For. Res. 1994, 24, 2477–2484. [Google Scholar] [CrossRef]
- Räisänen, M.; Repo, T.; Rikala, R.; Lehto, T. Effect of thawing time, cooling rate and Boron nutrition on freezing point of the primordial shoot in Norway spruce buds. Ann. Bot. 2006, 97, 593–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormling, I.; Gustavsson, Å.; von Wettstein, D. The experimental control of the life cycle in Picea abies (L.) Karst. 1. Some basic experiments on the vegetative cycle. Silvae Genet. 1968, 17, 44–64. [Google Scholar]
- Welling, A.; Palva, E.T. Involvement of CBF transcription factors in winter hardiness in birch. Physiol. Plant 2006, 127, 167–181. [Google Scholar] [CrossRef]
- Ögren, E. Relationship between temperature, respiratory loss of sugar and premature dehardening in dormant Scots pine seedlings. Tree Phys. 1997, 17, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, G.A. Carbohydrate reserves and root growth potential in Douglas fir seedlings before and after cold storage. Can. J. For. Res. 1982, 12, 905–912. [Google Scholar] [CrossRef]
- Luoranen, J.; Riikonen, J.; Rikala, R.; Sutinen, S. Frost hardiness, carbohydrates and bud morphology of Picea abies seedlings after different lengths of freezer storage. Scand. J. For. Res. 2012, 27, 414–419. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Chilling Hours | Test Temperatures (°C) | |
|---|---|---|---|
| Shoots | Roots | ||
| 12 September (245 days) | 68 | 5, −4, −7, −9, −11, −13, −15, −20, and −25 | 5, −3, −5, −7, and −10 |
| 21 September (231 days) | 116 | 5, −5, −10, −15, −18, −21, −24, −30, and −40 | 5, −4, −6, −8, and −12 |
| 5 October (215 days) | 144 | 5, −10, −18, −22, −26, −30, −35, −40, and −55 | 5, −5, −9, −13, and −17 |
| 22 October (198 days) | 361 | 5, −15, −25, −30, −35, −39, −45, −49, and −70 | 5, −6, −10, −15, and −20 |
| 2 November (188 days) | 627 | 5, −20, −25, −30, −40, −45, −50, −60, and −70 | 5, −8, −13, −17, and −29 |
| Date of Freezing Test | ||||||
|---|---|---|---|---|---|---|
| Plant Part | Treatment | 12 September | 21 September | 5 October | 22 October | 2 November |
| Bud | Control | −8 (0.5) a* | −14 (0.5 ) b* | −15 (1.4) bc | −13 (2.0) b* | −19 (7.3) c |
| SD | −14 (0.6) a | −22 (0.8) bc | −14 (1.5) a | −19 (1.6) ac | −21 (5.9) ac | |
| Needle | Control | −11 (0.3) a* | −15 (0.4) b* | −24 (1.3) c* | −37 (2.4) d | −42 (0.8) d* |
| SD | −20 (1.0) a | −22 (0.6) a | −28 (1.5) b | −45 (4.1) c | −48 (1.1) c | |
| Stem | Control | −11 (0.9) a* | −20 (0.4) b* | −22 (3.0) b | −36 (1.3) c* | −47 (2.1) c* |
| SD | −20 (2.3) a | −26 (0.7) b | −24 (3.5) c | −49 (2.4) d | −59 (3.3) e | |
| Root | Control | −8 (0.6) a | −11 (1.0) a | −11 (0.9) a | −14 (1.1) b | −22 (2.3) c |
| SD | −9 (0.8) a | −10 (4.8) a | −10 (5.0) a | −18 (1.5) b* | −22 (2.4) b | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riikonen, J.; Luoranen, J. An Assessment of Storability of Norway Spruce Container Seedlings in Freezer Storage as Affected by Short-Day Treatment. Forests 2020, 11, 692. https://doi.org/10.3390/f11060692
Riikonen J, Luoranen J. An Assessment of Storability of Norway Spruce Container Seedlings in Freezer Storage as Affected by Short-Day Treatment. Forests. 2020; 11(6):692. https://doi.org/10.3390/f11060692
Chicago/Turabian StyleRiikonen, Johanna, and Jaana Luoranen. 2020. "An Assessment of Storability of Norway Spruce Container Seedlings in Freezer Storage as Affected by Short-Day Treatment" Forests 11, no. 6: 692. https://doi.org/10.3390/f11060692