Nitrogen Forms Alter Triterpenoid Accumulation and Related Gene Expression in Cyclocarya paliurus (Batalin) Iljinsk. Seedlings


Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Experimental Design
2.2. Plant Material Sampling and Biomass Measurements
2.3. Measurement of Triterpenoid Contents
2.4. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis
2.5. Statistical Analysis
3. Results
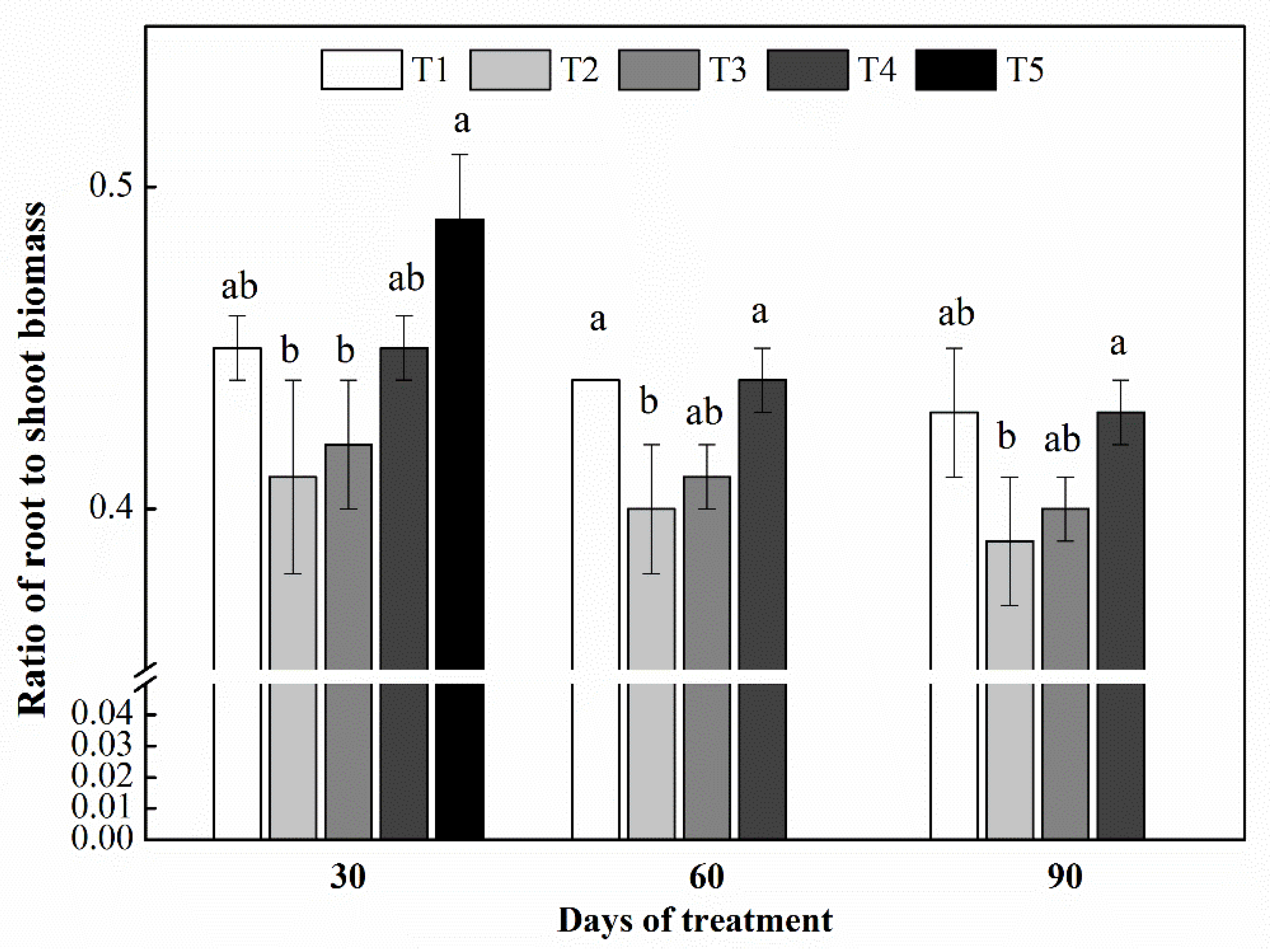
3.1. Seedling Biomass
3.2. Triterpenoid Contents
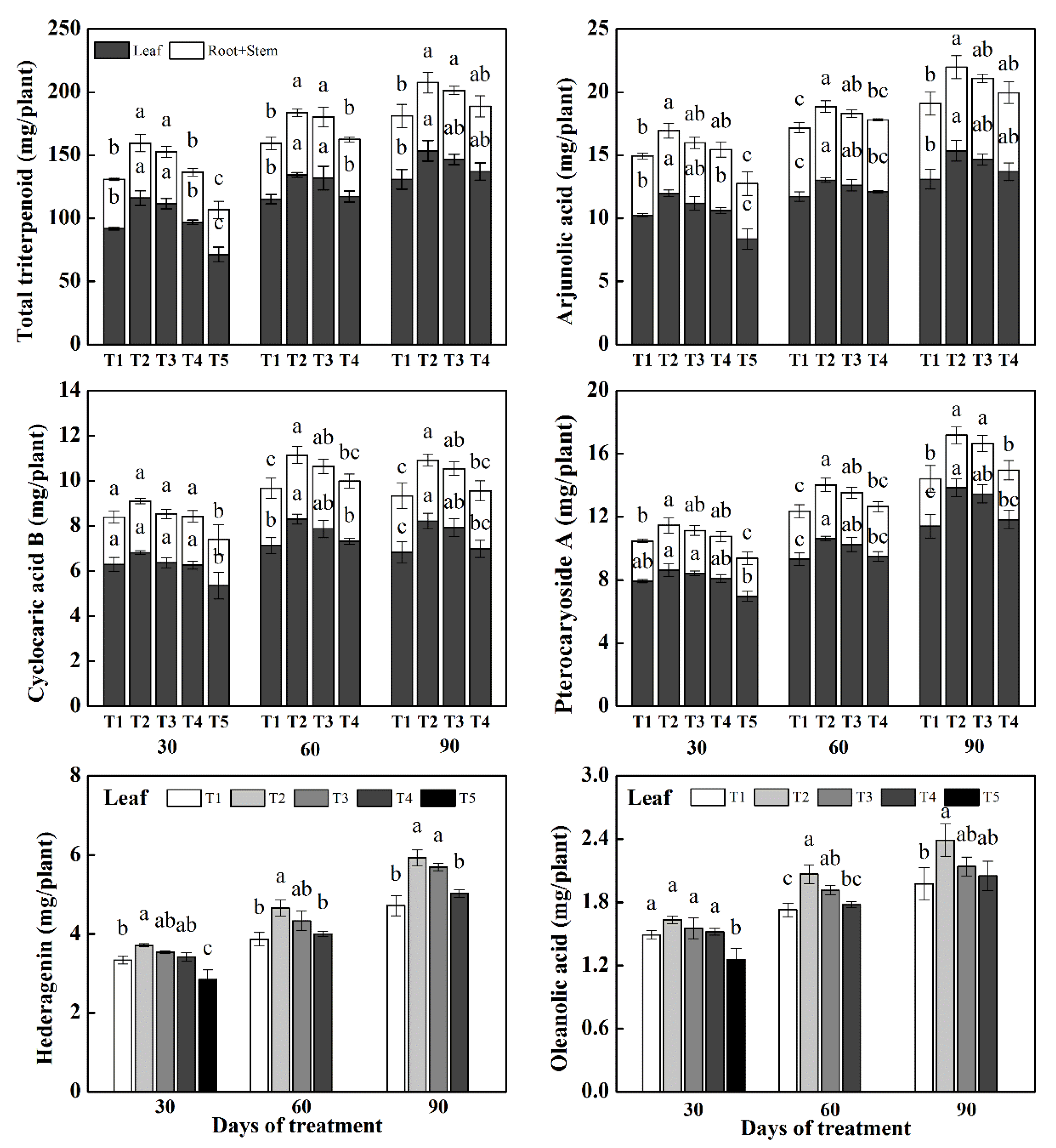
3.3. Triterpenoid Accumulation per Plant
3.4. Gene Expression Related to Triterpenoid Biosynthesis
4. Discussion
4.1. Effects of NO3−/NH4+ Ratio on Growth and Triterpenoid Content
4.2. Effects of NO3−/NH4+ Ratio on Relative Gene Expression
4.3. Effects of NO3−/NH4+ Ratio on Triterpenoid Production per Plant
5. Conclusions
Supplementary Files
Supplementary File 1Author Contributions
Funding
Conflicts of Interest
References
- Chung, I.M.; Miller, D.A. Natural herbicide potential of alfalfa residue on selected weed species. Agron. J. 1995, 87, 920–925. [Google Scholar] [CrossRef]
- Somova, L.O.; Nadar, A.; Rammanan, P.; Shode, F.O. Cardiovascular, antihyperlipidemic and antioxidant effects of oleanolic and ursolic acids in experimental hypertension. Phytomedicine 2003, 10, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, A.; Hamid, K.; Kam, A.; Wong, K.H.; Abdelhak, Z.; Razmovski-Naumovski, V.; Chan, K.; Li, K.M.; Groundwater, P.W.; Li, G.Q. The pentacyclic triterpenoids in herbal medicines and their pharmacological activities in diabetes and diabetic complications. Curr. Med. Chem. 2013, 20, 908–931. [Google Scholar]
- Zhou, M.M.; Chen, P.; Lin, Y.; Fang, S.Z.; Shang, X.L. A comprehensive assessment of bioactive metabolites, antioxidant and antiproliferative activities of Cyclocarya paliurus (Batal.) Iljinskaja leaves. Forests 2019, 10, 625. [Google Scholar] [CrossRef] [Green Version]
- Pulido, P.; Perello, C.; Rodriguez-Concepcion, M. New insights into plant isoprenoid metabolism. Mol. Plant. 2012, 5, 964–967. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Li, C. Biosynthesis of plant triterpenoid saponins in microbial cell factories. J. Agric. Food Chem. 2018, 66, 12155–12165. [Google Scholar] [CrossRef]
- Zhao, C.L.; Cui, X.M.; Chen, Y.P.; Liang, Q. Key enzymes of triterpenoid saponin biosynthesis and the induction of their activities and gene expressions in plants. Nat. Prod. Commun. 2010, 5, 1147–1158. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.Z.; Wang, J.Y.; Wei, Z.Y.; Zhu, Z.X. Methods to break seed dormancy in Cyclocarya paliurus (Batal.) Iljinskaja. Sci. Hortic. 2006, 110, 305–309. [Google Scholar] [CrossRef]
- Xie, J.H.; Xie, M.Y.; Nie, S.P.; Shen, M.Y.; Wang, Y.X.; Li, C. Isolation, chemical composition and antioxidant activities of a water-soluble polysaccharide from Cyclocarya paliurus (Batal.) Iljinskaja. Food Chem. 2010, 119, 1626–1632. [Google Scholar] [CrossRef]
- Fang, S.Z.; Yang, W.X.; Chu, X.L.; Shang, X.L.; She, C.Q.; Fu, X.X. Provenance and temporal variations in selected flavonoids in leaves of Cyclocarya paliurus. Food Chem. 2011, 124, 1382–1386. [Google Scholar] [CrossRef]
- Xie, J.H.; Wang, Z.J.; Shen, M.Y.; Nie, S.P.; Xie, M.Y. Sulfated modification, characterization and antioxidant activities of polysaccharide from Cyclocarya paliurus. Food Hydrocolloids. 2016, 53, 7–15. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, S.Z.; Zhou, M.M.; Shang, X.L.; Yang, W.X.; Fu, X.X. Geographic variation in water-soluble polysaccharide content and antioxidant activities of Cyclocarya paliurus leaves. Ind. Crop. Prod. 2018, 121, 180–186. [Google Scholar] [CrossRef]
- Wu, Z.F.; Meng, F.C.; Cao, L.J.; Jiang, C.H.; Zhao, M.G.; Shang, X.L.; Fang, S.Z.; Ye, W.C.; Zhang, Q.W.; Zhang, J.; et al. Triterpenoids from Cyclocarya paliurus and their inhibitory effect on the secretion of apoliprotein B48 in Caco-2 cells. Phytochemistry 2017, 142, 76–84. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, Y.N.; Fang, S.Z.; Wang, T.L.; Yin, Z.Q.; Shang, X.L.; Yang, W.X.; Fu, X.X. Antidiabetic effect of Cyclocarya paliurus leaves depends on the contents of antihyperglycemic flavonoids and antihyperlipidemic triterpenoids. Molecules 2018, 23, 1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.J.; Zhong, Z.C.; Xie, Z.M. Studies on the sweet principles from the leaves of Cyclocarya paliurus (Batal.) Iljinskaya. Acta Pharm Sin. 1992, 27, 841–844. [Google Scholar]
- Shu, R.G.; Xu, C.R.; Li, L.N. Studies on the sweet principles from the leaves of Cyclocarya paliurus (Batal.) Iljinsk. Acta Pharm Sin. 1995, 30, 757–761. [Google Scholar]
- Guo, S.; Zhou, Y.; Shen, Q.; Zhang, F. Effect of ammonium and nitrate nutrition on some physiological processes in higher plants-growth, photosynthesis, photorespiration, and water relations. Plant Biol. 2007, 9, 21–29. [Google Scholar] [CrossRef]
- Tang, Z.H.; Liu, Y.J.; Guo, X.R.; Zu, Y.G. The combined effects of salinity and nitrogen forms on Catharanthus roseus: The role of internal ammonium and free amino acids during salt stress. J. Plant Nutr. Soil Sci. 2011, 174, 135–144. [Google Scholar]
- Bowler, J.M.; Press, M.C. Effects of elevated CO2, nitrogen form and concentration on growth and photosynthesis of a fast- and slow-growing grass. New Phytol. 1996, 132, 391–401. [Google Scholar] [CrossRef]
- Makino, A.; Mae, T. Photosynthesis and plant growth at elevated levels of CO2. Plant Cell Physiol. 1999, 40, 999–1006. [Google Scholar] [CrossRef]
- Moore, B.D.; Cheng, S.H.; Sims, D.; Seemann, J.R. The biochemical and molecular basis for photosynthetic acclimation to elevated atmospheric CO2. Plant Cell Environ. 1999, 22, 567–582. [Google Scholar] [CrossRef]
- Chen, W.; Luo, J.K.; Shen, Q.R. Effect of NH4+-N/NO3–-N ratios on growth and some physiological parameters of Chinese cabbage cultivars. Pedosphere 2005, 15, 310–318. [Google Scholar]
- Prasad, A.; Mathur, A.; Singh, M.; Gupta, M.M.; Uniyal, G.C.; Lal, R.K.; Mathur, A.K. Growth and asiaticoside production in multiple shoot cultures of a medicinal herb, Centella asiatica (L.) Urban, under the influence of nutrient manipulations. J. Nat. Med. 2012, 66, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Tan, R.X. Artemisinin production in Artemisia annua hairy root cultures with improved growth by altering the nitrogen source in the medium. Biotechnol. Lett. 2002, 24, 1153–1156. [Google Scholar] [CrossRef]
- Liu, C.Z.; Guo, C.; Wang, Y.C.; Ouyang, F. Factors influencing artemisinin production from shoot cultures of Artemisia annua L. World J. Microbiol. Biotechnol. 2003, 19, 535–538. [Google Scholar] [CrossRef]
- Lee, E.J.; Paek, K.Y. Effect of nitrogen source on biomass and bioactive compound production in submerged cultures of Eleutherococcus koreanum Nakai adventitious roots. Biotechnol. Prog. 2012, 28, 508–514. [Google Scholar] [CrossRef]
- Zhu, Z.B.; Yu, M.M.; Chen, Y.H.; Guo, Q.S.; Zhang, L.X.; Shi, H.Z.; Liu, L. Effects of ammonium to nitrate ratio on growth, nitrogen metabolism, photosynthetic efficiency and bioactive phytochemical production of Prunella vulgaris. Pharm. Biol. 2014, 52, 1518–1525. [Google Scholar] [CrossRef] [Green Version]
- Figura, T.; Weiser, M.; Ponert, J. Orchid seed sensitivity to nitrate reflects habitat preferences and soil nitrate content. Plant Biol. 2020, 22, 21–29. [Google Scholar] [CrossRef]
- Liu, M.Y.; Burgos, A.; Zhang, Q.F.; Tang, D.D.; Shi, Y.Z.; Ma, L.F.; Yi, X.Y.; Ruan, J.Y. Analyses of transcriptome profiles and selected metabolites unravel the metabolic response to NH4+ and NO3− as signaling molecules in tea plant (Camellia sinensis L.). Sci. Hortic. 2017, 218, 293–303. [Google Scholar] [CrossRef]
- Karray-Bouraoui, N.; Harbaoui, F.; Rabhi, M.; Jallali, I.; Ksouri, R.; Attia, H.; Msilini, N.; Lachaâl, M. Different antioxidant responses to salt stress in two different provenances of Carthamus tinctorius L. Acta Physiol. Plant. 2010, 33, 1435–1444. [Google Scholar] [CrossRef]
- Deng, B.; Shang, X.L.; Fang, S.Z.; Li, Q.Q.; Fu, X.X.; Su, J. Integrated effects of light intensity and fertilization on growth and flavonoid accumulation in Cyclocarya paliurus. J. Agric. Food Chem. 2012, 60, 6286–6292. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.N.; Fang, S.Z.; Yin, Z.Q.; Fu, X.X.; Shang, X.L.; Yang, W.X.; Yang, H.M. Chemical fingerprint and multicomponent quantitative analysis for the quality evaluation of Cyclocarya paliurus leaves by HPLC–Q–TOF–MS. Molecules 2017, 22, 1927. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.P.; He, C.H. Simultaneous quantification of three major bioactive triterpene acids in the leaves of Diospyros kaki by high-performance liquid chromatography method. J. Pharm. Biomed. Anal. 2006, 41, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Mao, X.; Huang, P.; Fang, S.Z. Morphological characterization of flower buds development and related gene expression profiling at bud break stage in heterodichogamous Cyclocarya paliurus (Batal.) lljinskaja. Genes 2019, 10, 818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulconnier, Y.; Boby, C.; Pires, J.; Labonne, C.; Lerous, C. Effects of Azgp1(-/-) on mammary gland, adipose tissue and liver gene expression and milk lipid composition in lactating mice. Gene 2019, 692, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef] [Green Version]
- Hessini, K.; Issaoui, K.; Ferchichi, S.; Abdelly, C.; Siddique, K.H.M.; Cruz, C. Interactive effects of salinity and nitrogen forms on plant growth, photosynthesis and osmotic adjustment in maize. Plant Physiol. Biochem. 2019, 139, 171–178. [Google Scholar] [CrossRef]
- Zhang, C.X.; Meng, S.; Li, Y.M.; Su, L.; Zhao, Z. Nitrogen uptake and allocation in Populus simonii in different seasons supplied with isotopically labeled ammonium or nitrate. Trees Struct. Funct. 2016, 30, 2011–2018. [Google Scholar] [CrossRef]
- Boschiero, B.N.; Mariano, E.; Azevedo, R.A.; Trivelin, P.C.O. Influence of nitrate-ammonium ratio on the growth, nutrition, and metabolism of sugarcane. Plant Physiol. Biochem. 2019, 139, 246–255. [Google Scholar] [CrossRef]
- Ruan, J.; Gerendás, J.; Härdter, R.; Sattelmacher, B. Effect of nitrogen form and root-zone pH on growth and nitrogen uptake of tea (Camellia sinensis) plants. Ann. Bot. 2007, 99, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Glibert, P.M.; Wilkerson, F.P.; Dugdale, R.C.; Raven, J.A.; Dupont, C.L.; Leavitt, P.R.; Parker, A.E.; Burkholder, J.M.; Kana, T.M. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr. 2015, 61, 165–197. [Google Scholar] [CrossRef]
- Heuer, B. Growth, photosynthesis and protein content in cucumber plants as affected by supplied nitrogen form. J. Plant Nutr. 1991, 14, 363–373. [Google Scholar] [CrossRef]
- Leidi, E.O.; Silberbush, M.; Soares, M.I.M.; Lips, S.H. Salinity and nitrogen nutrition studies on peanut and cotton plants. J. Plant Nutr. 1992, 15, 591–604. [Google Scholar] [CrossRef]
- Walch-Liu, P.; Neumann, G.; Bangerth, F.; Engels, C. Rapid effects of nitrogen form on leaf morphogenesis in tobacco. J. Exp. Bot. 2000, 51, 227–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.E.; Lu, L.L.; Jin, Y.; Wei, J.H.; Christie, P. Effects of nitrogen on root development and contents of bioactive compounds in Salvia miltiorrhiza Bunge. Crop Sci. 2013, 53, 2028–2039. [Google Scholar] [CrossRef]
- Taulavuori, K.; Hyöky, V.; Oksanen, J.; Taulavuori, E.; Julkunen-Tiitto, R. Species-specific differences in synthesis of flavonoids and phenolic acids under increasing periods of enhanced blue light. Environ. Exp. Bot. 2016, 121, 145–150. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, S.Z.; Yang, W.X.; Shang, X.L.; Fu, X.X. Light quality affects flavonoid production and related gene expression in Cyclocarya paliurus. J. Photochem. Photobiol. B Biol. 2018, 179, 66–73. [Google Scholar] [CrossRef]
- Deng, B.; Li, Y.Y.; Lei, G.; Liu, G.H. Effects of nitrogen availability on mineral nutrient balance and flavonoid accumulation in Cyclocarya paliurus. Plant Physiol. Biochem. 2019, 135, 111–118. [Google Scholar] [CrossRef]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [Green Version]
- Hakulinen, J.; Julkunen-Tiitto, R.; Tahvanainen, J. Does nitrogen fertilization have an impact on the trade-off between willow growth and defensive secondary metabolism? Trees Struct. Funct. 1995, 9, 235–240. [Google Scholar] [CrossRef]
- Stamp, N. Out of the quagmire of plant defense hypotheses. Q. Rev. Biol. 2003, 78, 23–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamp, N. Can the growth–differentiation balance hypothesis be tested rigorously? Oikos 2004, 107, 439–448. [Google Scholar] [CrossRef]
- Strissel, T.; Halbwirth, H.; Hoyer, U.; Zistler, C.; Stich, K.; Treutter, D. Growth-promoting nitrogen nutrition affects flavonoid biosynthesis in young apple (Malus domestica Borkh.) Leaves. Plant Biol. 2005, 7, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Leser, C.; Treutter, D. Effects of nitrogen supply on growth, contents of phenolic compounds and pathogen(scab) resistance of apple trees. Physiol. Plant. 2005, 123, 49–56. [Google Scholar] [CrossRef]
- Huang, H.; Yao, Q.; Xia, E.; Gao, L. Metabolomics and transcriptomics analyses reveal nitrogen influences on the accumulation of flavonoids and amino acids in young shoots of tea plant (Camellia sinensis L.) associated with tea flavor. J. Agric. Food Chem. 2018, 66, 9828–9838. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Kong, D.X.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence of Primer (5′-3′) |
|---|---|
| HMGR-F | TTTAGCGATGGACATGAGCA |
| HMGR-R | GGAGTGGCAGAGCGTCAGAGGC |
| DXR-F | GCTGGTTCAATGTAACTCTTC |
| DXR-R | CTCTATGACTCCTTGCTCCC |
| GPS-F | GAGCGAAGTTATTCCTGGTG |
| GPS-R | GTGTAAATGGGAGATGAACG |
| SQS-F | GAACAGGCTGGATGCGATAC |
| SQS-R | TCAATTATTTGGTCGTTTGG |
| β-AS-F | TGGTTCATGGTTTGCACTTGGAG |
| β-AS-R | CTCTCTCAGCCTGTCCAGCATGA |
| UDP-F | TCTACTATCACCTCGACCTCCT |
| UDP-R | TTTTTACATCCTGAAATGCCTT |
| 18s-F | AGTATGGTCGCAAGGCTGAAA |
| 18s-R | CAGACAAATCGCTCCACCAA |
| Gene Expression | Total Triterpenoid | Arjunolic Acid | Cyclocaric Acid B | Pterocaryoside A | Hederagenin | Oleanolic Acid |
|---|---|---|---|---|---|---|
| HMGR | 0.50 ** | 0.69 ** | 0.91 ** | 0.57 ** | 0.53 ** | 0.68 ** |
| DXR | 0.61 ** | 0.76 ** | 0.78 ** | 0.79 ** | 0.77 ** | 0.78 ** |
| GPS | 0.43 ** | 0.54 ** | 0.81 ** | 0.44 ** | 0.41 ** | 0.58 ** |
| SQS | 0.73 ** | 0.80 ** | 0.87 ** | 0.73 ** | 0.71 ** | 0.81 ** |
| β-AS | 0.71 ** | 0.74 ** | 0.64 ** | 0.82 ** | 0.81 ** | 0.77 ** |
| UDP | 0.50 ** | 0.69 ** | 0.91 ** | 0.57 ** | 0.53 ** | 0.68 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, J.; Yue, X.; Shang, X.; Fang, S. Nitrogen Forms Alter Triterpenoid Accumulation and Related Gene Expression in Cyclocarya paliurus (Batalin) Iljinsk. Seedlings. Forests 2020, 11, 631. https://doi.org/10.3390/f11060631
Qin J, Yue X, Shang X, Fang S. Nitrogen Forms Alter Triterpenoid Accumulation and Related Gene Expression in Cyclocarya paliurus (Batalin) Iljinsk. Seedlings. Forests. 2020; 11(6):631. https://doi.org/10.3390/f11060631
Chicago/Turabian StyleQin, Jian, Xiliang Yue, Xulan Shang, and Shengzuo Fang. 2020. "Nitrogen Forms Alter Triterpenoid Accumulation and Related Gene Expression in Cyclocarya paliurus (Batalin) Iljinsk. Seedlings" Forests 11, no. 6: 631. https://doi.org/10.3390/f11060631