A New Approach for Modeling Volume Response from Mid-Rotation Fertilization of Pinus taeda L. Plantations

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
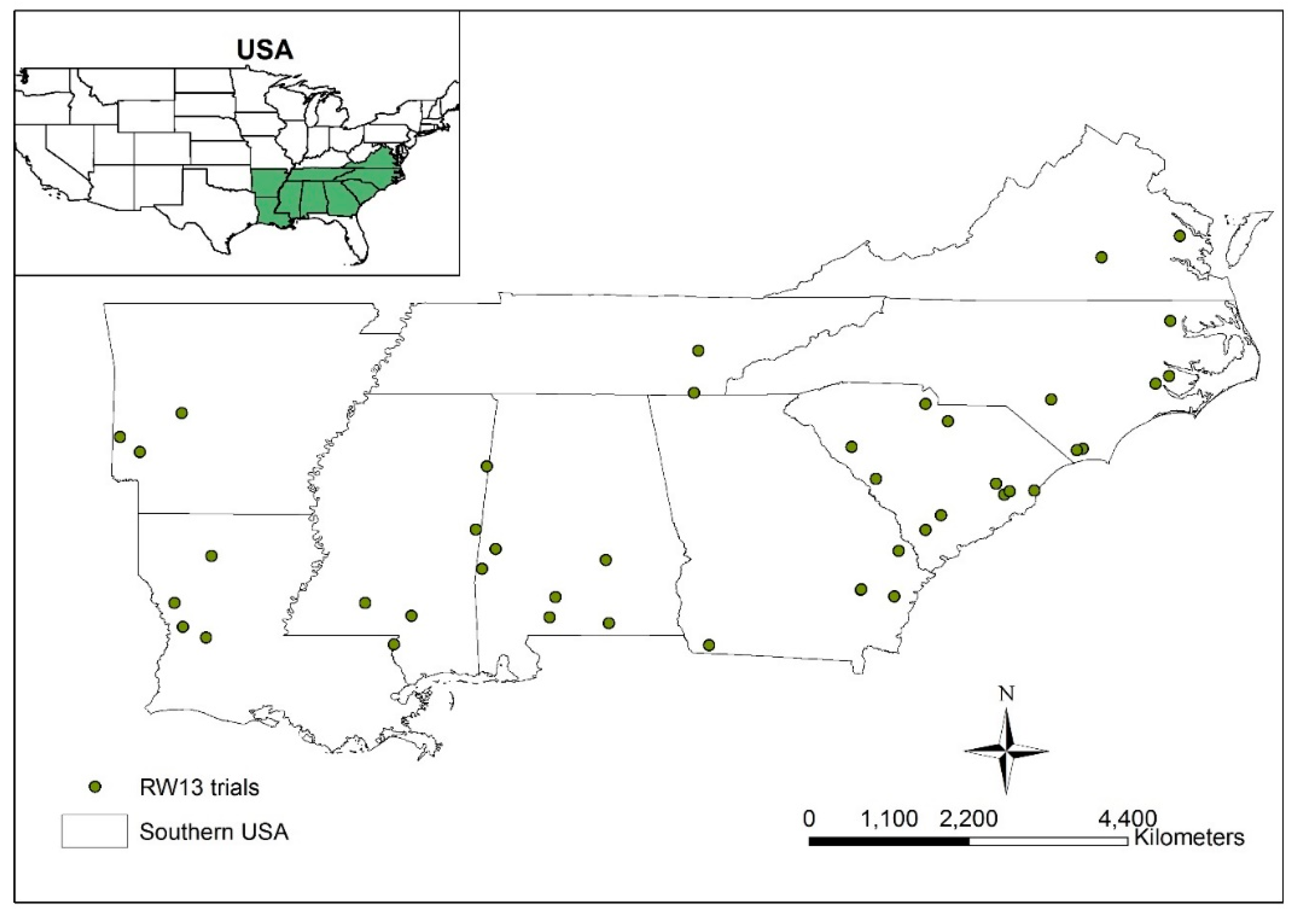
2.1. Study Area and Database Description
2.2. Model Rationale
2.2.1. Fitting Procedure
2.2.2. Data Standardization
2.2.3. Model Development
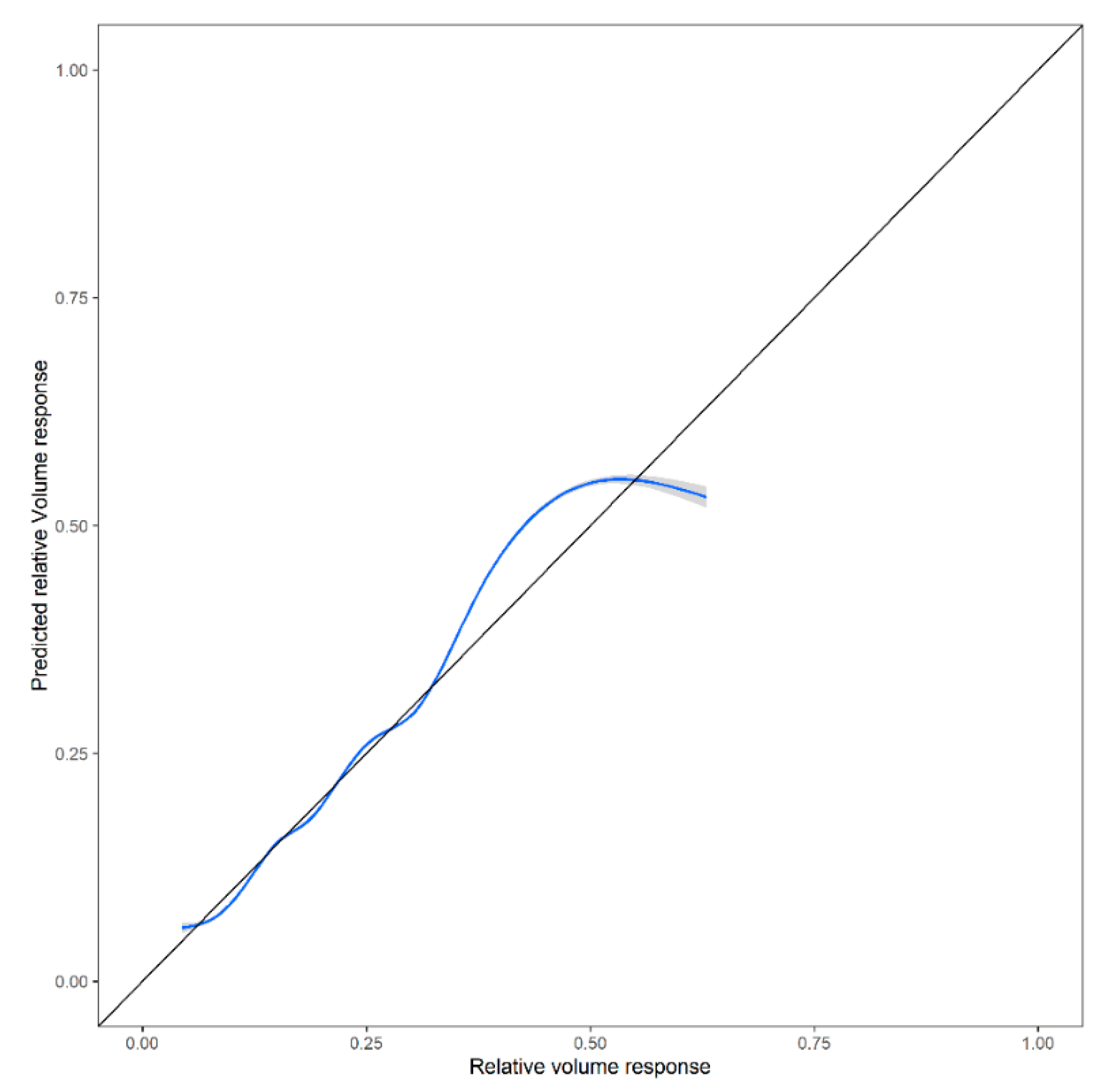
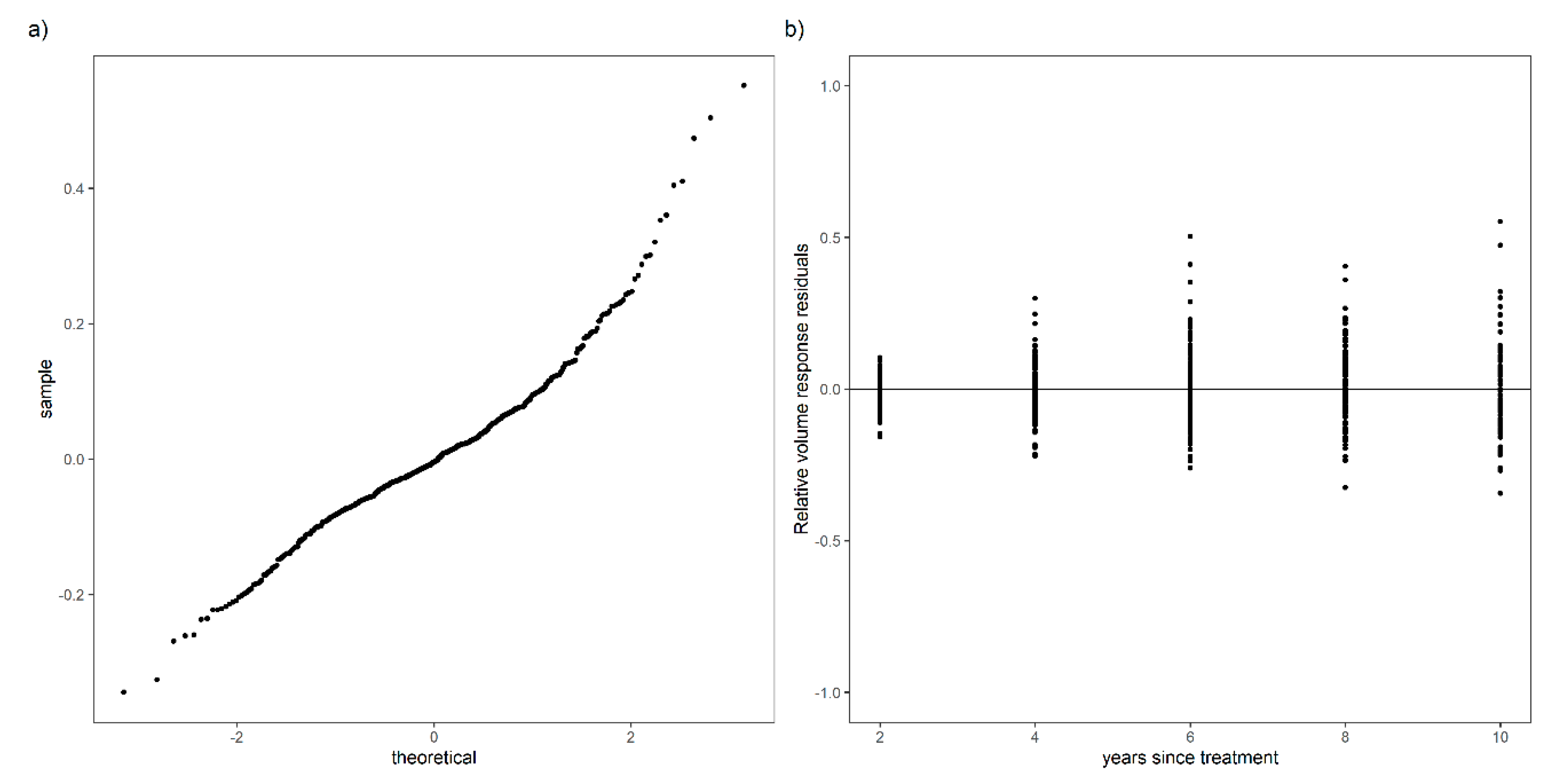
2.3. Validation
3. Results
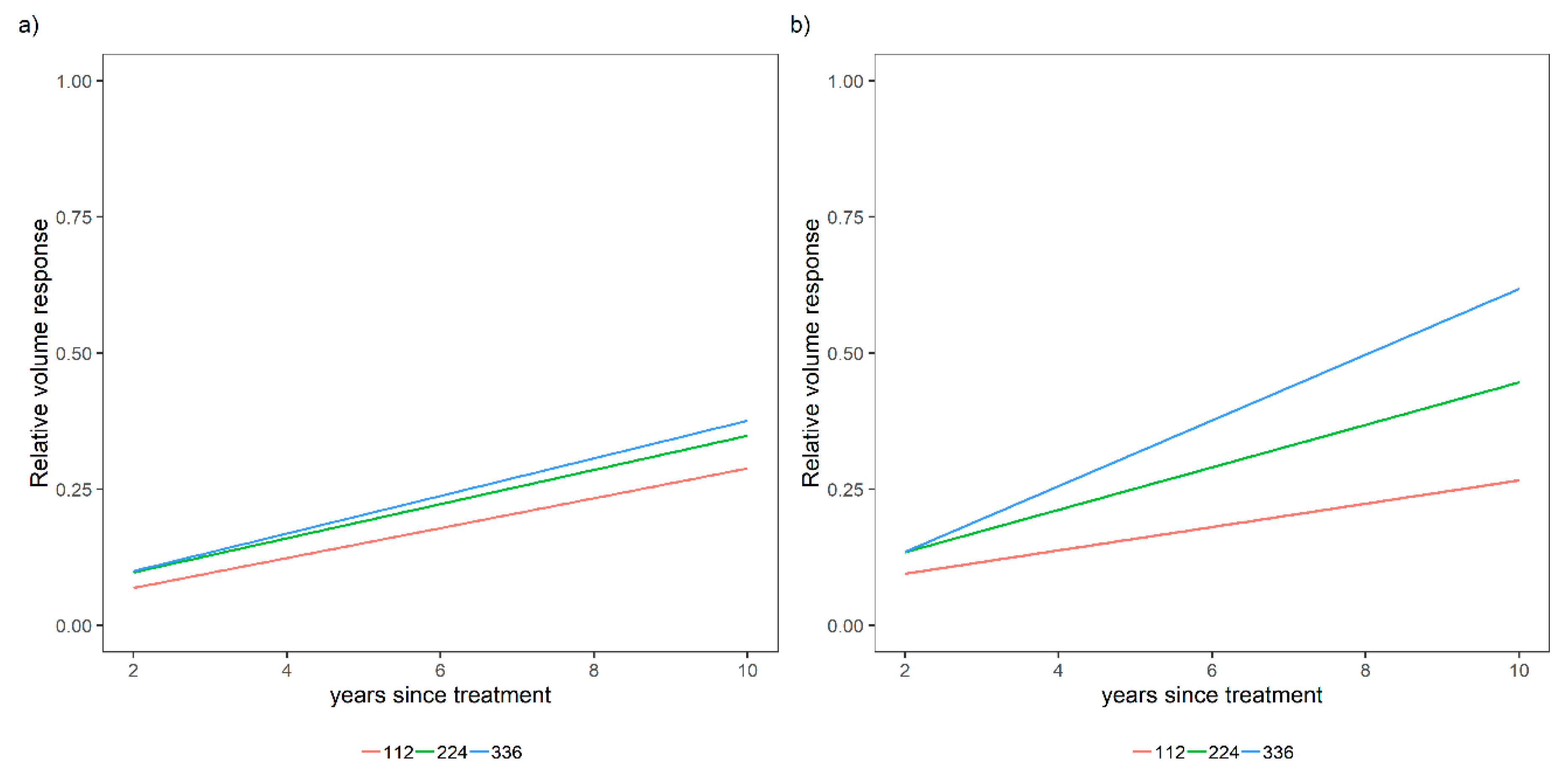
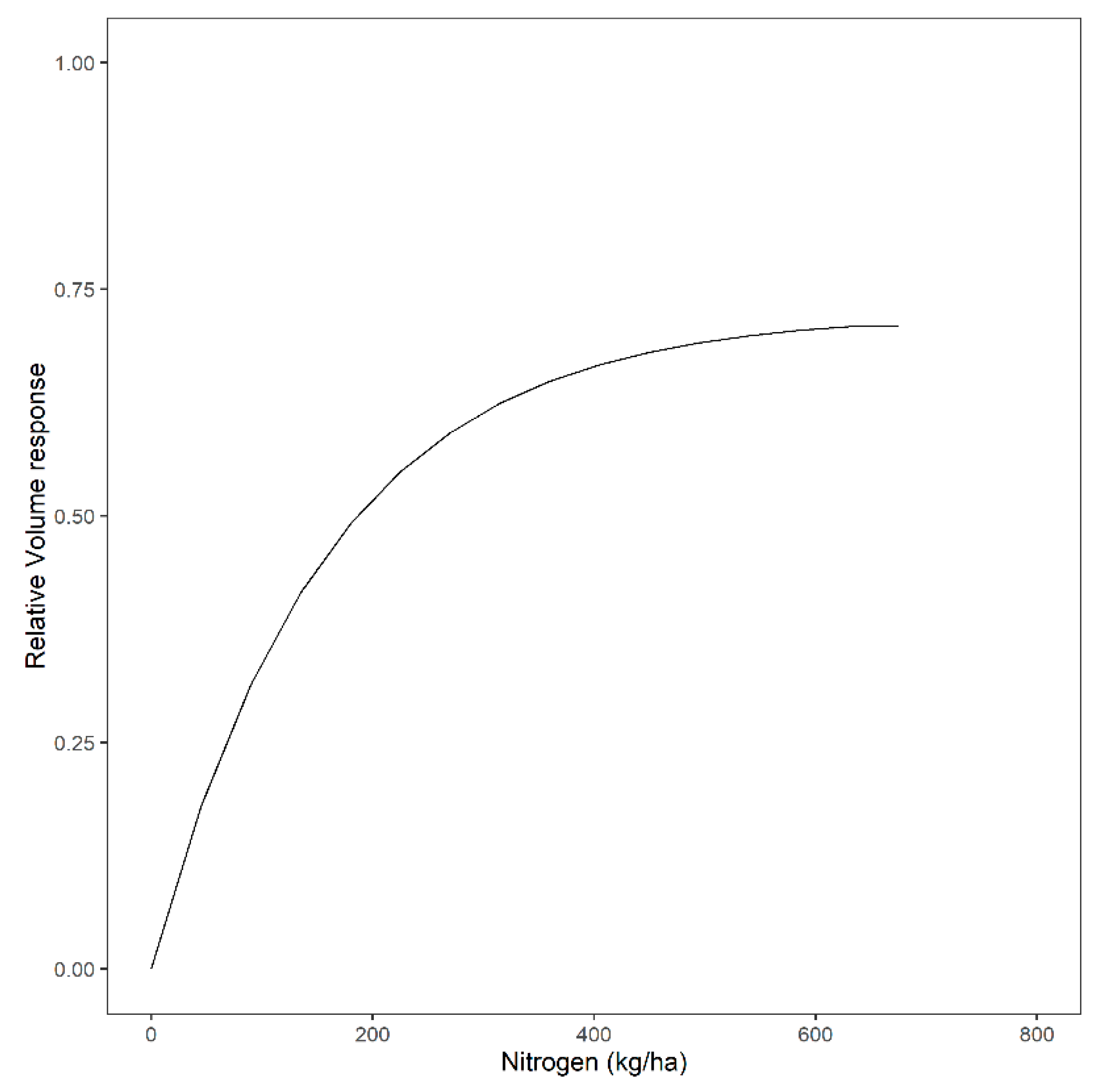
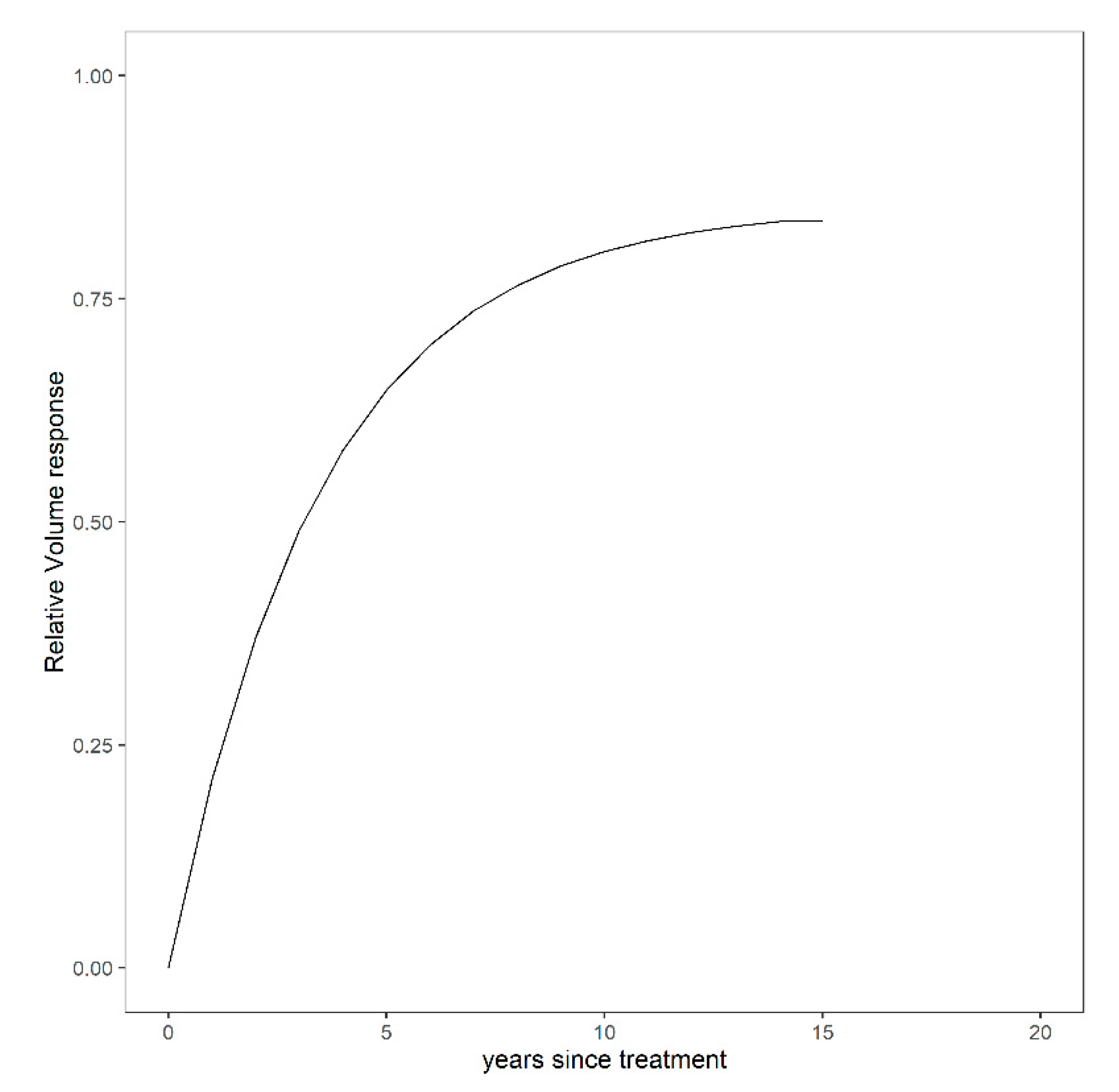
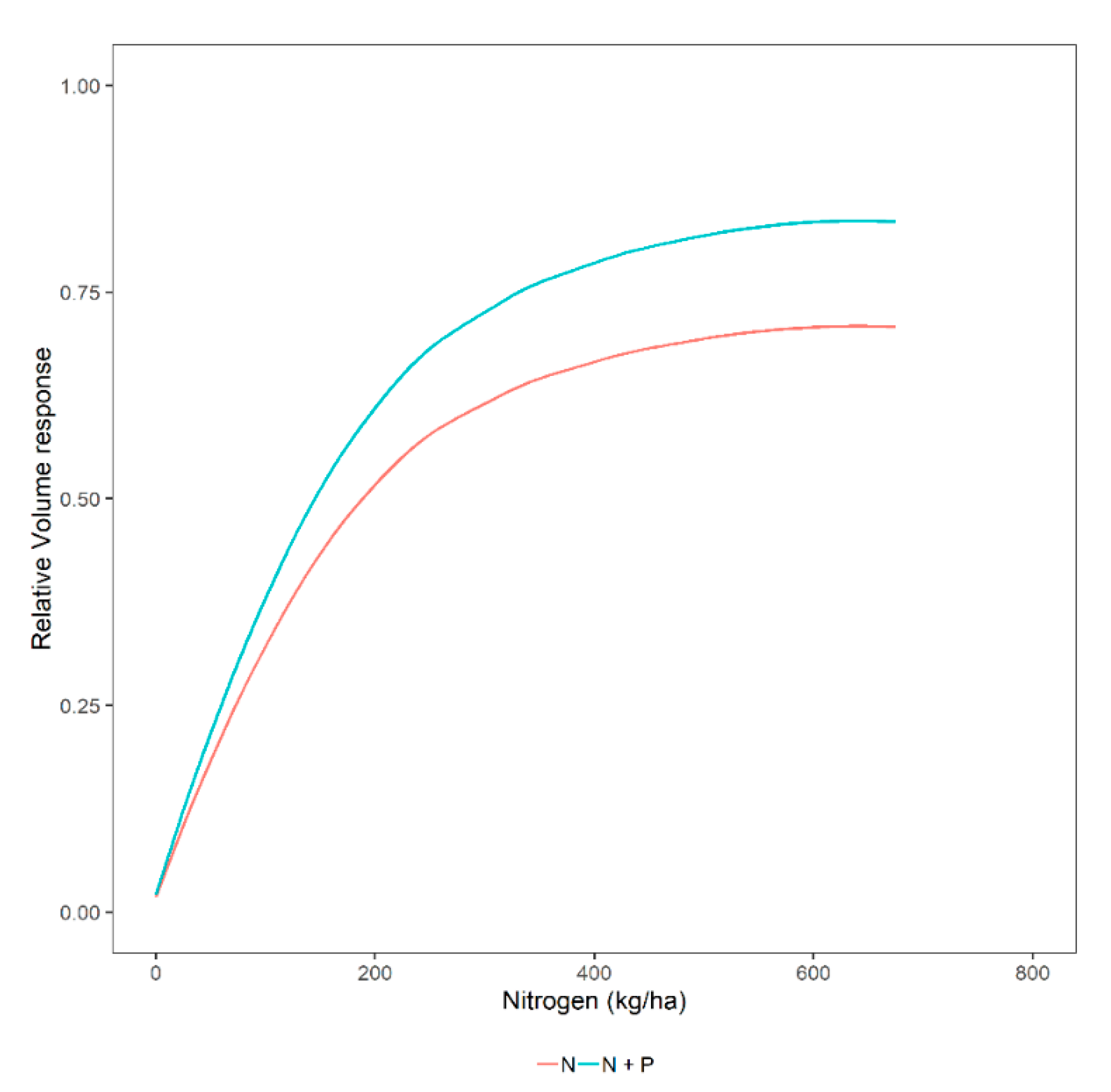
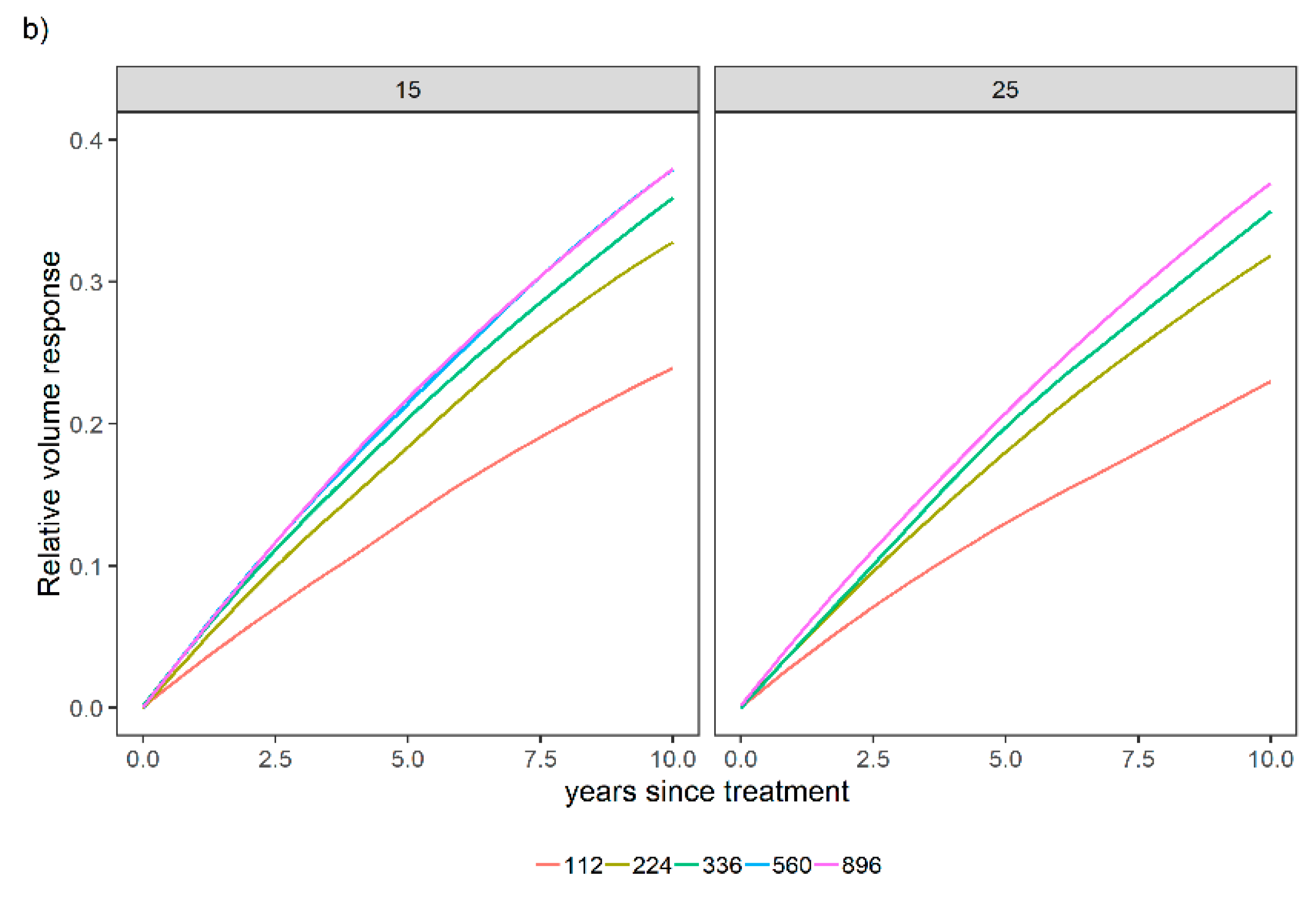
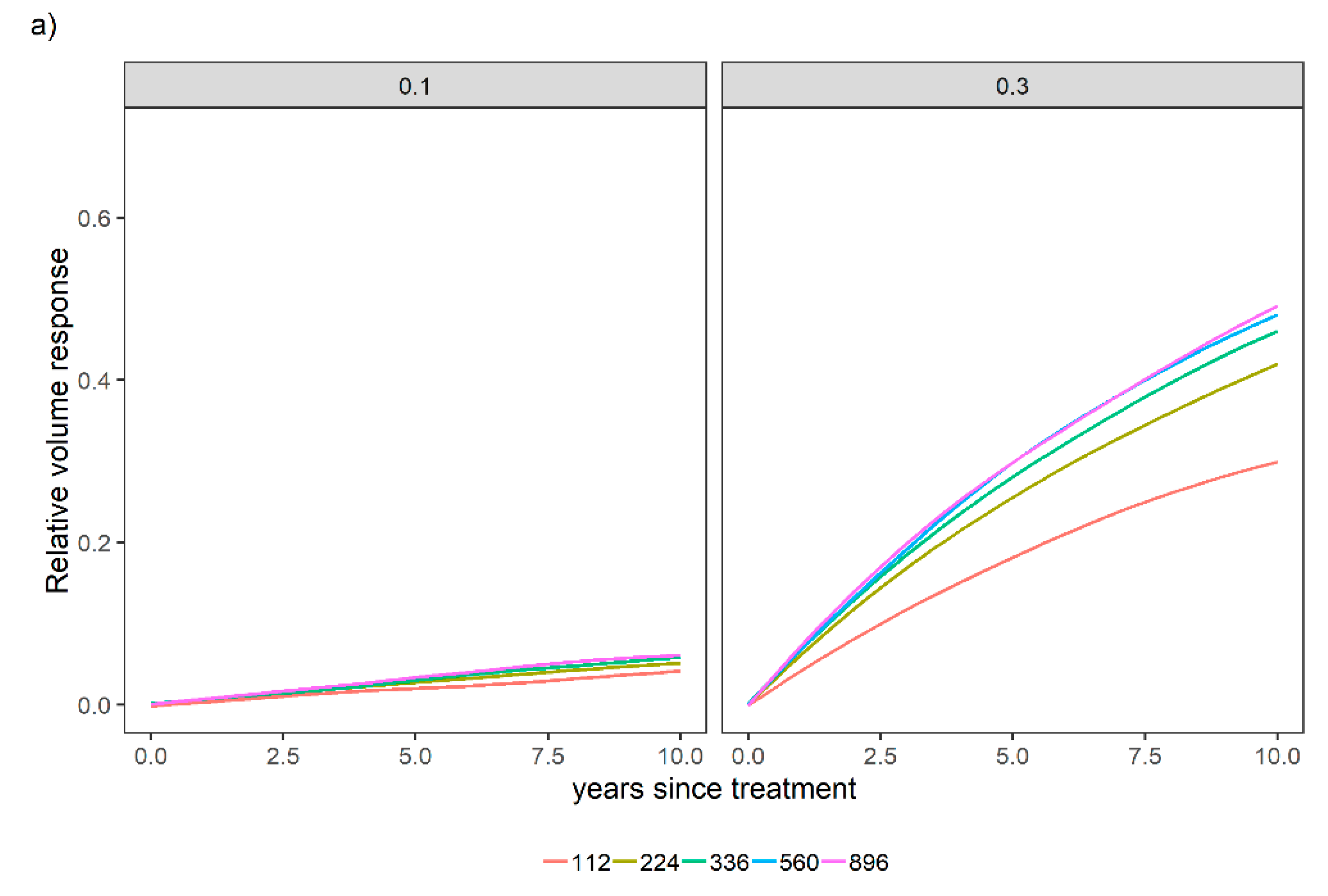
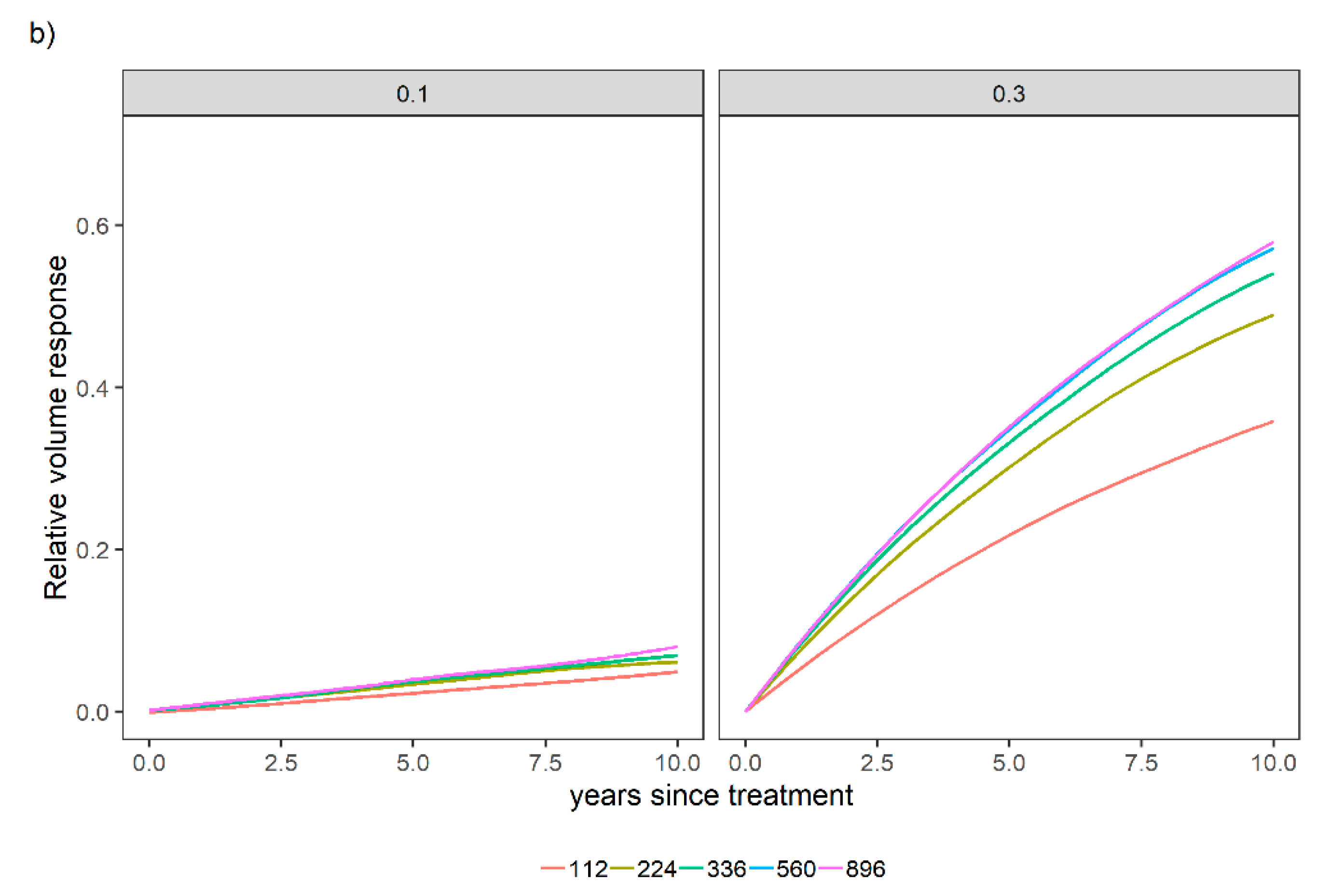
3.1. Overall Relative Volume Response to Different Fertilization Levels
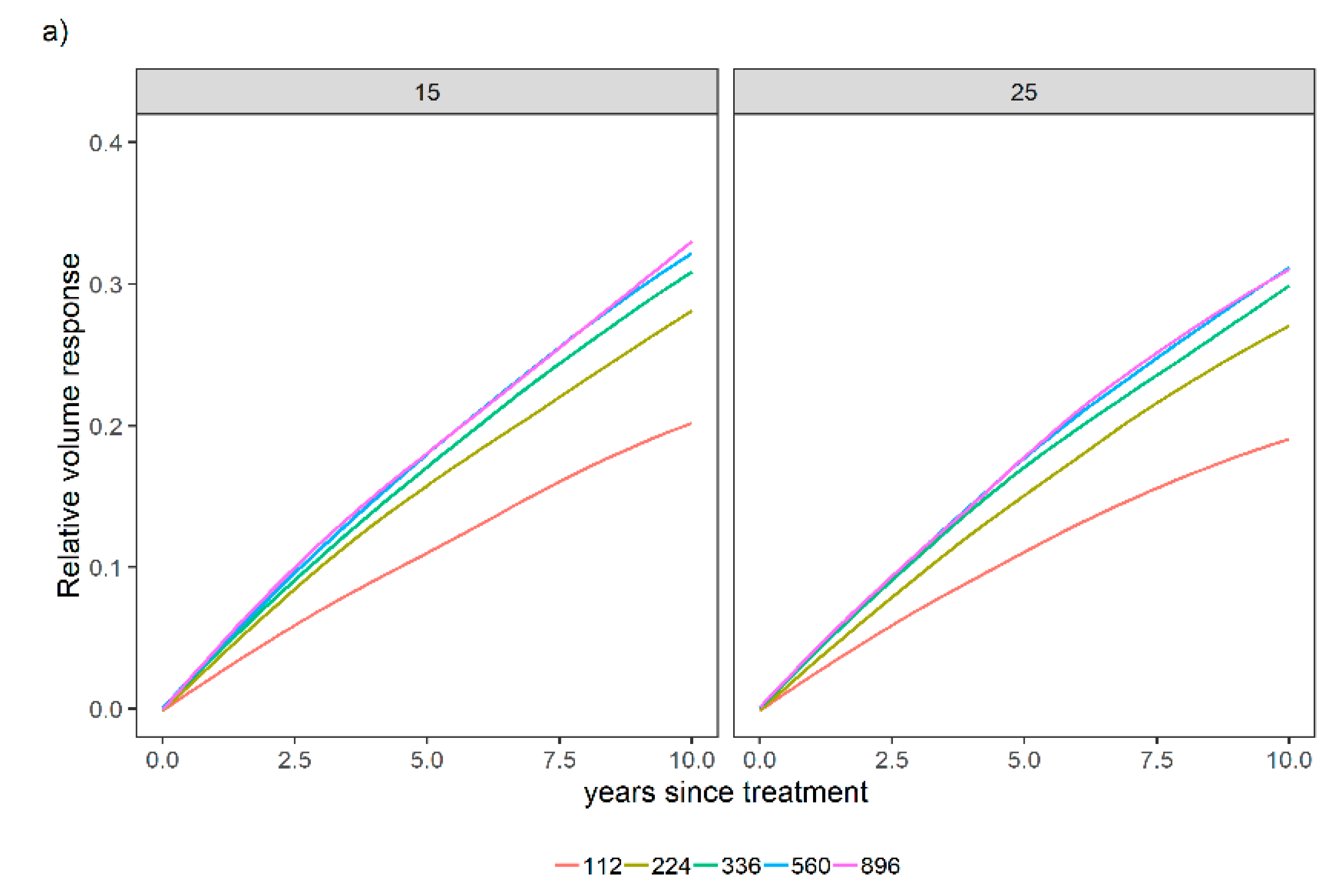
3.2. Sensitivity Analysis for a Variety of Relative Spacing and Site Index Scenarios over a Variety of N and P Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stape, J.-L.; Binkley, D.; Ryan, M.G.; Fonseca, S.; Loos, R.A.; Takahashi, E.N.; Silva, C.R.; Silva, S.R.; Hakamada, R.E.; Ferreira, J.M.D.A.; et al. The Brazil Eucalyptus Potential Productivity Project: Influence of water, nutrients and stand uniformity on wood production. For. Ecol. Manag. 2010, 259, 1684–1694. [Google Scholar] [CrossRef]
- Gyawali, N.; Burkhart, H.E. General response functions to silvicultural treatments in loblolly pine platations. Can. J. For. Res. 2015, 45, 252–265. [Google Scholar] [CrossRef]
- McTague, J.P.; O’Loughlin, D.; Roise, J.P.; Robison, D.J.; Kellison, R.C. The SOHARC Model System for Growth and Yield of Southern Hardwoods. South. J. Appl. For. 2008, 32, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Montes, C. A Resource Driven Growth and Yield Model for Loblolly Pine Plantations. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 2012. [Google Scholar]
- Scolforo, H.F.; McTague, J.P.; Burkhart, H.; Roise, J.; Campoe, O.; Stape, J.L. Eucalyptus growth and yield system: Linking individual-tree and stand-level growth models in clonal Eucalypt plantations in Brazil. For. Ecol. Manag. 2019, 432, 1–16. [Google Scholar] [CrossRef]
- Allen, H.; Dougherty, P.; Campbell, R. Manipulation of water and nutrients—Practice and opportunity in Southern U.S. pine forests. For. Ecol. Manag. 1990, 30, 437–453. [Google Scholar] [CrossRef]
- Ramírez, A.M.; Ra, R.; Montes, C.; Hl, A.; Tr, F.; Sanfuentes, E.; Alzate, M.R. Mid-rotation response to fertilizer by Pinus radiata D. Don at three contrasting sites. J. For. Sci. 2016, 62, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Fox, T.R.; Allen, H.L.; Albaugh, T.J.; Rubilar, R.; Carlson, C.A. Tree Nutrition and Forest Fertilization of Pine Plantations in the Southern United States. South. J. Appl. For. 2007, 31, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Albaugh, T.J.; Allen, H.L.; Fox, T.R. Historical Patterns of Forest Fertilization in the Southeastern United States from 1969 to 2004. South. J. Appl. For. 2007, 31, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Snowdon, P. Modeling Type 1 and Type 2 growth responses in plantations after application of fertilizer or other silvicultural treatments. For. Ecol. Manag. 2002, 163, 229–244. [Google Scholar] [CrossRef]
- Woollons, R.C.; Whyte, A.G.D.; Mead, D.J. Long-term growth responses in Pinus Radiata fertilizer experiments. N. Z. J. For. Sci. 1988, 18, 199–209. [Google Scholar]
- McInnis, L.M.; Oswald, B.P.; Williams, H.M.; Farrish, K.W.; Unger, D.R. Growth response of Pinus taeda L. to herbicide, prescribed fire, and fertilizer. For. Ecol. Manag. 2004, 199, 231–242. [Google Scholar] [CrossRef]
- Carlson, C.A.; Fox, T.R.; Allen, H.L.; Albaugh, T.J. Modeling mid-rotation fertilizer responses using the age-shift approach. For. Ecol. Manag. 2008, 256, 256–262. [Google Scholar] [CrossRef]
- Pienaar, L.; Rheney, J.W. Modeling stand level growth and yield response to silvicultural treatments. For. Sci. 1995, 41, 629–638. [Google Scholar]
- Amateis, R.; Burkhart, H.; Allen, H.L.; Montes, C. FASTLOB: Fertilized and Selectively Thinned Loblolly Pine Plantations (a Stand-Level Growth and Yield Model for Fertilized and Thinned Loblolly Pine Plantations). In Loblolly Pine Growth and Yield Cooperative; VPISU: Blacksburg, VA, USA, 2001. [Google Scholar]
- Rojas, J. Factors influencing responses of loblolly pine stands to fertilization. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 2005. [Google Scholar]
- Hökkä, H.; Repola, J.; Moilanen, M. Modelling volume growth response of young Scots pine (Pinus sylvestris) stands to N, P, and K fertilization in drained peatland sites in Finland. Can. J. For. Res. 2012, 42, 1359–1370. [Google Scholar] [CrossRef]
- Ingestad, T. Relative addition rate and external concentration; Driving variables used in plant nutrition research. Plant. Cell Environ. 1982, 5, 443–453. [Google Scholar] [CrossRef]
- Albaugh, T.J.; Fox, T.R.; Allen, H.L.; Rubilar, R. Juvenile Southern Pine Response to Fertilization Is Influenced by Soil Drainage and Texture. Forests 2015, 6, 2799–2819. [Google Scholar] [CrossRef] [Green Version]
- Mason, E.G.; Milne, P.G. Effects of weed control, fertilization, and soil cultivation on the growth of Pinus radiata at midrotation in Canterbury, New Zealand. Can. J. For. Res. 1999, 29, 985–992. [Google Scholar] [CrossRef]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org (accessed on 3 May 2019).
- Zhao, D.; Kane, M.; Borders, B.E. The Crown Ratio and Relative Spacing Relationships for Loblolly Pine Plantation Stands; PMRC Technical Report: Athens, GA, USA, 2009; p. 24. [Google Scholar]
- Kozak, A.; Kozak, R. Does cross validation provide additional information in the evaluation of regression models? Can. J. For. Res. 2003, 33, 976–987. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; p. 211. [Google Scholar]
- Miehle, P.; Battaglia, M.; Sands, P.J.; Forrester, D.I.; Feikema, P.M.; Livesley, S.; Morris, J.D.; Arndt, S. A comparison of four process-based models and a statistical regression model to predict growth of Eucalyptus globulus plantations. Ecol. Model. 2009, 220, 734–746. [Google Scholar] [CrossRef]
- Haines, L.W.; Haines, S.G.; Allen, V.H., Jr. Fertilizing established loblolly pine stands. In Proc. 6th S. Forest Soils Workshop; US Forest: Charleston, SC, USA, 1976; pp. 56–67. [Google Scholar]
- King, J.S.; Albaugh, T.J.; Allen, H.L.; Buford, M.; Strain, B.R.; Dougherty, P. Below-ground carbon input to soil is controlled by nutrient availability and fine root dynamics in loblolly pine. New Phytol. 2002, 154, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Fox, T.R. Sustained productivity in intensively managed forest plantations. For. Ecol. Manag. 2000, 138, 187–202. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | N (kg ha−1) | P (kg ha−1) |
|---|---|---|
| 1 | 0 | 0 |
| 2 | 112 | 0 |
| 3 | 112 | 28 |
| 4 | 224 | 0 |
| 5 | 224 | 28 |
| 6 | 336 | 0 |
| 7 | 336 | 28 |
| Pretreatment Site Characteristics | Minimum | Average | Maximum |
|---|---|---|---|
| Foliar N (%) | 0.87 | 1.05 | 1.34 |
| pH | 3.78 | 4.85 | 7.17 |
| Carbon (%) | 0.91 | 2.03 | 4.63 |
| Nitrogen (%) | 0.02 | 0.07 | 0.13 |
| Phosphorus (mg kg−1) | 1.04 | 5.29 | 17.50 |
| Inventory Variables | Minimum | Average | Maximum |
|---|---|---|---|
| Stand age (years) | 9.00 | 13.40 | 19.00 |
| TPH | 551 | 1142 | 2059 |
| H (m) | 7.91 | 12.58 | 17.51 |
| S (m) | 16.64 | 20.13 | 24.96 |
| B (m2 ha−1) | 11.97 | 20.83 | 29.65 |
| V (m3 ha−1) | 44.86 | 120.73 | 198.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scolforo, H.F.; Montes, C.; Cook, R.L.; Lee Allen, H.; Albaugh, T.J.; Rubilar, R.; Campoe, O. A New Approach for Modeling Volume Response from Mid-Rotation Fertilization of Pinus taeda L. Plantations. Forests 2020, 11, 646. https://doi.org/10.3390/f11060646
Scolforo HF, Montes C, Cook RL, Lee Allen H, Albaugh TJ, Rubilar R, Campoe O. A New Approach for Modeling Volume Response from Mid-Rotation Fertilization of Pinus taeda L. Plantations. Forests. 2020; 11(6):646. https://doi.org/10.3390/f11060646
Chicago/Turabian StyleScolforo, Henrique F., Cristian Montes, Rachel L. Cook, Howard Lee Allen, Timothy J. Albaugh, Rafael Rubilar, and Otavio Campoe. 2020. "A New Approach for Modeling Volume Response from Mid-Rotation Fertilization of Pinus taeda L. Plantations" Forests 11, no. 6: 646. https://doi.org/10.3390/f11060646