4.1. Natural Regeneration
Small-scale disturbances maintain the old-growth forest structure mainly through the emergence and survival of natural regeneration in the variety of conditions in canopy openings. Total density as well as tree species composition of natural regeneration in species-rich old-growth forest in NNR Sitno corresponds well with the abundance found in other broadleaved-dominated, mixed old-growth forests of the Western Carpathians (e.g., [
39]). In tree species composition,
Acer species (
A. pseudoplatanus and
A. platanoides) are the most abundant except the category of young trees under the canopy.
Acer sp. achieve a high density of regeneration in the positions under the canopy near larger gaps and for survival they need decreased shading achieved mainly through the lower sum of crown projections.
A. pseudoplatanus, however, unlike
A. platanoides clearly survives well in open gaps.
F. excelsior has the highest relative share in the category of young trees under the canopy and the second highest in the same category in gaps.
F. excelsior regenerates and survives well especially in smaller gaps, although in later ages it demands lower sum of crown projections and prefer expanded or even open gap position.
Considering the density of (potentially) seed-producing F. excelsior stems, i.e., the canopy trees with dbh over 20 cm comprising about 10%, the abundance of F. excelsior seedlings and saplings is over-proportionately high. F. sylvatica regenerates especially under the cover of the mature stand near larger gaps, but in a later developmental stage of saplings it may survive better in larger gaps. T. cordata, U. scabra, and C. betulus reveal less clear pattern. These species show the very limited ability to regenerate (low absolute and relative densities) and they ingrow into upper stand layers only in gaps. The higher number of tree species is able to survive initial regeneration stages and ingrowth into the category of young trees in gaps.
Natural regeneration of
Q. petraea is scarce in the long-term. Saniga et al. [
40] have reported the lack of
Q. petraea individuals above the height of 80 cm on all permanent research plots established in NNR Sitno, with the maximum density of natural regeneration 667 individuals per hectare since the late 80-ties of the previous century. Species composition as well as abundance and height structure of natural regeneration are affected not only by gap properties, but also by other factors that are relevant for the survival and height growth of seedlings and saplings. The influence of herbivory may limit the competitive strength of particular species and possibly to change tree species composition [
41,
42,
43]. Therefore, we assume that especially regeneration of light-demanding oak suffers under repeated browsing and is unable to grow out of its competitors.
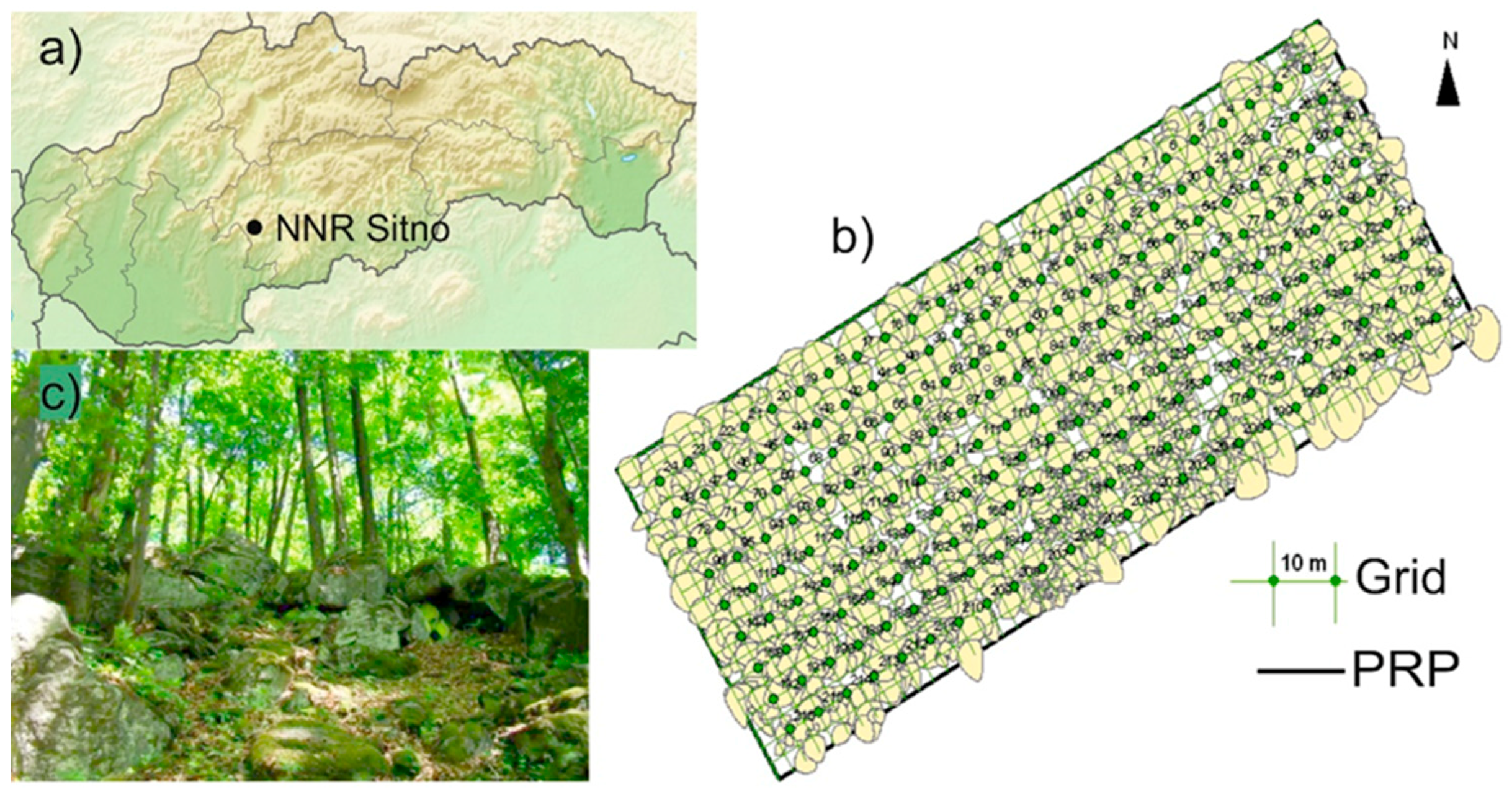
4.2. Disturbance Regime
Many studies are devoted to gap dynamics and to the undoubted effect, which canopy gaps have on regeneration density [
44]. Forest management aimed at the importance of diverse forests relies on that natural disturbances are key drivers of promoting and maintaining tree species diversity [
45]. Gap dynamics of mixed beech-oak old-growth forests characterized by diverse composition of canopy layer are less frequently studied in Central Europe [
11,
31,
46,
47] and provide fragmented base of scientific information. Canopy layer of investigated preserved forest remnant consists of nine tree species with contrasting light requirements (
F. sylvatica,
A. pseudoplatanus,
Q. petraea,
F. excelsior,
T. cordata,
A. platanoides,
A. alba,
C. betulus, and
U. scabra) and possess relatively higher basal area (39.8 m
2 ha
−1) and stem density of trees (603 trees ha
−1). In the investigated forest, mid-scale disturbances (16 expanded gaps >300 m
2) prevail with a low occurrence of disturbances corresponding with very large gaps (two expanded gaps >1000 m
2). Disturbance regime and to this related light availability may thus favour tree species of less shade (as
A. pseudoplatanus or
F. excelsior) and intermediate shade tolerant species (
Q. petraea) to regenerate, establish, grow, and recruit into the forest canopy [
7,
23,
48].
Large proportion of stand area (up to 60%) is under gaps created by the mortality of canopy trees. The mortality of both Q. petraea and F. sylvatica drives the disturbance regime of NNR Sitno, which are the main gapmakers and represent the highest density and basal area among dead trees.
Our results clearly indicate the significant shift in species composition over the time. The Quercus has declined in the recent period. Additionally, massive spread of F. excelsior and associated decrease of F. sylvatica dominance is registered. Results point to significant disproportion between the expected tree species composition corresponding to the habitat, the current tree species composition of the main tree layer and the tree species composition of the next generation.
In the long-term (over 30 years of inventory) the relative density of adult living trees in tree species composition decreases for both
Q. petraea and
F. sylvatica, but more rapidly for
Q. petraea [
40]. This agrees with the study showing increased mortality during 50 years of natural forest development for both
Q. petraea and
F. sylvatica, but stronger for
Q. petraea in Swiss lowlands [
30]. The study from Romanian oak-beech forests also support our results declaring disturbance regime is primarily driven by the mortality of
Q. petraea, showing the absence of this species in the process of tree recruitment [
31].
The largest number of gapmakers is represented by lying deadwood in advanced stage of decomposition (89%), while recently dead trees represent only 7%. Wind has an important role in creating gaps by single or by consecutive disturbance events (only 17% trees died standing). The group mortality prevails in the process of gap formation.
Progressive mortality of trees in separate canopy gaps is the typical feature. Successive extensions of more than a half of gaps resulted in formation of large and very large gaps, which is in agreement with reports from mixed beech forests [
49]. Higher proportion of progressively enlarged gaps (above 2/3) is reported in mixed old-growth oak forests of NNR Kašívárová [
47]. Differences in gap dynamics of beech-oak forest in NNR Sitno (lower frequency of small gaps, absence of very large gaps, and lower proportion of stand area under gaps) in comparison to mixed oak old-growth forest Kašivárová could be attributed to the fact that trees in NNR Sitno grow on less productive volcanic site with high share of rocks. Interestingly, distribution of expanded gaps in NNR Sitno does not follow negative exponential form described in other temperate oak and beech forests [
13,
15,
31,
47], probably due to the recent lack of small gap-scale disturbances. No single gap without extension has been formed recently, suggesting recent changes in disturbance regime.
4.3. Gap Partitioning
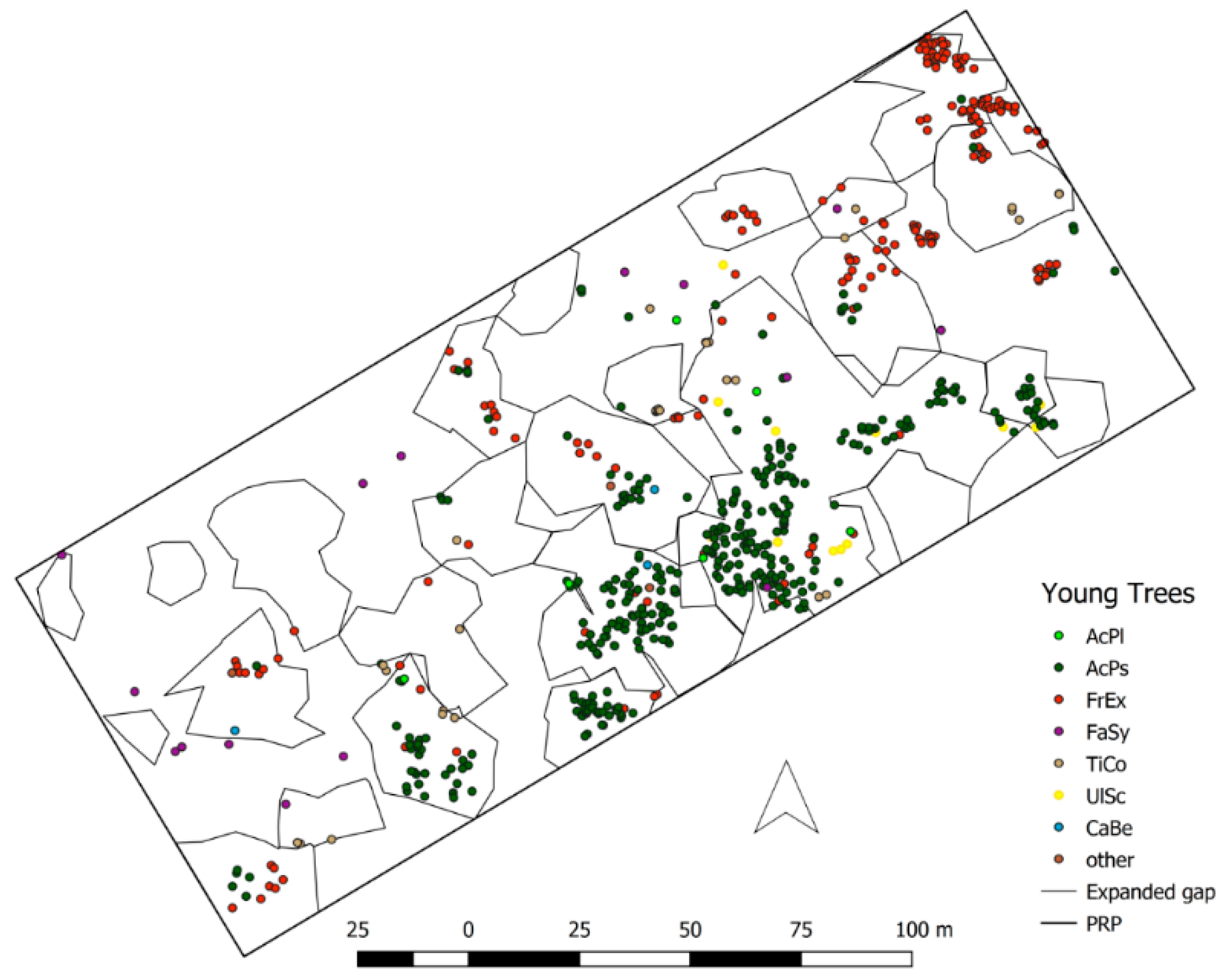
The distribution of natural regeneration corresponds with the distribution of gaps, which agrees with gap-recruitment studies [
44]. Our results suggest that wind disturbances creating successively expanding large gaps do allow maintaining of mid- and late-successional tree species and do not allow early-successional ones and ground vegetation to hamper the regeneration process. Originally, gap partitioning hypothesis assumes that the variety of micro environmental conditions along the gradient shaded understorey—canopy gap enable co-existence of several tree species partly because of higher stem density in gaps [
48]. Different tree species supposedly have different probability of establishing seedlings in particular range of gap sizes [
48].
However, as shown in several studies, gaps not necessarily provide primary environment for regeneration e.g., [
32,
50]. Our results, the higher regeneration density of seedlings observed under the forest canopy compared to gaps, the continuous recruitment, and narrow and linear ordination gradient confirmed by DCA, which suggests relatively homogenous tree species composition on the most plots, confirm these later studies.
The influence of environmental gradient between closed canopy and gap (denoted also as gap-partitioning) in seedling category is not evident for any of the investigated tree species. The variability that can be uniquely attributed to gap and stand related environmental factors is very low. The species composition under the canopy and gaps is very similar—all species can establish also under canopy, even in higher densities due to the proximity of seed trees. At seedling stage (≤50 cm), species relationships and abiotic soil related factors have probably the higher importance than distribution of environmental conditions along stand-gap gradient [
51].
On the other hand, the relative importance of tree species interactions changes with regeneration developmental category [
51]. Our results suggest that environmental gradient is manifested through differential survival rate of different species in saplings and young trees phase. The position under canopy and increasing sum of crown projections reduce the density of saplings. The environment within the gap has significant impact on the tree species composition and positively influences four out of eight studied tree species in sapling category (>50 cm), in agreement with original gap-partitioning theory [
48].
Under the assumption that fluctuations in light/moisture availability and spatial variation in disturbance allow limited number of tree species to coexists [
23], the two most successful gap fillers
A. pseudoplatanus and
F. excelsior and recent shifts in disturbance regime suggest future changes in tree species composition in the investigated nature reserve.
The probability of establishment of
intermediate shade tolerant tree species increases with increasing of gap size [
37]. The presence of two very large gaps (above 1000 m
2) does not create sufficient conditions for
Q. petraea regeneration. Complete lack of regeneration along with the high mortality of old-grown
Q. petraea trees underlines
its decline in NNR Sitno.
Quercus can successfully establish in gaps of sizes between 300–500 m
2, however for further successful growth, saplings require gaps of approximately 1000 m
2 [
11] or even more than 2000 m
2 [
52].
To emphasize the contrasting light requirements of oak in comparison to more shade tolerant species, dendroecological study from oak-beech old-growth forest in Romania demonstrates that the oak successfully establishes after stand replacing disturbance [
29]. This is analogous with establishment of other intermediate shade-tolerant species as
M. obovata in mixed beech forest in western Japan showing patterns suggesting the ability to establish following the large-scale catastrophic disturbance [
53]. In the process of recruitment into the canopy, oak unlike beech is less able to respond with multiple releases to repeating canopy openings. In addition, regeneration of oak is not able to recover once the light conditions in canopy improve.
Therefore, match in the masting of parent oak trees and canopy disturbance events is needed or release of oak saplings at their very young life stage [
4]. It is worth of mentioning that another intermediate shade-tolerant tree species,
T. cordata, responds to disturbance regime in greater densities and shows some potential for continuous recruitment. The recent expansion of
T. cordata,
C. betulus, and
A. pseudoplatanus has been observed in the Bialowieža Primeval Forest in Poland [
19].
Among
less shade tolerant tree species,
F. excelsior greatly benefits from the presence in small gaps. Due to its sensitivity to late spring frost, it avoids exposed sites in the centre of large gaps and regenerates in greater densities in small gaps and under the closed canopy possibly in surroundings of gap edges [
54]. To gap extension, it responds with increased growth succeeding in gap capturing over the
F. sylvatica [
55,
56].
The highest frequency of large expanded gaps (300–1000 m
2) allowed recruitment of less shade tolerant
F. excelsior,
A. pseudoplatanus, and
A. platanoides, which agrees with findings demonstrating that intermediate severity wind disturbances create conditions beneficial for recruitment of less shade tolerant species [
57]. The high densities of natural regeneration of
F. excelsior and
A. pseudoplatanus and patterns of diameter distributions indicating their contemporary establishment suggest that under recent disturbance regime they are the most successful gap fillers. Spatially clumped exclusively in the gap centres, young trees of
A. pseudoplatanus display clear pattern of gap-size partitioning i.e., dependency on environmental gradient. Seedlings regenerate in higher densities in large expanded gaps and oppositely, saplings and young trees are completely absent in small gaps and under closed forest canopy.
Similar to our findings,
A. pseudoplatanus shows evident gap-size partitioning in beech-fir forest reserve in the Dinaric Mountains [
58]. Along with the
A. pseudoplatanus,
A. platanoides is the second most abundant regenerating tree species. Due to its weak competitive ability [
59] it recruits in very low density and frequency of young trees, even lower than for example
U. scabra. Spatio-temporal changes in disturbance regime could have distinct consequences for gap replacement patterns.
Decreases in growth performance of drought-sensitive
F. sylvatica [
60,
61] and recent lack of small gaps may hamper regeneration of
shade tolerant tree species
F. sylvatica on low water retention site. The importance of small gaps illustrates the fact that
F. sylvatica is the only species, whose young trees responded to gaps by 50% decrease relative to forest canopy. This agrees with low (below 10%) relative density and frequency of
F. sylvatica seedlings and saplings and with indicated contemporary difficulties in establishment based on the diameter distribution of trees. Young trees of
F. sylvatica establish under close canopy, prior to gap formation. Based on our results, shade tolerant
F. sylvatica reveals no clear gap partitioning similar to reports for old-growth beech-oak forests in Northern Spain [
62] or beech-fir forests in the Dinaric Mountains [
58]. Analogous patterns of
F. sylvatica establishment—understorey regeneration with successive releases of trees—reports study from the oak-beech forest in Romania [
29].
Our findings confirm that in mixed forests with oak, the more shade tolerant tree species than oak have competitive advantage in recruitment [
31,
63,
64]. Under the canopy and to this related low light availability,
F. sylvatica shows the highest survival,
A. psedoplatanus the lowest and
F. excelsior in between these two species [
65]. In expanded gaps with higher light availability,
A. pseudoplatanus and
F. excelsior show higher height growth rates and allocate more biomass to their stems than
F. sylvatica, which allocates more biomass to branches [
66]. Different growth strategy points to the ability of
F. sylvatica adults to expand and close small canopy gaps by lateral growth of branches [
67], but saplings and young trees to lose with
Acer sp., which growth rates can be twofold higher than those of
Fagus even in small gaps [
22,
48].
Quantification of relationship between the various gap sizes and the related environment along with the spatial arrangement and the species composition of natural regeneration seems not to be simple. Low amount of extracted variability in our study suggests that a regeneration process is influenced by other factors. Beyond the gap size, many factors and gap-related processes generate the mosaic spatial structure of the regeneration and recruitment in mixed forest stands. Differentiation in the regeneration niche (production of seeds—mast seeding, seed dispersal, germination, and establishment of seedlings [
68]), presence, location and abundance of parent trees and seed dispersers, degree of exposed mineral soil and amount of wood debris [
69], micro-environmental factors, including position within gaps [
48], advance regeneration [
70], different life strategies of tree species in accessing the canopy [
22], degree of browsing pressure [
71] above- and below-ground competition among tree species, and climate change [
72], consequently creating new disturbance regimes [
73] can be mentioned.
The composition of natural regeneration, its life stage and spatial structure and distribution of gaps might reveal more clear relationships and allow for quantification of higher amount of variability if initial gap size and time since gap formation is related to density of young trees capturing gap. This highlights the future research to concentrate on repeated gap survey studies [
67] or to combine the retrospective dendroecological approach and static survey. The studies of disturbance regime in various spatiotemporal scales [
6,
74] can allow to more accurately predict recruitment of tree species for forest management purposes.
4.4. Implication to Forest Practice
In close-to-nature forest management, the emphasis on stability and continuity of forest ecosystem results in an effort to retain their structural diversity at small spatial scales. The reliance on natural processes, especially the natural regeneration leads to the use of gap-based regeneration as a fundamental silvicultural technique [
75]. Small-area shelterwood system successfully uses harvest-created gaps to initiate natural regeneration of shade-tolerant species in mixed forest stands. The negative side of this approach is that size-asymmetric competition eliminates light-demanding or slow-growing species during the regeneration phase entirely [
76]. According to tree species composition and gap size distribution found in our study, regeneration in gaps is dominated by shade-tolerant or intermediate species while light-demanding
Q. petraea is rare.
The most common silvicultural practice, to establish and maintain the light-demanding oak recruitment is the regular shelterwood system with short duration of regeneration period. However, recent efforts for minimization of the canopy layer disruption lead to employment of regeneration techniques based on small gaps and complicate thus the oak regeneration considerably [
64]. Moreover, advanced regeneration of shade-tolerant
Acer sp. and
F. excelsior reduces the potential of larger gaps for establishment of
Q. petraea regeneration. Therefore, regeneration requirements of all tree species are often not met with harvest-created gaps alone [
77].
If light-demanding species should be regenerated, while the mixed ecosystem should be established and maintained, lags in regeneration of shade tolerant species following creation of large gaps (at least 0.2 ha) would create better situation for light-demanding Q. petraea. The selection of shade-tolerant A. pseudoplatanus and platanoides in the first (seed) cutting contribute to reducing their seed bank in favour of oak. After oak is successfully established, its overtopping should be prevented by combination of fast progressing of gaps enlargement and subsequent control of fast-growing Acer sp., Fagus and Fraxinus recruitment.
The results of our study confirm that to regenerate the mixtures of Acer, Fagus and Fraxinus does not represent a complicated task. Subsequent changes in the proportions of separate tree species in the thicket stage (i.e., saplings and young trees in our study) are dependent on the decision of the forest manager, with the possible modifications caused by the other factors, especially by the selective damage. On the other hand, the maintenance of Quercus must be thoroughly planned in advance—already at the identification of suitable conditions for location of the gaps, the determination of their size and the realization of seed cutting therein. The duration of regeneration period should by last 40 years, whereby the duration of regeneration periods within groups should be differentiated: 15 years for Q. petraea and 25 years period for Acer, Fagus and Fraxinus.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









