1. Introduction
Forests are believed to provide a large set of ecosystem services in addition to wood and biomass production [
1,
2,
3]. Climate change can reduce, or at least alter, quality and quantity of the ecosystem services that forests provide, thus affecting resilience and cultural identity of local communities [
4,
5,
6]. It is, therefore, of primary importance to develop and use reliable and adequate climatic surfaces for the purpose [
7,
8,
9]. This would impact on the possibility to derive useful instruments for policymakers able to anticipate changes in forest ecosystems’ functionality and their capacity to deliver services [
10]. Ecosystem services of forests are related to specific functions, such as growth and carbon stock into woody tissues, carbon sequestration and oxygen enrichment via photosynthesis, regeneration, element cycling, soil development, biodiversity holding, and so on. For this reason, a wide set of attributes should be taken into account to evaluate the overall functionality of forests and their capacity to provide ecosystem services [
11,
12].
The spatial distribution of forest tree species (range) is often recognized as a good proxy to derive their ecological requirements and to build up statistical models [
9,
13]. Analysis of field data and tree-level information allows a comprehensive understanding of stand-level health as well as the impact of environmental factors on tree growth dynamics [
14,
15,
16,
17]. The growth rate (increment) of forest tree-species and their standing biomass is a fundamental indicator to evaluate their health and stability across space and time [
18,
19,
20]. This aspect is also directly connected to carbon sequestration and climate regulation (regulatory services) and not only to the economic value of the forest (provisional services). Growth integrates the physiological condition of trees and the associated changes are a reliable indicator of climate impact [
21]. Therefore, tree growth is a key variable evaluating the ability of forests to adapt to a changing climate, to mitigate climate change and to provide valuable ecosystem services [
22].
Expected impacts of climate change on forest ecosystems’ health include shifted tree species distributions, a lower growth rate and changes in species composition and related spatial arrangement. These processes are mainly associated with rising mean temperatures, variations in precipitation regimes, longer drought conditions and the increasing frequency of extreme weather events [
6,
23,
24,
25,
26]. Europe-wide reduction in primary productivity caused by heat waves and droughts was already detected in 2003 [
27]. More recently, a widespread tree mortality has been observed in forests around the globe [
28,
29]. Temperature extremes, precipitation extremes and wind throws have been pointed out as the most important climatic and atmospheric stressors for forest health [
25,
30].
Under climate change, the complexity of forest ecosystems demands the development of a prompt and effective system for detecting the impacts of pressure factors. In Europe, such activities are carried out routinely by means of terrestrial surveys [
31,
32] within the International Co-operative Program on ‘Assessment and Monitoring of Air Pollution Effects on Forests’ (ICP Forests), which is the most comprehensive European program for large-scale detection of forest ecosystem health [
33,
34]. An interdisciplinary research agenda integrated with monitoring networks and modelling is needed to provide information at all levels of decision making, from policy development to the management unit [
25,
35,
36].
The European beech (
Fagus sylvatica L.) is one of the most frequently assessed deciduous tree species within the ICP Forests monitoring program. This tree can be found on both Level I (extensive network) and Level II (intensive network) plots along the North-South and West-East gradients in Europe ranging from Southern Scandinavia to Southern Italy, and from the Atlantic coast of Northern Spain to the Bulgarian Black Sea coast. A total of 87 beech Level II plots wide spread over Europe are currently being monitored. The relevance of this species in the scientific literature is also well known, given its sensitivity to climatic fluctuations and spatial extent [
37,
38]. The impact of climate change on the geographic distribution of European beech was explored by means of dendrochronological studies [
39,
40], phenological methods [
41], species distribution modelling techniques and process-based approaches [
13,
42].
Since the Italian peninsula represents the rear-edge for the species, many ecological studies were carried out in the last decades on beech forests. Elevation was significantly correlated with dendrochronological variables along the Alps and Northern Apennines and this correlation diverges in central-southern Apennines [
43]. Summer drought was recognized to be the most influencing driver with different effects at different elevations [
39,
40]. Studies on old-growth forests revealed the roles of disturbance, competition and climate in structuring tree stands [
39,
44]. Taken together, earlier studies indicate that climate change will impact adaptive capacity and spatial structure of forests along both latitudinal and elevation gradients.
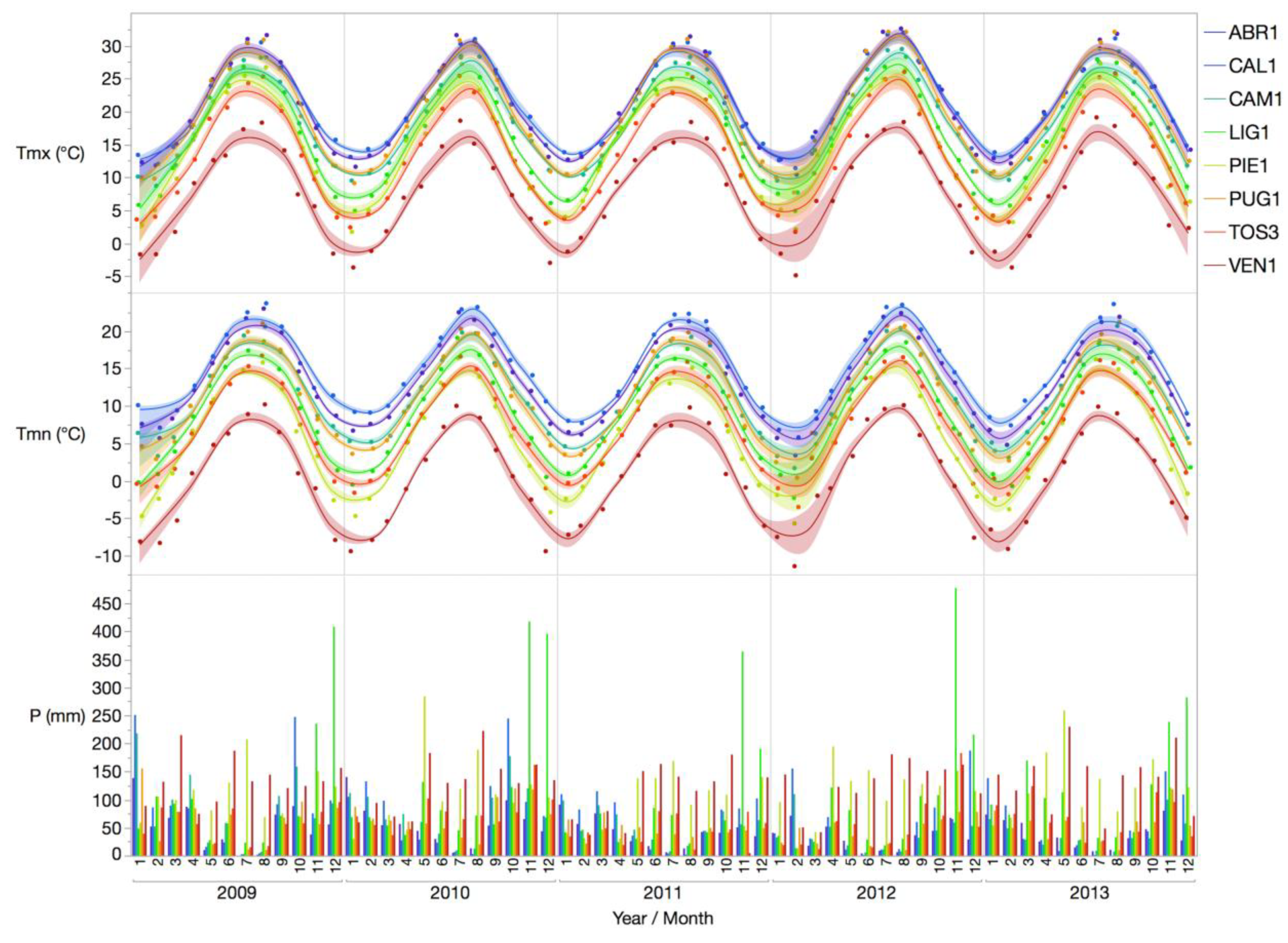
In this paper, a modelling analysis is proposed, with the aim to evaluate the intra-annual growth rate of European beech and identify possible climate-growth interactions. The species was monitored across eight sites distributed along the whole Italian peninsula. Eight 5-year inter-annual time series datasets on tree growth within the Italian ICP-Forests Level II intensive monitoring network were evaluated in correlation with local climatic variables. Statistical techniques were applied to evaluate the influence of the main climatic variables on beech growth trends, to assess whether ecological conditions affected local growth and where (which plots) a potential source of adaptation to climatic drivers (i.e., growth dynamics not significantly affected by climate variables), could be detected. Finally, different climate change scenarios were applied to predict future development at each site.
4. Discussion
The spatial arrangement and growth trend of forest systems are strictly related to management, which influences tree density and vertical/horizontal stand structure. In such a context, GAMM analysis showed how climate was able to explain just a reduced amount of variability across plots. In this case, forest management, i.e., spatial structure and standing volume (not included in our model) might be looked at as the basic drivers for the growth of the analyzed Fagus sylvatica trees across Italy.
In a changing environment, accounting for forest response to climate changes is functional to sustainable forest management. Time series represent the time-evolution of the meteorological dynamic process and are fundamental to evaluate patterns and responses of forest species to climate change [
26]. The repeated, short-term occurrence of anomalous seasons or extreme events make each year potentially sensitive to climate deviations and therefore even limited time series may provide useful insights today. Many environmental and modelling studies are focused on long-term climatic averages to determine the effects of climate change on forest populations. However, in cases of databases with detailed spatio-temporal resolution, the presence of missing values in a dataset can heavily affect any kind of analysis, leading to the underestimation of natural processes [
45]. An increasing degree of representativeness and precision in statistical techniques to data analysis is needed in studies requiring a higher temporal-resolution, such as dendrochronology, or studies based on seasonal effects on plant growth [
17].
Results highlighted latent growth patterns in beech populations along the latitude gradient in Italy, providing an example of the possible use of joint mensurational and climatic high-resolution time series. However, the low variance explained by GAMM was an expected result of this study. Indeed, while most of the variability on growth and stand structure is often driven by climate in non-disturbed stands [
9,
20,
57,
58] the same does not hold true for the ICP-Forests network. The analyzed dataset is composed by different forest categories including both coppices (TOS3, LIG1), grown-up high forests (ABR1, CAL1, CAM1, PIE1, PUG1, VEN1) with different structures and responses driven by age [
32]. In light of this, our results should be interpreted as a starting point for further investigation. More informative and complete models could actually be built with ICP-Forests datasets including, for instance, soil variables or structural indices which might improve the amount of variance explained in GAMM models. However, we must keep in mind thatthis network is mainly made by managed forest stands, chosen by experts in order to monitor stands’ dynamics across a long monitoring period in specific environmental contexts. Consequently, results should be evaluated in the framework of the ICP-Forests only and statistical inference would be just speculative.
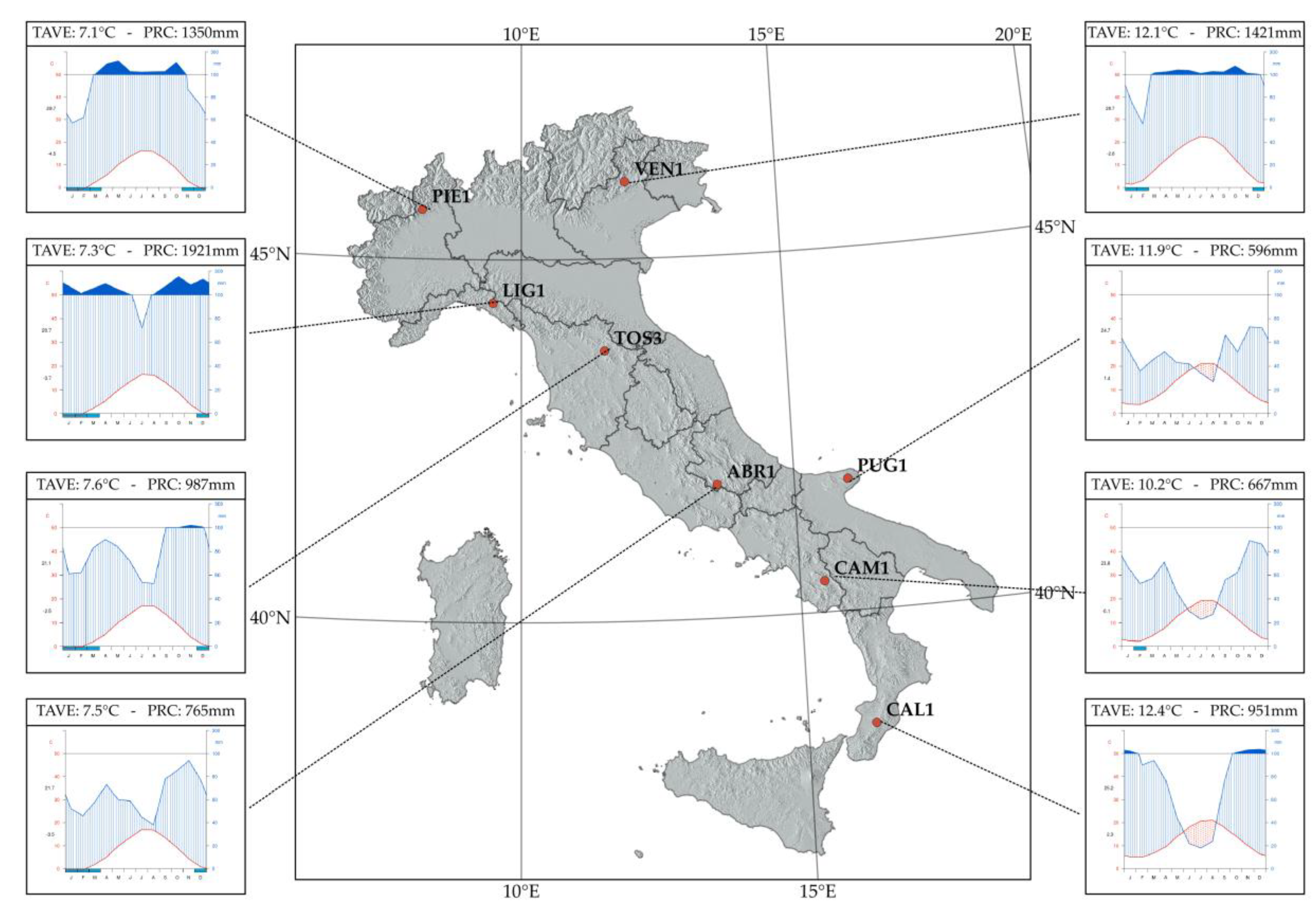
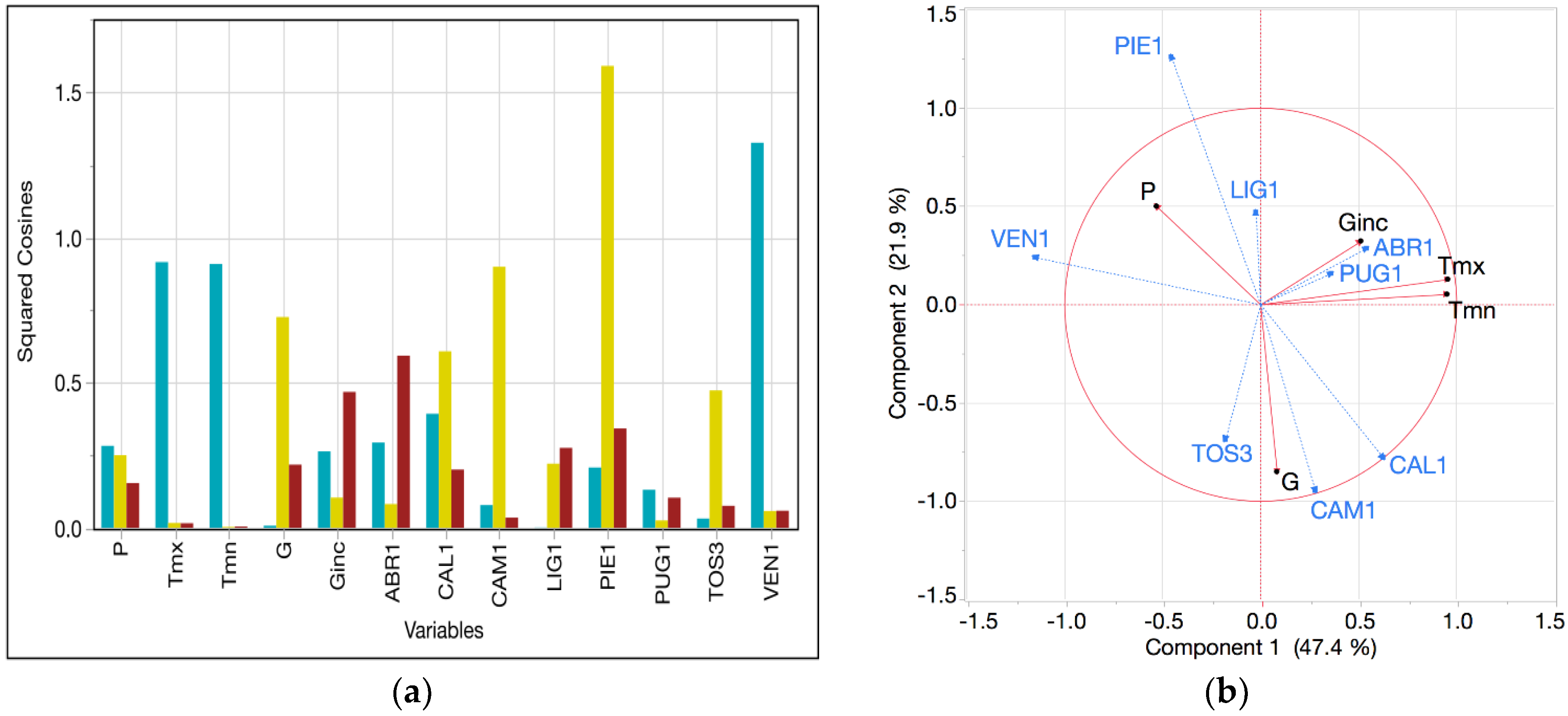
Specific outcomes of this study are particularly interesting in light of forest ecology. For instance, the high variance explained by GAMM models in LIG1 might be motivated with the particular environmental condition of the area. In actuality, LIG1 is the first Italian ICP-Forests plot (from North to South) with a small reduction of rainfall over the growing season. The high water availability in a warmer condition than areas located at higher latitudes might influence the growing period without summer stress and a relatively stable climate. Interestingly, the spatially closer plot TOS3 is characterized by different predictors (only SPEI and P were significant) and a peculiar climatic diagram (see
Figure 1). The spatial proximity between these two sites, in combination with the results of our study, pointed out the inherent environmental and biological variability across the Mediterranean basin even at small spatial scales and with an (almost) genetically stable forest tree species like European beech [
59,
60]. Accounting for forest response to climate shift is functional to an adaptive forest management in a changing environment. In this regard, high temporal resolution long-term time series might reveal growth patterns which allow identification of dynamic meteorological-growth relationships [
54]. This information is a basic requirement for the ICP-Forests framework, where biotic and abiotic disturbances are being monitored. Moreover, high-quality and long-term time series are increasingly needed for representing meteorological-growth relationship dynamics and fundamental for the evaluation of both processes and responses of forest species to a shifting climate. Even if this issue can be seen as a weakness of this paper, useful information could be read from even a 5-year time series which could drive future monitoring efforts. It is well known that extreme events have been shown to have a crucial role for forest systems and plant communities in the Mediterranean areas [
3]. Under future climate changes, such events will play an even more critical role. While the proposed approach can only indirectly outline the importance of climatic extremes in beech forest dynamics, unpredictable events will require longer time-series analysis and refined statistical modelling to understand their effects on forest ecosystems [
27]. However, GAMM and dendrometers could be seen as valuable tools for the purpose.
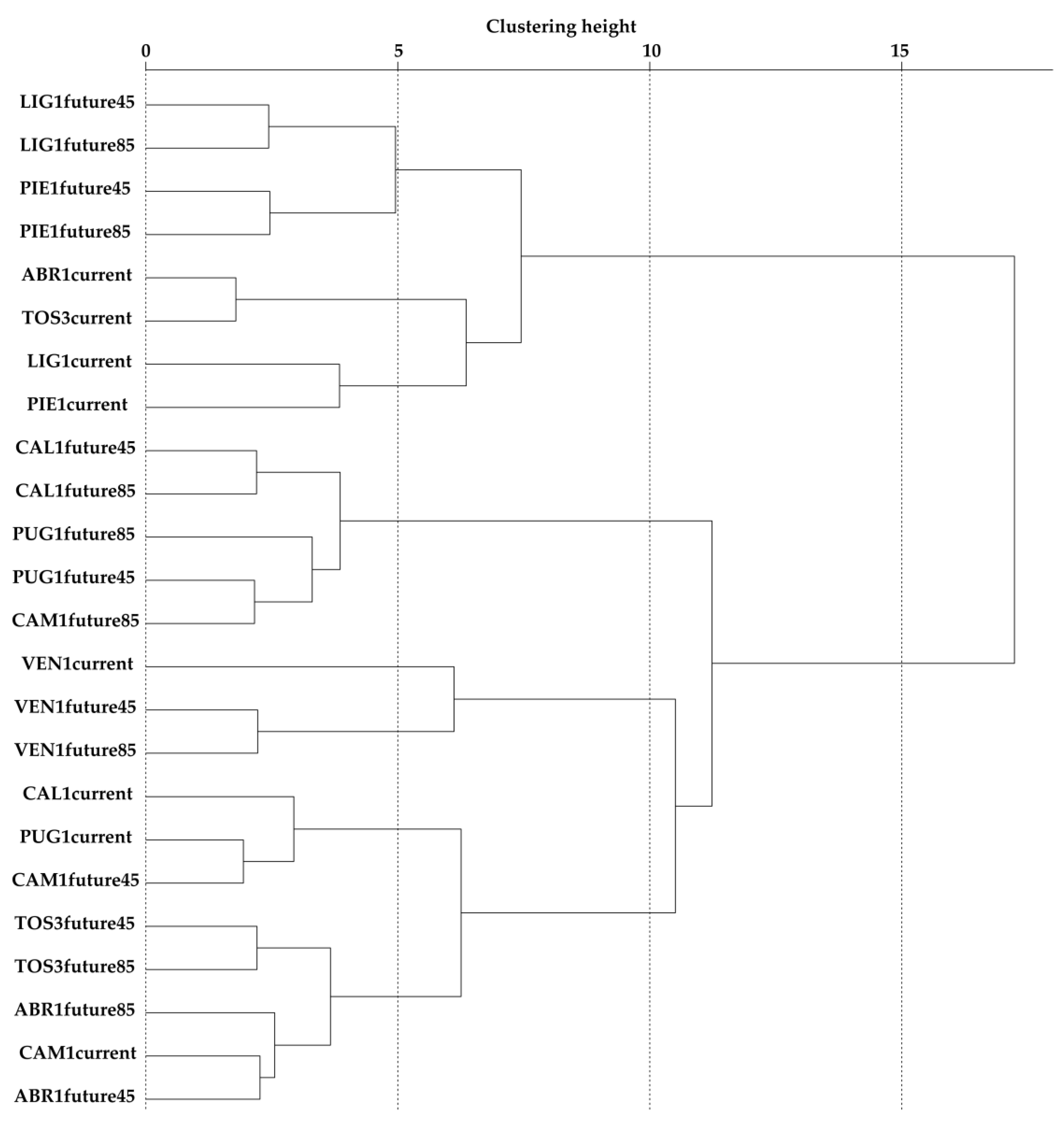
Concerning future scenarios, VEN1 plot represents a peculiar case-study where the current climate and future scenarios seemed to be totally disconnected from all the other beech plots of the Italian monitoring network. Here, local climate regimes might be seen as an important driver to be monitored carefully in the future. An adaptive response of functional traits at the leading and trailing edge of the current distribution will help researchers in understanding possible adaptive measures to balance forest management strategies. Indeed, the central-peripheral adaptive trend will probably play a key role in the framework of a changing climate as an adaptive response of functional traits of small populations, i.e., the marginal and peripheral forest populations [
59,
60,
61].
Moreover, the bimodal growth of some Mediterranean forest tree species has the potential to affect dendrochronological analysis based on annual tree ring width [
17,
20]. Knowledge of long-term climate-growth relationships is, therefore, a basic step to address suited future forest management strategies and tackle effectively both climate change-induced effects and human-related disturbances [
62,
63]. In this respect, a large-scale, intensive monitoring network may provide valuable information about tree growth responses across the annual variability of climate at both latitudinal and elevation gradients.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}