Presence of Mycorrhizal Fungal Hyphae Rather than Living Roots Retards Root Litter Decomposition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Material Collection and Preparation
2.2. Pot Experiment
2.3. Initial Chemical Traits of Root Litter
2.4. Enzyme Assay
2.5. Mycorrhizal Colonization of Seedling Roots
2.6. Calculation and Statistical Analysis
3. Results
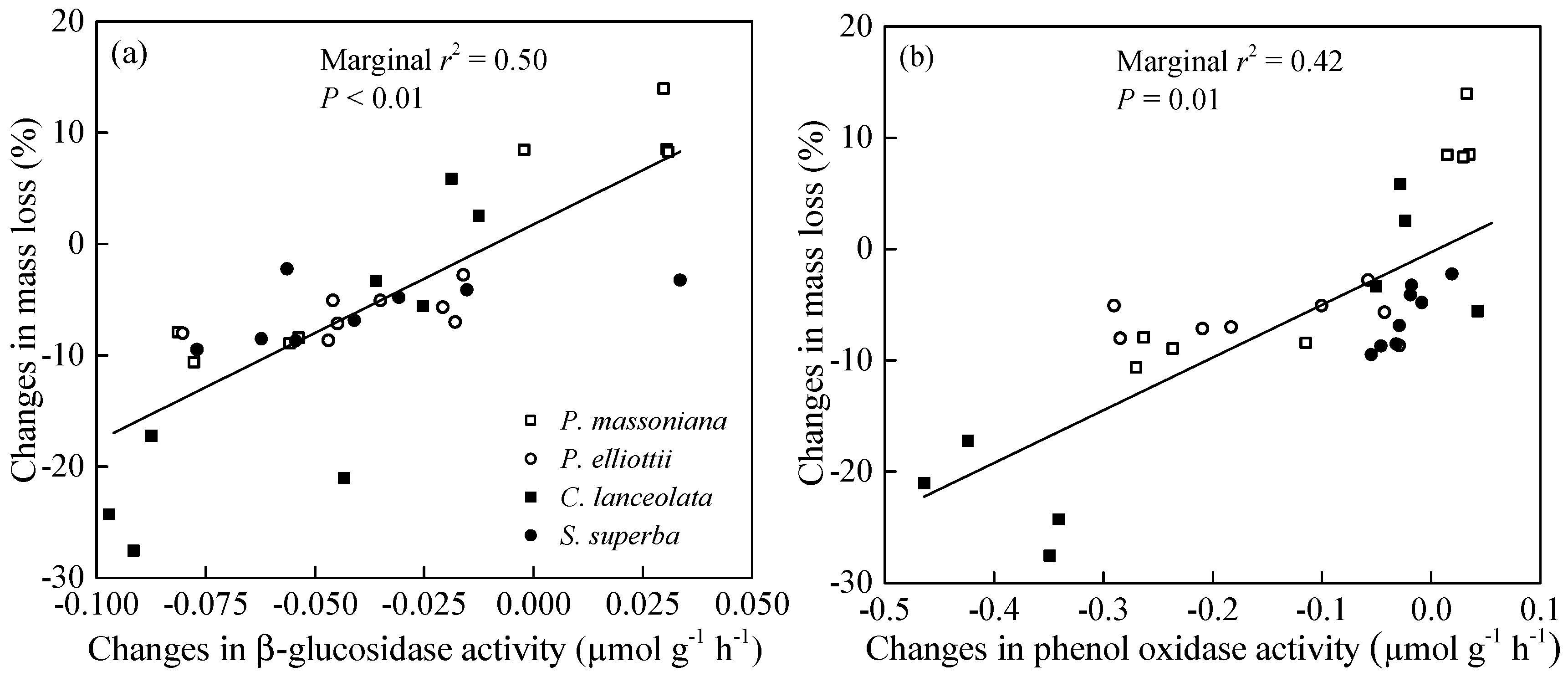
3.1. Initial Chemical Traits and Their Relationships with Root Litter Decomposition
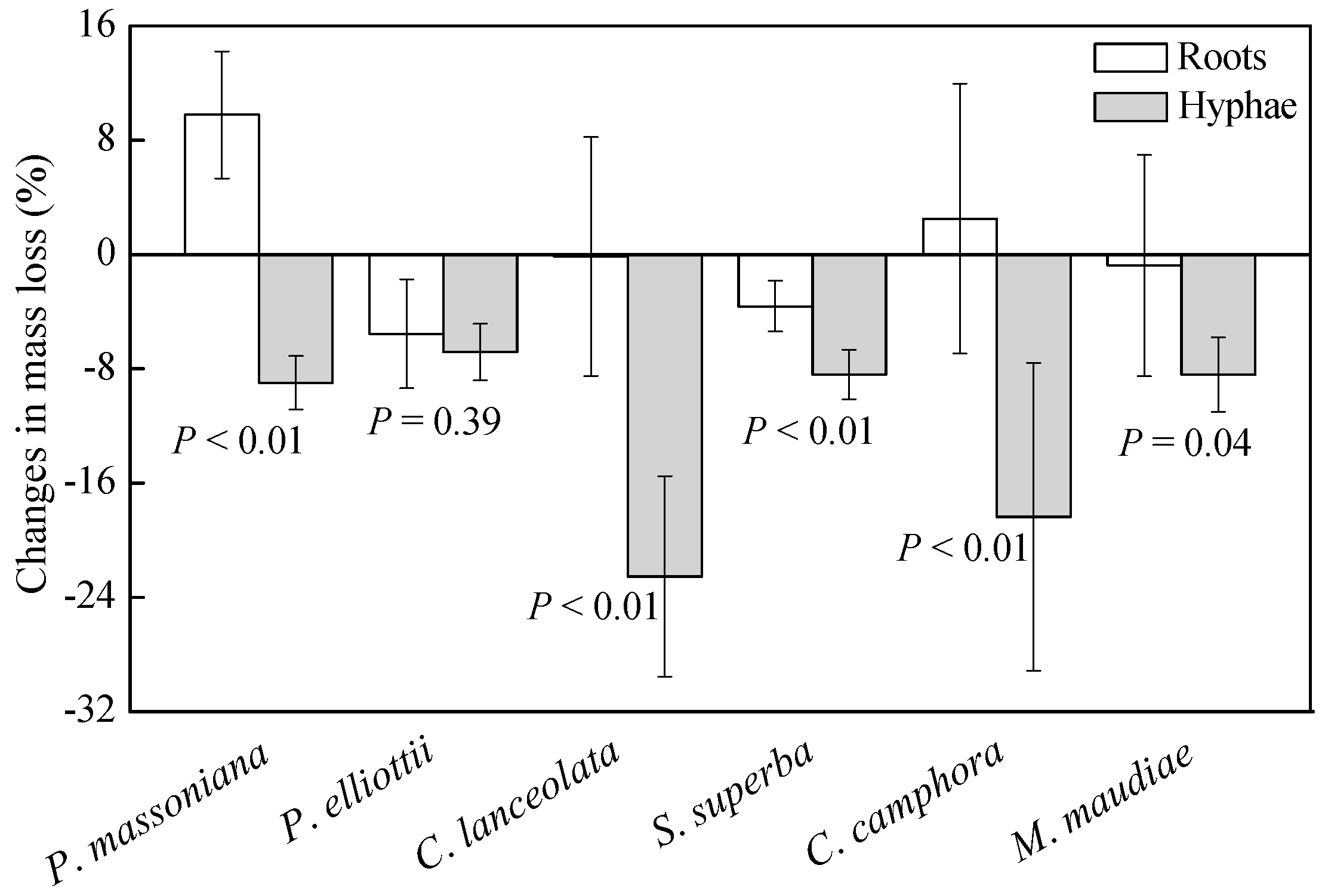
3.2. Relative Effects of Living Roots and Mycorrhizal Hyphae on Root Litter Decomposition
4. Discussion
4.1. Relative Effects of Living Roots and Mycorrhizal Hyphae on Root Litter Decomposition
4.2. Mycorrhizal Type Associated Mycorrhizal Fungal Effects on Root Litter Decomposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silver, W.L.; Miya, R.K. Global patterns in root decomposition: Comparisons of climate and litter quality effects. Oecologia 2001, 129, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Keiblinger, K.M.; Schmid, E. Who is who in litter decomposition? Metaproteomics reveals major microbial players and their biogeochemical functions. ISME J. 2012, 6, 1749–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, C.W.; Kennedy, P.G. Revisiting the ‘Gadgil effect’: Do interguild fungal interactions control carbon cycling in forest soils? New Phytol. 2016, 209, 1382–1394. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, E.; Pena, R.; Fernandez, C.W.; Soong, J.L. Mycorrhizal interactions with saprotrophs and impact on soil carbon storage. In Mycorrhizal Mediation of Soil; Johnson, N.C., Gehring, C., Jansa, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 441–460. [Google Scholar]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Averill, C.; Hawkes, C.V. Ectomycorrhizal fungi slow soil carbon cycling. Ecol. Lett. 2016, 19, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Brzostek, E.R.; Dragoni, D.; Brown, Z.A.; Phillips, R.P. Mycorrhizal type determines the magnitude and direction of root-induced changes in decomposition in a temperate forest. New Phytol. 2015, 206, 1274–1282. [Google Scholar] [CrossRef]
- Wang, F.; Fang, X.; Ding, Z.; Wan, Z.; Chen, F. Effects of understory plant root growth into the litter layer on the leaf litter decomposition of two woody species in a subtropical forest. For. Ecol. Manag. 2016, 364, 39–45. [Google Scholar] [CrossRef]
- Prieto, I.; Birouste, M.; Zamora-Ledezma, E.; Gentit, A.; Goldin, J.; Volaire, F.; Roumet, C. Decomposition rates of fine roots from three herbaceous perennial species: Combined effect of root mixture composition and living plant community. Plant Soil 2017, 415, 359–372. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Factors affecting rhizosphere priming effects. J. Plant Nutr. Soil Sci. 2002, 165, 382–396. [Google Scholar] [CrossRef]
- Saar, S.; Semchenko, M.; Barel, J.M.; De Deyn, G.B. Legume presence reduces the decomposition rate of non-legume roots. Soil Biol. Biochem. 2016, 94, 88–93. [Google Scholar] [CrossRef]
- Gadgil, R.; Gadgil, P.D. Mycorrhiza and litter decomposition. Nature 1971, 233, 133. [Google Scholar] [CrossRef] [PubMed]
- Koide, R.T.; Wu, T. Ectomycorrhizas and retarded decomposition in a Pinus resinosa plantation. New Phytol. 2003, 203, 401–407. [Google Scholar] [CrossRef]
- Kaiser, C.; Killburn, M.R.; Clode, P.L.; Fuchslueger, L.; Koranda, M.; Cliff, J.B.; Solaiman, Z.M.; Murphy, D.V. Exploring the transfer of recent plant photosynthates to soil microbes: Mycorrhizal pathway vs direct root exudation. New Phytol. 2015, 205, 1537–1551. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, J.; Yuan, Y.; Zhao, C.; Liu, Q.; Yin, H. Mycelium- and root-derived C inputs differ in their impacts on soil organic C pools and decomposition in forests. Soil Biol. Biochem. 2018, 123, 257–265. [Google Scholar] [CrossRef]
- Meier, I.C.; Pritchard, S.G.; Brzostek, E.R.; McCormack, L.; Phillips, R.P. The rhizosphere and hyphosphere differ in their impacts on carbon and nitrogen cycling in forests exposed to elevated CO2. New Phytol. 2015, 205, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, B.D.; Tunlid, A. Ectomycorrhizal fungi–potential organic matter decomposers, yet not saprotrophs. New Phytol. 2015, 205, 1443–1447. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.P.; Brzostek, E.; Midgley, M.G. The mycorrhizal-associated nutrient economy: A new framework for predicting carbon-nutrient couplings in temperate forests. New Phytol. 2013, 199, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; McCormack, M.; Ma, C.; Guo, D. Similar below-ground carbon cycling dynamics but contrasting modes of nitrogen cycling between arbuscular mycorrhizal and ectomycorrhizal forests. New Phytol. 2017, 213, 1440–1451. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Wheeler, E.; Phillips, R.P. Root-induced changes in nutrient cycling in forests depend on exudation rates. Soil Biol. Biochem. 2014, 78, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Lin, G.; Guo, D.; Li, L.; Ma, C.; Zeng, D. Contrasting effects of ectomycorrhizal and arbuscular mycorrhizal tropical tree species on soil nitrogen cycling: The potential mechanisms and corresponding adaptive strategies. Oikos 2018, 127, 518–530. [Google Scholar] [CrossRef]
- Li, C.; Li, Q.; Qiao, N.; Xu, X.; Li, Q.; Wang, H. Inorganic and organic nitrogen uptake by nine dominant subtropical tree species. iForest 2015, 9, 253–258. [Google Scholar] [CrossRef]
- Lin, G.; Zeng, D. Heterogeneity in decomposition rates and annual litter inputs within fine-root architecture of tree species: Implications for forest soil carbon accumulation. For. Ecol. Manag. 2017, 389, 386–394. [Google Scholar] [CrossRef]
- Johnson, D.; Leake, J.R.; Read, D.J. Novel in-growth core system enables functional studies of grassland mycorrhizal mycelial networks. New Phytol. 2001, 152, 555–562. [Google Scholar] [CrossRef]
- Bradford, M.A.; Tordoff, G.; Eggers, T.; Jones, H.; Newington, J. Microbiota, fauna, and mesh size interactions in litter decomposition. Oikos 2002, 99, 317–323. [Google Scholar] [CrossRef]
- Ryan, M.G.; Melillo, J.M.; Ricca, A. A comparison of methods for determining proximate carbon fractions of forest litter. Can. J. For. Res. 1990, 20, 166–171. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.L.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular–arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference. R package version 1.40.4. Available online: https://cran.r-project.org/web/packages/MuMIn/index (accessed on 30 January 2018).
- Gadgil, R.; Gadgil, P.D. Suppression of litter decomposition by mycorrhizal roots of Pinus radiata. N. Z. J. For. Sci. 1975, 5, 33–41. [Google Scholar]
- Paterson, E.; Sim, A.; Davidson, J.; Daniell, T.J. Arbuscular mycorrhizal hyphae promote priming of native soil organic matter mineralisation. Plant Soil 2016, 408, 243–254. [Google Scholar] [CrossRef]
- Ernfors, M.; Rütting, T.; Klemedtsson, L. Increased nitrous oxide emissions from a drained organic forest soil after exclusion of ectomycorrhizal mycelia. Plant Soil 2011, 343, 161–170. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Turner, B.L.; Winter, K.; Chamberlain, P.M.; Stott, A.; Tanner, E.V.J. Root and arbuscular mycorrhizal mycelial interactions with soil microorganisms in lowland tropical forest. FEMS Microbiol. Ecol. 2013, 85, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Olsson, P.A.; Bååth, E.; Jakobsen, I.; Söderström, B. Soil bacteria respond to presence of roots but not to mycelium of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 1996, 28, 463–470. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Cheng, W.X. Interactions between soil and tree roots accelerate long-term soil carbon decomposition. Ecol. Lett. 2007, 10, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Godbold, D.L.; Hoosbeek, M.R.; Lukac, M.; Cotrufo, M.F.; Janssens, I.A.; Ceulemans, R.; Polle, A.; Velthorst, E.J.; Scarascia-Mugnozza, G.; De Angelis, P.; et al. Mycorrhizal hyphal turnover as a dominant process for carbon input into soil organic matter. Plant Soil 2006, 281, 15–24. [Google Scholar] [CrossRef]
- Staddon, P.L.; Ramsey, C.B.; Ostle, N.; Ineson, P.; Fitter, A.H. Rapid turnover of hyphae of mycorrhizal fungi determined by AMS microanalysis of 14C. Science 2003, 300, 1138–1140. [Google Scholar] [CrossRef] [PubMed]
- Högberg, M.N.; Högberg, P. Extramatrical ectomycorrhizal mycelium contributes one-third of microbial biomass and produces, together with associated roots, half the dissolved organic carbon in soil. New Phytol. 2002, 154, 791–795. [Google Scholar] [CrossRef]
- Hobbie, J.E.; Hobbie, E.A. 15N in symbiotic fungi and plants estimates nitrogen and carbon flux rates in arctic tundra. Ecology 2006, 87, 816–822. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.A.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, E.; Jansa, J.; Hammer, E.C.; Rillig, M.C. Do arbuscular mycorrhizal fungi stabilize litter-derived carbon in soil? J. Ecol. 2016, 104, 261–269. [Google Scholar] [CrossRef]
- Jones, D.L.; Hodge, A.; Kuzyakov, Y. Plant and mycorrhizal regulation of rhizodeposition. New Phytol. 2004, 163, 459–480. [Google Scholar] [CrossRef]
- Meier, I.C.; Avis, P.G.; Phillips, R.P. Fungal communities influence root exudation rates in pine seedlings. FEMS Microbiol. Ecol. 2013, 83, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.; Naik, D.; Cumming, J.R. The influence of phosphorus availability and Laccaria bicolor symbiosis on phosphate acquisition, antioxidant enzyme activity, and rhizospheric carbon flux in Populus tremuloides. Mycorrhiza 2014, 24, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Wurzburger, N.; Brookshire, E.N.J. Experimental evidence that mycorrhizal nitrogen strategies affect soil carbon. Ecology 2017, 98, 1491–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheeke, T.E.; Phillips, R.P.; Brzostek, E.R.; Rosling, A.; Bever, J.D.; Fransson, P. Dominant mycorrhizal association of trees alters carbon and nutrient cycling by selecting for microbial groups with distinct enzyme function. New Phytol. 2017, 214, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Koele, N.; Dickie, I.A.; Oleksyn, J.; Richardson, S.J.; Reich, P.B. No globally consistent effect of ectomycorrhizal status on foliar traits. New Phytol. 2002, 196, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Mucha, J.; Zadworny, M.; Werner, A. Cytoskeleton and mitochondrial morphology of saprotrophs and the pathogen Heterobasidion annosum in the presence of Suillus bovinus metabolites. Mycol. Res. 2009, 113, 981–990. [Google Scholar] [CrossRef]
- Trap, J.; Akpa-Vinceslas, M.; Margerie, P.; Boudsocq, S.; Richard, F.; Decaëns, T.; Aubert, M. Slow decomposition of leaf litter from mature Fagus sylvatica trees promotes offspring nitrogen acquisition by interacting with ectomycorrhizal fungi. J. Ecol. 2017, 105, 528–539. [Google Scholar] [CrossRef]
- Jannoura, R.; Kleikamp, B.; Dyckmans, J.; Joergensen, R.G. Impact of pea growth and arbuscular mycorrhizal fungi on the decomposition of 15N-labeled maize residues. Biol. Fertil. Soils 2012, 48, 547–560. [Google Scholar] [CrossRef]
- Leifheit, E.F.; Verbruggen, E.; Rillig, M.C. Arbuscular mycorrhizal fungi reduce decomposition of woody plant litter while increasing soil aggregation. Soil Biol. Biochem. 2015, 81, 323–328. [Google Scholar] [CrossRef]
- Welc, M.; Ravnskov, S.; Kieliszewska-Rokicka, B.; Larsen, J. Suppression of other soil microorganisms by mycelium of arbuscular mycorrhizal fungi in root-free soil. Soil Biol. Biochem. 2010, 42, 1534–1540. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, M.G.; Liu, Y.; Zhang, F.S.; Hodge, A.; Feng, G. Carbon and phosphorus exchange may enable cooperation between an arbuscular mycorrhizal fungus and a phosphate-solubilizing bacterium. New Phytol. 2016, 210, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.S.; Niklas, K.J.; Liu, Y.; Fang, X.M.; Wan, S.Z.; Wang, H.M. Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age. Tree Physiol. 2015, 35, 1106–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Mycorrhizal Type | C (%) | N (mg g−1) | OSF (%) | ASF (%) | AIF (%) | C:N Ratio | AIF:N Ratio |
|---|---|---|---|---|---|---|---|---|
| Pinus massoniana | ECM | 53.8 | 11.2 | 16.0 | 37.2 | 46.8 | 48.0 | 41.8 |
| Pinus elliottii | ECM | 46.4 | 9.5 | 11.1 | 49.8 | 39.1 | 48.8 | 41.2 |
| Cunninghamia lanceolata | AM | 57.3 | 15.1 | 11.2 | 28.7 | 60.0 | 38.0 | 39.8 |
| Schima superba | AM | 52.8 | 13.2 | 13.7 | 35.2 | 51.0 | 40.0 | 38.7 |
| Cinnamomum camphora | AM | 50.0 | 15.7 | 9.6 | 33.2 | 57.2 | 31.9 | 36.5 |
| Michelia maudiae | AM | 50.0 | 16.2 | 9.8 | 31.9 | 58.3 | 30.9 | 36.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, G.; Chen, Z.; Zeng, D.-H. Presence of Mycorrhizal Fungal Hyphae Rather than Living Roots Retards Root Litter Decomposition. Forests 2019, 10, 502. https://doi.org/10.3390/f10060502
Lin G, Chen Z, Zeng D-H. Presence of Mycorrhizal Fungal Hyphae Rather than Living Roots Retards Root Litter Decomposition. Forests. 2019; 10(6):502. https://doi.org/10.3390/f10060502
Chicago/Turabian StyleLin, Guigang, Zhengxia Chen, and De-Hui Zeng. 2019. "Presence of Mycorrhizal Fungal Hyphae Rather than Living Roots Retards Root Litter Decomposition" Forests 10, no. 6: 502. https://doi.org/10.3390/f10060502