Dynamics in Stoichiometric Traits and Carbon, Nitrogen, and Phosphorus Pools across Three Different-Aged Picea asperata Mast. Plantations on the Eastern Tibet Plateau

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Field Sampling and Measurements
2.3. Statistical Analysis
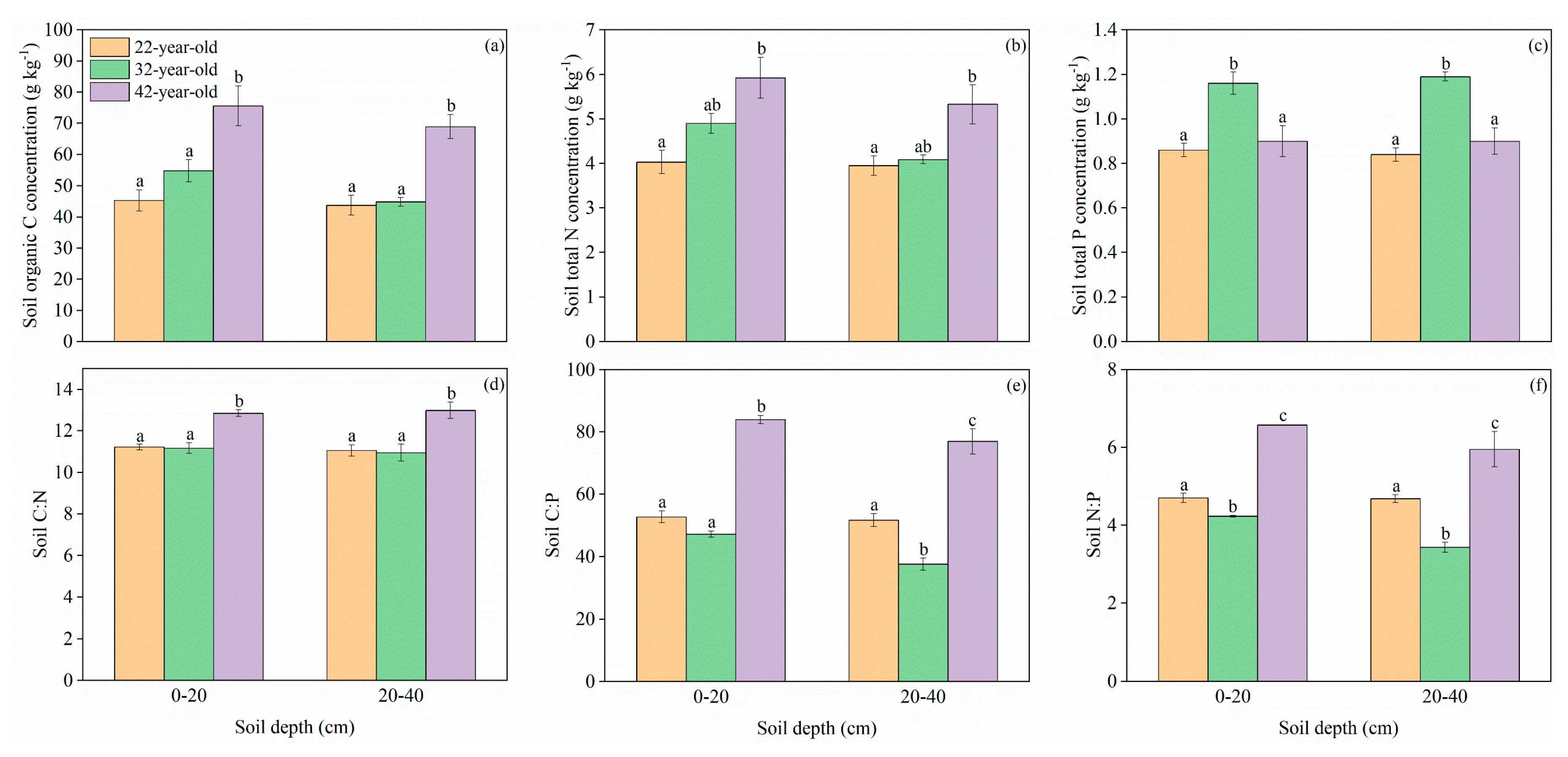
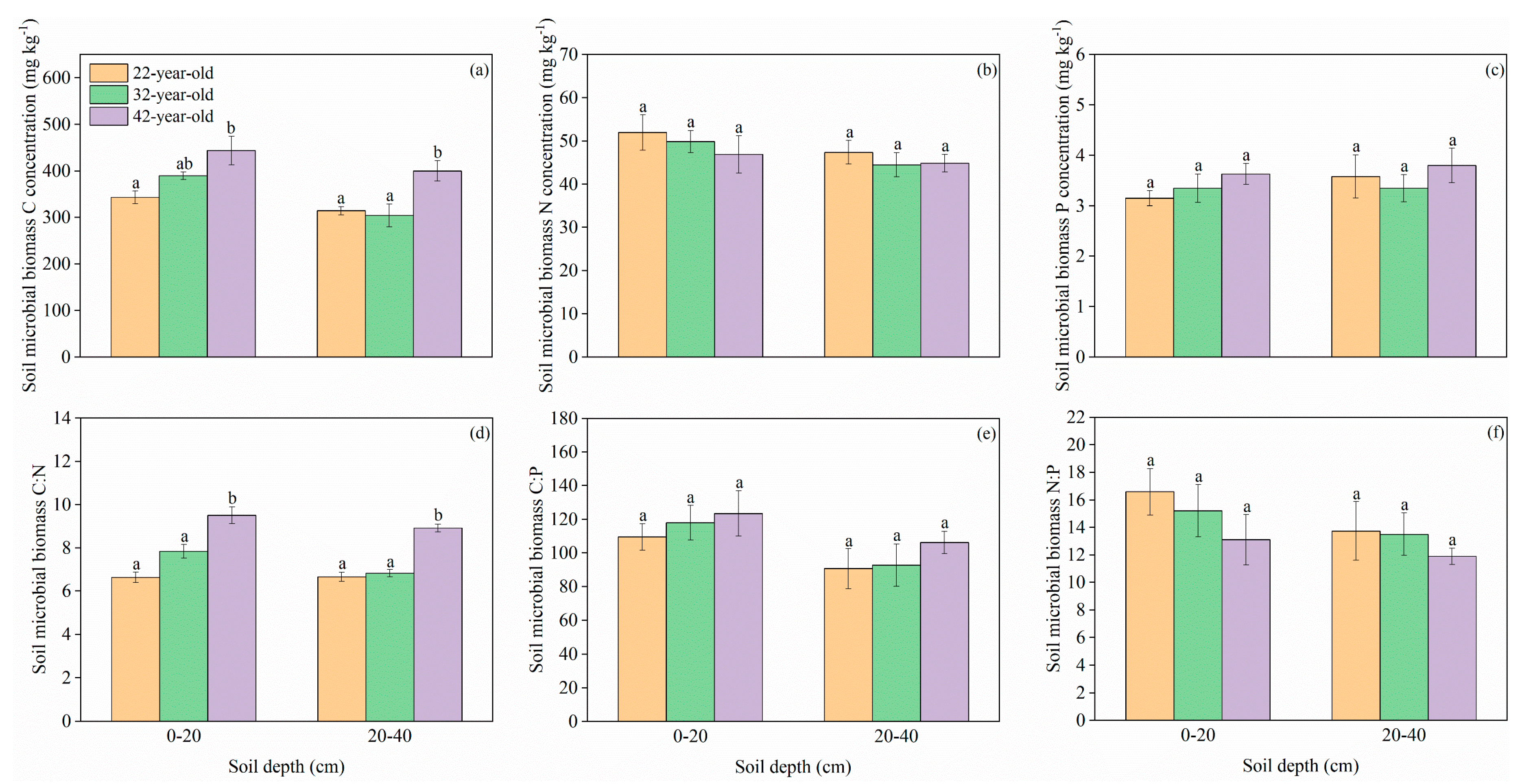
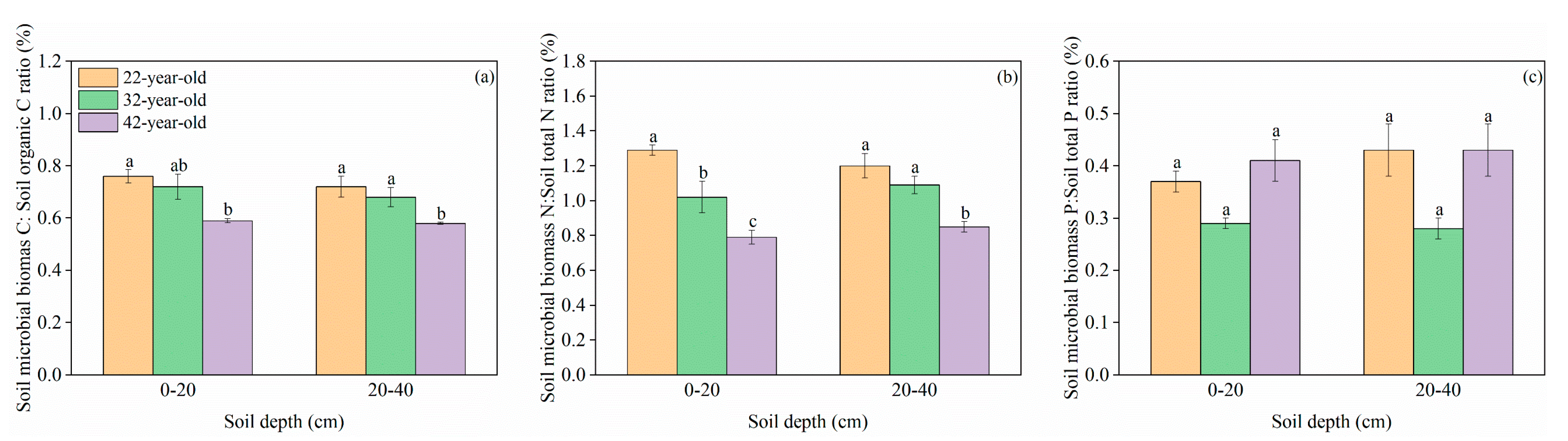
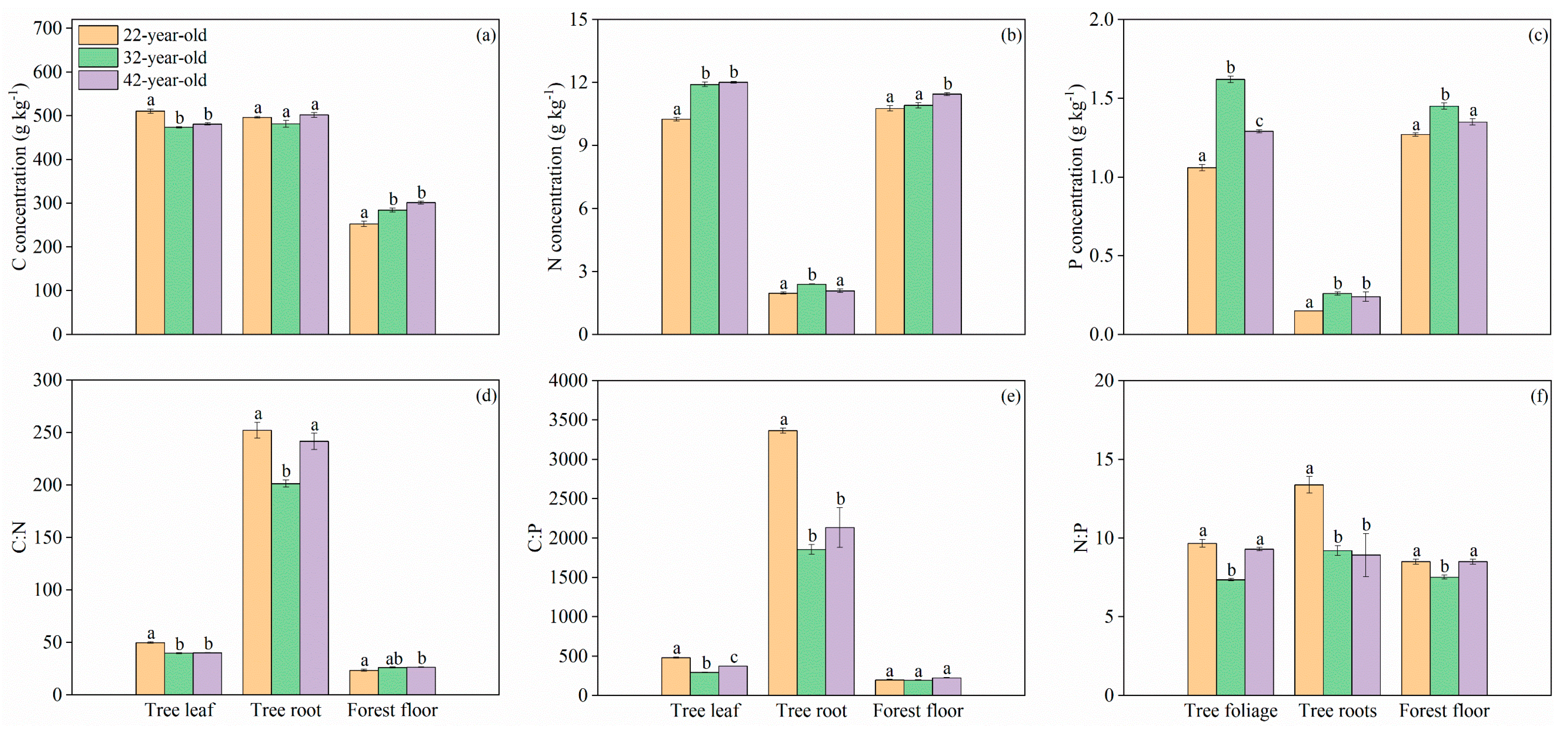
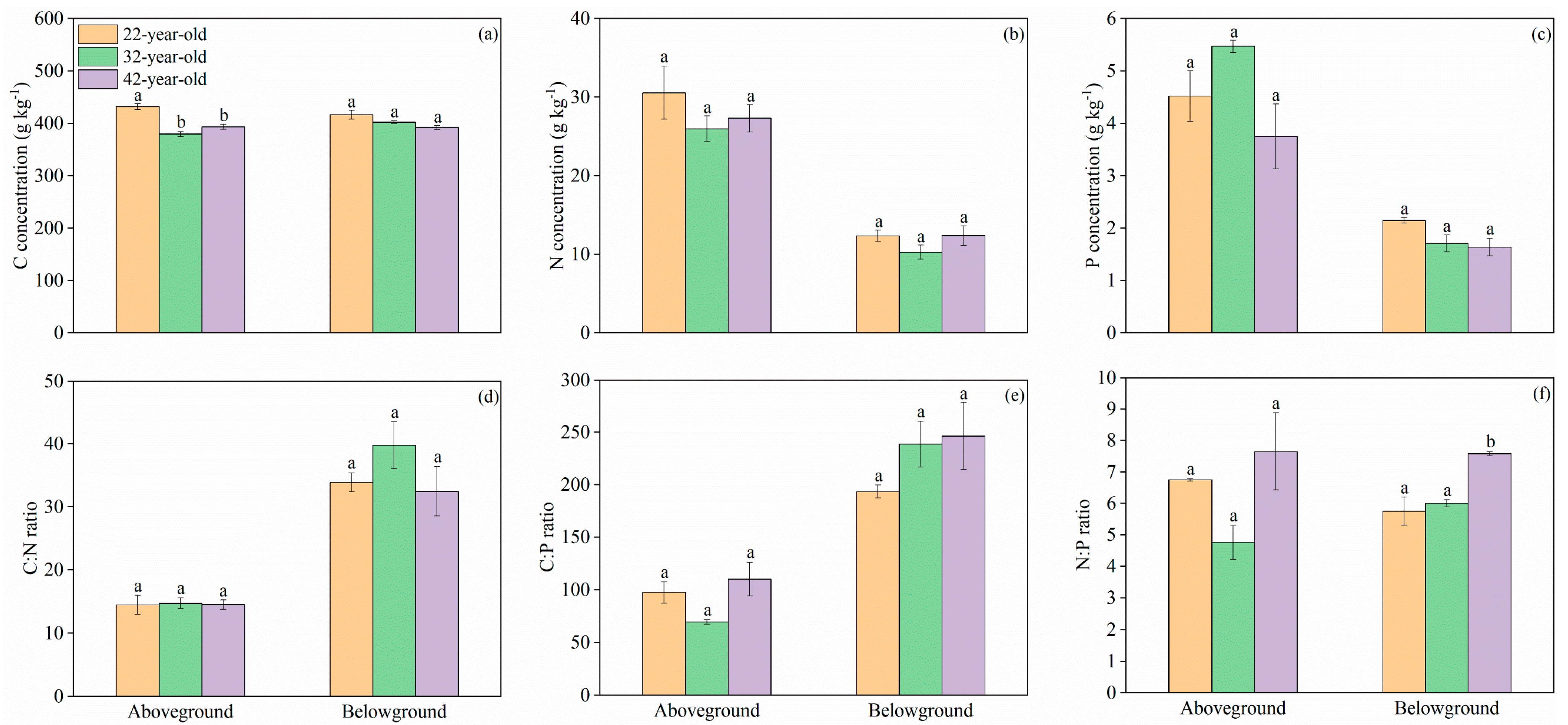
3. Results
3.1. C, N, and P Concentrations and Stoichiometric Traits in Mineral Soil and Soil Microbial Biomass
3.2. C, N, and P Concentrations and Stoichiometric Traits in Plants and Forest Floor
3.3. Relationship between the Stoichiometric Traits of Mineral Soil and Other Ecosystem Components
3.4. Ecosystem C, N, and P Pools
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sun, L.; Zhang, B.; Wang, B.; Zhang, G.; Zhang, W.; Zhang, B.; Chang, S.; Chen, T.; Liu, G. Leaf elemental stoichiometry of Tamarix Lour. species in relation to geographic, climatic, soil, and genetic components in China. Ecol. Eng. 2017, 106, 448–457. [Google Scholar] [CrossRef]
- Elser, J.J. Ecological stoichiometry: From sea to lake to land. Trends Ecol. Evol. 2000, 15, 393–394. [Google Scholar] [CrossRef]
- Elser, J.J.; Dobberfuhl, D.R.; Mackay, N.A.; Schampel, J.H. Organism size, life history and N:P stoichiometry. Bioscience 1996, 46, 674–684. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Wang, X.; Tian, Y.; Wen, Z.; Zha, T. Pattern of carbon allocation across three different stages of stand development of a Chinese pine (Pinus tabulaeformis) forest. Ecol. Res. 2012, 27, 883–892. [Google Scholar] [CrossRef]
- Larsen, M.L.; Wihlelm, S.W.; Lennon, J.T. Nutrient stoichiometry shapes microbial coevolution. Ecol. Lett. 2019, 22, 1009–1018. [Google Scholar] [CrossRef]
- Chen, L.; Deng, Q.; Yuan, Z.; Mu, X.; Kallenbach, R.L. Age-related C:N:P stoichiometry in two plantation forests in the Loess Plateau. Ecol. Eng. 2018, 120, 14–22. [Google Scholar] [CrossRef]
- Peng, Y.; Li, F.; Zhou, G.; Fang, K.; Zhang, D.; Li, C.; Yang, G.; Wang, G.; Wang, J.; Yang, Y. Linkages of plant stoichiometry to ecosystem production and carbon fluxes with increasing nitrogen inputs in an alpine steppe. Glob. Chang. Biol. 2016, 23, 5249–5259. [Google Scholar] [CrossRef]
- Bielski, R.L. Phosphate pools, phosphate transport, and phosphate availability. Ann. Rev. Plant. Physiol. 1973, 24, 225–252. [Google Scholar] [CrossRef]
- Rong, Q.; Liu, J.; Cai, Y.; Lu, Z.; Zhao, Z.; Yue, W.; Xia, J. Leaf carbon, nitrogen and phosphorus stoichiometry of Tamarix chinensis Lour. in the Laizhou Bay coastal wetland, China. Ecol. Eng. 2015, 76, 57–65. [Google Scholar] [CrossRef]
- Li, P.; Yang, Y.; Han, W.; Fang, J. Global patterns of soil microbial nitrogen and phosphorus stoichiometry in forest ecosystems. Glob. Ecol. Biogeogr. 2014, 23, 979–987. [Google Scholar] [CrossRef]
- Leal, M.C.; Seehausen, O.; Matthews, B. The ecology and evolution of stoichiometric phenotypes. Trends Ecol. Evol. 2017, 32, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Fan, J.; Li, Y. Foliar carbon, nitrogen, and phosphorus stoichiometry in a grassland ecosystem along the Chinese Grassland Transect. Acta Ecol. Sin. 2017, 37, 133–139. [Google Scholar] [CrossRef]
- Cao, J.; Shang, H.; Chen, Z.; Tian, Y.; Yu, H. Effects of elevated ozone on stoichiometry and nutrient pools of Phoebe bourneri (Hemsl.) Yang and Phoebe Zhennan S. Lee et F. N. Wei seedlings in subtropical China. Forests 2016, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Heuck, C.; Weig, A.; Spohn, M. Soil microbial biomass C:N:P stoichiometry and microbial use of organic phosphorus. Soil Biol. Biochem. 2015, 85, 119–129. [Google Scholar] [CrossRef]
- Fan, H.; Wu, J.; Liu, W.; Yuan, Y.; Hu, L.; Cai, Q. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant. Soil 2015, 392, 127–138. [Google Scholar] [CrossRef]
- Visser, S.; Keesstra, S.; Maas, G.; de Cleen, M.; Molenaar, C. Soil as a basis to create enabling conditions for transitions towards sustainable land management as a key to achieve the SDGs by 2030. Sustainability 2019, 11, 6792. [Google Scholar] [CrossRef] [Green Version]
- Minden, V.; Kleyer, M. Internal and external regulation of plant organ stoichiometry. Plant. Biol. 2014, 16, 897–907. [Google Scholar] [CrossRef]
- Fierer, N.; Strickland, D.L.; Bradford, M.A.; Cleveland, C.C. Global patterns in belowground communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Ren, C.J.; Han, X.H.; Yang, G.H.; Doughty, R. Changes of soil microbial and enzyme activities are linked to soil C, N and P stoichiometry in afforested ecosystems. For. Ecol. Manag. 2018, 427, 289–295. [Google Scholar] [CrossRef]
- Zhou, Z.-H.; Wang, C.-K. Changes of the relationships between soil and microbes in carbon, nitrogen and phosphorus stoichiometry during ecosystem succession. Chin. J. Plant. Ecol. 2016, 40, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India lead in greening of the world through land-use management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.-S.; Piao, S.; Zeng, Z.; Ciais, P.; Zhou, L.; Li, L.Z.X.; Myneni, R.B.; Yin, Y.; Zeng, H. Afforestation in China cools local land surface temperature. Proc. Natl. Acad. Sci. USA 2014, 111, 2915–2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Wang, X.; Li, N. Combating climate change: What will China’s Forestry do? Ambio 2010, 39, 340–343. [Google Scholar] [CrossRef] [Green Version]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, L.; Gallichand, J. Runoff responses to afforestation in watershed of the Loess Plateau, China. Hydrol. Process. 2003, 17, 2599–2609. [Google Scholar] [CrossRef]
- Li, C.; Zhou, X. Status and future trends in plantation silviculture in China. Ambio 2000, 29, 354–355. [Google Scholar] [CrossRef]
- Miao, N.; Liu, S.; Shi, Z.; Yu, H.; Liu, X. Spatial patterns of dominant tree species in sub-alpine Betula-abies forest in West Sichuan of China. Chin. J. Appl. Ecol. 2009, 20, 1263–1270. [Google Scholar]
- Liu, Y.; Chen, J.; Liu, Q.; Wu, Y. Nitrification and denitrification in subalpine coniferous forests of different restoration stages in western Sichuan, China. Front. For. China 2007, 2, 260–265. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhao, C.; Liu, S. The influence factors of sub-alpine forest restoration in Miyaluo, West Sichuan. Sci. Silvae Sin. 2005, 41, 189–193. [Google Scholar]
- Pang, X.-Y.; Liu, Q.; Liu, S.-Q.; Wu, Y.; Lin, B.; He, H.; Zhang, Z.-J. Changes of soil fertility quality properties under subalpine spruce plantation in Western Sichuan. Acta Ecol. Sin. 2004, 24, 261–267. [Google Scholar]
- Pang, X.; Huang, J.; Zhao, Q.; Feng, D.; Bao, W.; Tian, G. Ecosystem carbon stock across a chronosequence of spruce plantations established on cutovers of a high-elevation region. J. Soil Sediments 2016, 17, 2239–2249. [Google Scholar] [CrossRef]
- Xu, Z.; Yin, H.; Xiong, P.; Wan, C.; Liu, Q. Short-term responses of Picea asperata seedlings of different ages grown in two contrasting forest ecosystems to experimental warming. Environ. Exp. Bot. 2012, 77, 1–11. [Google Scholar] [CrossRef]
- Liu, G.S. Soil Physical and Chemical Analysis and Description of Soil Profiles, 1st ed.; Standards Press of China: Beijing, China, 1996; pp. 123–167. [Google Scholar]
- Bao, S.D. Agricultural Soil Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 1999; pp. 74–270. [Google Scholar]
- Cao, J.; Yan, W.; Xiang, W.; Chen, X.; Lei, P. Stoichiometry characterization of soil C, N, and P of Chinese fir plantations of three different ages in Huitong, Hunan province, China. Sci. Silvae Sin. 2015, 51, 1–8. [Google Scholar]
- Cui, N.; Liu, X.; Zhang, D.; Zhang, J.; Liu, Y.; Deng, C.; Ji, T.; Chen, Y. The distribution pattern of carbon, nitrogen and phosphorus and the stoichiometry characteristics of Pinus massoniana plantation in different ages. Ecol. Environ. Sci. 2014, 23, 188–195. [Google Scholar]
- Li, Q.; Mo, Q.; Wang, F.; Li, Y.; Xu, X.; Zou, B.; Li, X.; Chen, Y.; Li, Z. Nutrient utilization by Casuarina equisetifolia plantation of different ages in the tropical coastal area of South China. Chin. J. Appl. Environ. Biol. 2015, 21, 139–146. [Google Scholar]
- Mao, R.; Zeng, D.-H.; Hu, Y.-L.; Li, L.-J.; Yang, D. Soil organic carbon and nitrogen stocks in an age-sequence pf poplar stands planted on marginal agricultural land in Northeast China. Plant. Soil 2010, 332, 277–287. [Google Scholar] [CrossRef]
- Noh, N.-J.; Son, Y.; Lee, S.-K.; Seo, K.-W.; Heo, S.-J.; Yi, M.-J.; Park, P.-S.; Kim, R.-H.; Son, Y.-M.; Lee, K.-H. Carbon and nitrogen storage in an age-sequence of Pinus densiflora stands in Korea. Sci. China Life Sci. 2010, 53, 822–830. [Google Scholar] [CrossRef]
- Wu, H.; Xiang, W.; Chen, L.; Ouyang, S.; Xiao, W.; Li, S.; Forrester, D.I.; Lei, P.; Zeng, Y.; Deng, X.; et al. Soil phosphorus bioavailability and recycling increased with stand age in Chinese fir plantations. Ecosystems 2020, 23, 973–988. [Google Scholar] [CrossRef]
- Wu, H.; Xiang, W.; Ouyang, S.; Xiao, W.; Li, S.; Chen, L.; Lei, P.; Deng, X.; Zeng, Y.; Zeng, L.; et al. Tree growth rate and soil nutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence. For. Ecol. Manag. 2020, 476, 117896. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, C.K. Reviews and syntheses: Soil resources and climate jointly drive variations in microbial biomass carbon and nitrogen in China’s forest ecosystems. Biogeosciences 2015, 6751–6760. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Schimel, J.P.; Thornton, P.E.; Song, X.; Yuan, F.; Goswami, S. Substrate and environmental controls on microbial assimilation of soil organic carbon: A framework for earth system models. Ecol. Lett. 2014, 17, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.-H.; Wang, C.-K. Responses and regulation mechanisms of microbial decomposers to substrate carbon, nitrogen, and phosphorus stoichiometry. Chin. J. Plant. Ecol. 2016, 40, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Fanin, N.; Fromin, N.; Buatois, B.; Hättenschwiller, S. An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system. Ecol. Lett. 2013, 16, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Cui, Q.; Zhao, Q.; Ai, G.; Li, L.; Zeng, D. Soil microbial biomass and activity in relation to stand age of poplar shelterbelts. Chin. J. Appl. Ecol. 2009, 20, 2079–2084. [Google Scholar]
- Ren, H.; Chen, H.; Li, Z.; Han, W. Biomass accumulation and carbon storage of four different aged Sonneratia apetala plantations in Southern China. Plant. Soil 2010, 327, 279–291. [Google Scholar] [CrossRef]
- Bert, D.; Danjon, F. Carbon concentration variations in the roots, stem and crown of mature Pinus Pinaster (Ait.). For. Ecol. Manag. 2006, 222, 279–295. [Google Scholar] [CrossRef]
- Li, M.; Yu, L.; Du, M.; Huang, Z.; Shi, J. C, N, and P stoichiometry and their interaction with plants, litter, and soil in a Cunninghamia lanceolata plantation with different ages. Acta Ecol. Sin. 2018, 38, 7772–7781. [Google Scholar]
- Nehe, A.S.; Misra, S.; Misra, S.; Murchie, E.H.; Chinnathambi, K.; Foulkes, M.J. Genetic variation in N-use efficiency and associated traits in Indian wheat cultivars. Field Crops Res. 2018, 225, 152–162. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant. Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.J. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Matzek, V.; Vitousek, P.M. N:P stoichiometry and protein: RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis. Ecol. Lett. 2009, 12, 765–771. [Google Scholar] [CrossRef]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Shi, X.-M.; Qi, J.-H.; Song, L.; Liu, W.-Y.; Huang, J.-B.; Li, S.; Lu, H.-Z.; Chen, X. C, N and P stoichiometry of two dominant seedlings and their responses to nitrogen additions in the montane moist evergreen broad-leaved forest in Ailao Mountains, Yunnan. Chin. J. Plant. Ecol. 2015, 39, 962–970. [Google Scholar]
- Schreeg, L.A.; Santiago, L.S.; Wright, S.J.; Turner, B.L. Stem, root, and old leaf N:P ratios are more responsive indicators of soil nutrient availability than new foliage. Ecology 2014, 95, 2062–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrish, V.; Cernusak, L.; Winter, K.; Turner, B.L. Nitrogen to phosphorus ratio of plant biomass versus soil solution in a tropical pioneer tree, Ficus insipida. J. Exp. Bot. 2010, 61, 3735–3748. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, W.; Huang, J.; Hu, T.; Tang, D.; Chen, Q. Characteristics of plant ecological stoichiometry homeostasis. Guihaia 2019, 39, 701–712. [Google Scholar] [CrossRef]
- Alcañiz, M.; Úbeda, X.; Cerdà, A. A 13-Year Approach to Understand the Effect of Prescribed Fires and Livestock Grazing on Soil Chemical Properties in Tivissa, NE Iberian Peninsula. Forests 2020, 11, 1013. [Google Scholar] [CrossRef]
- Novara, A.; Pulido, M.; Rodrigo-Comino, J.; Di Prima, S.; Smith, P.; Gristina, L.; Giménez-Morera, A.; Terol, E.; Salesa, D.; Keesstra, S. Long-term organic farming on a citrus plantation results in soil organic carbon recovery. Geogr. Res. Lett. 2019, 45, 271–286. [Google Scholar] [CrossRef] [Green Version]
- Keesstra, S.; Mol, G.; de Leeuw, J.; Okx, J.; Molenaar, C.; de Cleen, M.; Visser, S. Soil-related sustainable development goals: Four concepts to make land degradation neutrality and restoration work. Land 2018, 7, 133. [Google Scholar] [CrossRef] [Green Version]
- Keesstra, S.D.; Bouma, J.; Wallinga, J.; Tittonell, P.; Smith, P.; Cerdà, A.; Montanarella, L.; Quinton, J.N.; Pachepsky, Y.; van der Putten, W.H.; et al. The significance of soils and soil science towards realization of the United Nations sustainable development goals. Soil 2016, 2, 111–128. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Age | Elevation (m a.s.l.) | Slope Degree (°) | Slope Aspect | Density (Trees hm−2) | Mean DBH (cm) |
|---|---|---|---|---|---|
| 22 | 3308 | 15–20 | East | 1733 | 11.93 |
| 32 | 3330 | 18–25 | East | 1558 | 18.34 |
| 42 | 3360 | 11–24 | East | 792 | 23.32 |
| Tree Leaf | Tree Root | Aboveground Herb | Belowground Herb | Forest Floor | Soil Microbes (0–20 cm) | Soil Microbes (20–40 cm) | |
|---|---|---|---|---|---|---|---|
| Mineral Soil (0–20 cm) | −0.405 | 0.324 | −0.189 | −0.353 | −0.340 | 0.734 * | 0.919 ** |
| Mineral Soil (20–40 cm) | −0.383 | 0.425 | 0.362 | −0.274 | 0.514 | 0.743 * | 0.840 ** |
| Tree Leaf | Tree Root | Aboveground Herb | Belowground Herb | Forest Floor | Soil Microbes (0–20 cm) | Soil Microbes (20–40 cm) | |
|---|---|---|---|---|---|---|---|
| Mineral Soil (0–20 cm) | 0.046 | −0.218 | 0.546 | 0.298 | 0.787 * | 0.300 | 0.370 |
| Mineral Soil (20–40 cm) | 0.267 | 0.069 | 0.749 * | 0.154 | 0.852 ** | 0.039 | 0.385 |
| Tree Leaf | Tree Root | Aboveground Herb | Belowground Herb | Forest Floor | Soil Microbes (0–20 cm) | Soil Microbes (20–40 cm) | |
|---|---|---|---|---|---|---|---|
| Mineral Soil (0–20 cm) | 0.504 | −0.307 | 0.652 | 0.834 ** | 0.596 | −0.380 | −0.388 |
| Mineral Soil (20–40 cm) | 0.717 * | 0.083 | 0.803 ** | 0.711 * | 0.799 * | −0.421 | −0.278 |
| 22-Year-Old Stand | 32-Year-Old Stand | 42-Year-Old Stand | ||||
|---|---|---|---|---|---|---|
| C Stock (kg hm−2) | Ratio (%) | C Stock (kg hm−2) | Ratio (%) | C Stock (kg hm−2) | Ratio (%) | |
| Aboveground tree | 64,325.75 (12,202.95) a | 57.87 (2.56) A | 114,847.90 (10,212.70) b | 62.55 (0.73) A | 90,866.62 (9007.32) ab | 56.32 (2.10) A |
| Aboveground herb | 414.04 (119.37) a | 0.37 (0.08) B | 117.22 (8.84) a | 0.06 (0.00) B | 262.08 (96.10) a | 0.16 (0.05) B |
| Forest floor | 16,575.12 (1111.11) a | 15.46 (1.26) C | 24,780.46 (1232.39) b | 13.61 (0.82) C | 19,474.49 (1734.13) a | 12.35 (1.79) C |
| Tree roots | 10,871.25 (2209.40) a | 9.75 (0.55) C | 25,638.54 (2690.03) b | 13.94 (0.48) C | 22,484.83 (2123.27) b | 13.94 (0.45) C |
| Herb roots | 420.96 (112.14) a | 0.38 (0.06) B | 156.36 (12.04) a | 0.09 (0.01) B | 243.45 (80.67) a | 0.15 (0.04) B |
| Mineral soil | 17,140.35 (727.440) a | 16.17 (1.91) C | 17,731.46 (667.12) a | 9.75 (0.54) D | 27,295.93 (936) b | 17.08 (0.80) C |
| Total | 109,747.47 (16,062.27) a | 183,271.83 (14,054.88) b | 160,627.41 (10,247.59) ab | |||
| 22-Year-Old Stand | 32-Year-Old Stand | 42-Year-Old Stand | ||||
|---|---|---|---|---|---|---|
| N Stock (kg hm−2) | Ratio (%) | N Stock (kg hm−2) | Ratio (%) | N Stock (kg hm−2) | Ratio (%) | |
| Aboveground tree | 189.14 (38.62) a | 7.36 (1.02) A | 591.22 (51.90) b | 17.97 (0.95) A | 430. 45 (62.17) b | 12.54 (1.52) A |
| Aboveground herb | 30.87 (11.42) a | 1.21 (0.39) B | 7.95 (0.18) a | 0.24 (0.01) B | 18.79 (7.74) a | 0.54 (0.22) B |
| Forest floor | 710.50 (69.23) a | 28.05 (0.83) C | 950.67 (42.07) a | 29.05 (1.49) C | 739.66 (69.60) a | 21.80 (2.55) C |
| Tree roots | 43.72 (10.22) a | 1.70 (0.29) B | 127.11 (12.23) b | 3.86 (0.23) B | 93.36 (11.97) ab | 2.72 (0.28) B |
| Herb roots | 12.24 (2.87) a | 0.48 (0.09) B | 3.94 (0.08) a | 0.12 (0.01) B | 8.11 (3.26) a | 0.24 (0.09) B |
| Mineral soil | 1537.36 (69.52) a | 61.21 (2.50) D | 1597.77 (52.65) a | 48.76 (1.15) D | 2124.10 (101.40) b | 62.16 (0.64) D |
| Total | 2523.82 (178.50) a | 3278.67 (111.71) b | 3414.49 (129.34) b | |||
| 22-Year-Old Stand | 32-Year-Old Stand | 42-Year-Old Stand | ||||
|---|---|---|---|---|---|---|
| P Stock (kg hm−2) | Ratio (%) | P Stock (kg hm−2) | Ratio (%) | P Stock (kg hm−2) | Ratio (%) | |
| Aboveground tree | 17.82 (4.32) a | 4.00 (0.81) A | 54.57 (3.91) b | 8.86 (0.35) A | 33.33 (3.03) a | 7.04 (0.70) A |
| Aboveground herb | 4.56 (1.66) a | 1.03 (0.35) A | 1.69 (0.14) a | 0.28 (0.02) BD | 2.36 (0.67) a | 0.50 (0.15) B |
| Forest floor | 83.51 (7.46) a | 18.97 (0.92) B | 126.29 (3.61) b | 20.60 (0.86) C | 87.39 (9.53) a | 18.35 (1.66) C |
| Tree roots | 3.22 (0.62) a | 0.72 (0.11) A | 13.76 (1.01) b | 2.24 (0.10) B | 10.51 (1.92) b | 2.19 (0.31) B |
| Herb roots | 2.19 (0.60) a | 0.50 (0.12) A | 0.66 (0.01) a | 0.11 (0.00) D | 1.08 (0.44) a | 0.23 (0.10) B |
| Mineral soil | 327.54 (12.09) a | 74.78 (2.22) C | 417.27 (15.60) a | 67.92 (0.41) E | 340.12 (11.48) a | 71.68 (0.92) D |
| Total | 438.84 (21.32) a | 614.24 (20.47) b | 474.78 (19.22) a | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, J.; Pan, H.; Chen, Z.; Shang, H. Dynamics in Stoichiometric Traits and Carbon, Nitrogen, and Phosphorus Pools across Three Different-Aged Picea asperata Mast. Plantations on the Eastern Tibet Plateau. Forests 2020, 11, 1346. https://doi.org/10.3390/f11121346
Cao J, Pan H, Chen Z, Shang H. Dynamics in Stoichiometric Traits and Carbon, Nitrogen, and Phosphorus Pools across Three Different-Aged Picea asperata Mast. Plantations on the Eastern Tibet Plateau. Forests. 2020; 11(12):1346. https://doi.org/10.3390/f11121346
Chicago/Turabian StyleCao, Jixin, Hong Pan, Zhan Chen, and He Shang. 2020. "Dynamics in Stoichiometric Traits and Carbon, Nitrogen, and Phosphorus Pools across Three Different-Aged Picea asperata Mast. Plantations on the Eastern Tibet Plateau" Forests 11, no. 12: 1346. https://doi.org/10.3390/f11121346