Whitebark Pine Recruitment in Sierra Nevada Driven by Range Position and Disturbance History

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plot-Based Model Development
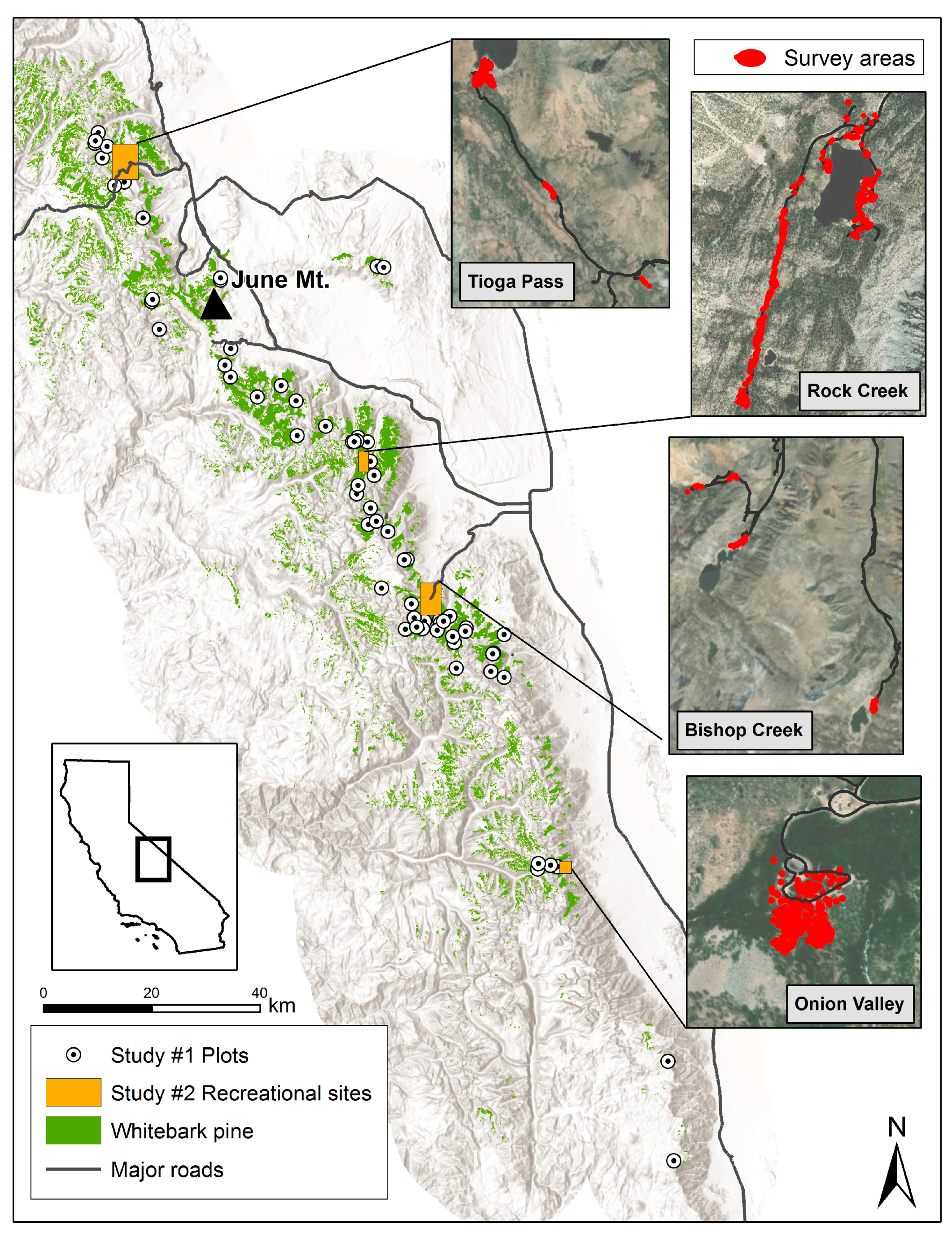
2.1.1. Field Sampling Design
2.1.2. Model Development
2.1.3. Model Evaluation
2.2. Stem Density in Recreational Sites
2.3. Observational Case Study in Mountain Pine Beetle Attack at June Mt.
3. Results
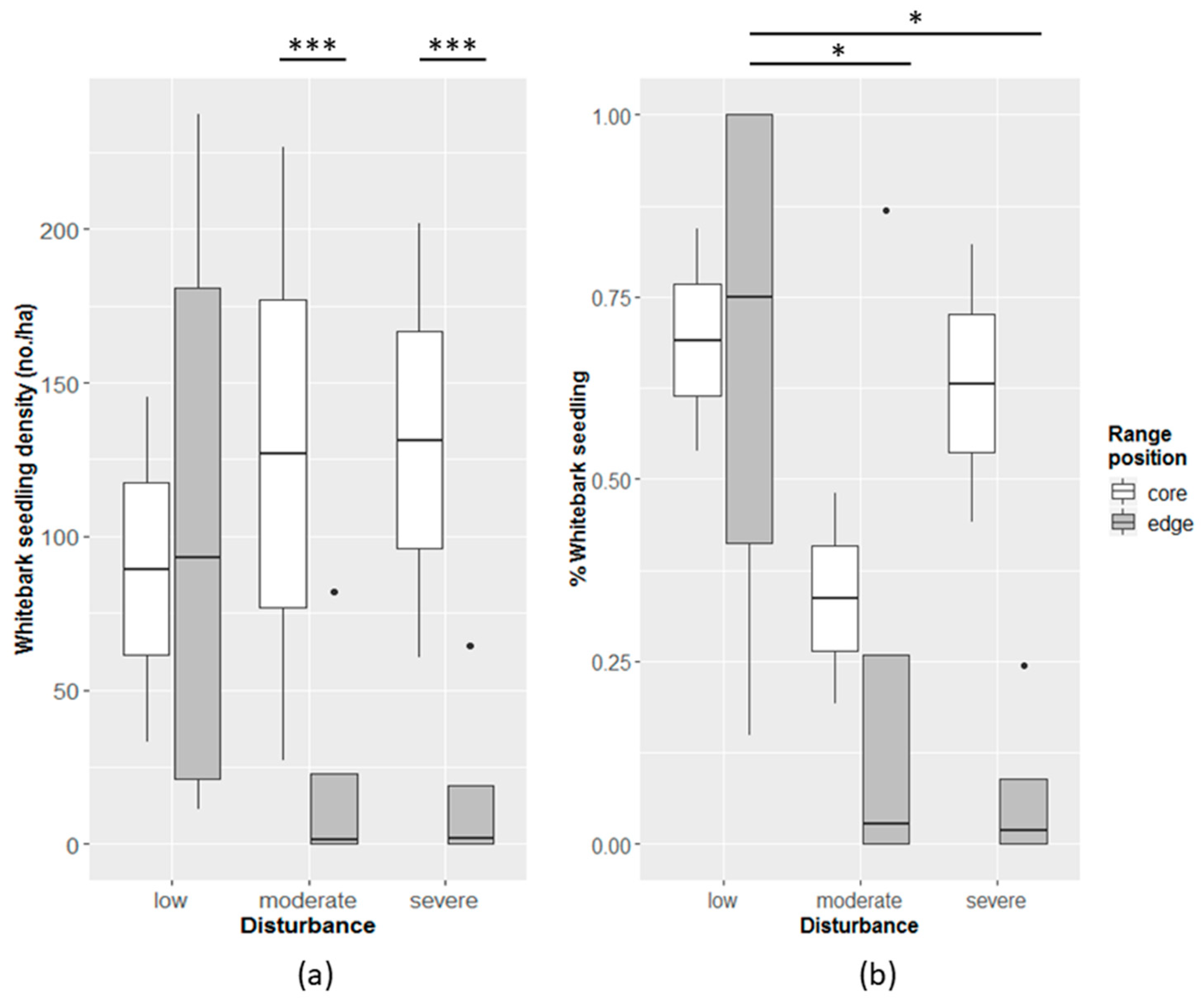
3.1. Plot-Based Model Development
3.2. Stem Density in Recreational Sites
3.3. Observational Case Study in Mountain Pine Beetle Attack at June Mt.
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rice, R.; Bales, C.; Painter, T.H.; Dozier, J. Snow water equivalent along elevation gradients in the Merced and Tuolumne River basins of the Sierra Nevada. Water Resour. Res. 2011, 47, W08515. [Google Scholar] [CrossRef]
- United States Fish and Wildlife Service. 12-month finding on a petition to list Pinus albicaulis as Endangered or Threatened with critical habitat. Fed. Regist. 2011, 76, 42631–42654. [Google Scholar]
- Slaton, M.R.; Meyer, M.; Gross, S.; Nesmith, J.; Dudney, J.; van Mantgem, P.; Butz, R. Subalpine sentinels: The status of whitebark pine (Pinus albicaulis) in California. Fremontia 2019, 47, in press. [Google Scholar]
- Schoettle, A.W.; Sniezko, R.A. Proactive intervention to sustain high-elevation pine ecosystems threatened by white pine blister rust. J. For. Res. 2007, 12, 327–336. [Google Scholar] [CrossRef]
- Schwandt, J.W.; Lockman, I.B.; Kliejunas, J.T.; Muir, J.A. Current health issues and management strategies for white pines in the western United States and Canada. For. Pathol. 2010, 40, 226–250. [Google Scholar] [CrossRef]
- Fiedler, C.E.; McKinney, S.T. Forest structure, health, and mortality in two Rocky Mountain whitebark pine ecosystems: Implications for restoration. Nat. Areas J. 2014, 34, 290–299. [Google Scholar] [CrossRef]
- Keane, R.E.; Tomback, D.F.; Aubry, C.A.; Bower, A.D.; Campbell, E.M.; Cripps, C.L.; Jenkins, M.B.; Mahalovich, M.F.; Manning, M.; McKinney, S.T.; et al. A Range-Wide Restoration Strategy for Whitebark Pine (Pinus albicaulis); RMRS-GTR-279; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2012.
- Perkins, J.L. Fire enhances whitebark pine seedling establishment, survival, and growth. Fire Ecol. 2015, 11, 84–99. [Google Scholar] [CrossRef]
- Arno, S.F.; Weaver, T. Whitebark pine community types and their patterns on the landscape. In Symposium on Whitebark Pine Ecosystems: Ecology and Management of a High-Mountain Resource; General Technical Report INT-270; Schmidt, W.C., McDonald, K.J., Eds.; USDA Forest Service, Intermountain Research Station: Ogden, UT, USA, 1990; pp. 97–105. [Google Scholar]
- Larson, E.R.; Kipfmueller, K.F. Patterns in whitebark pine regeneration and their relationships to biophysical site characteristics in southwest Montana, central Idaho, and Oregon, USA. Can. J. For. Res. 2010, 40, 476–487. [Google Scholar] [CrossRef]
- Tomback, D.F.; Arno, S.F.; Keane, R.E. The compelling case for management intervention. In Whitebark Pine Communities: Ecology and Restoration; Tomback, D., Arno, S.F., Keane, R.E., Eds.; Island Press: Washington, DC, USA, 2001; pp. 3–28. [Google Scholar]
- Perkins, D.L.; Roberts, D.W. Predictive models of whitebark pine mortality from mountain pine beetle. For. Ecol. Manag. 2003, 174, 495–510. [Google Scholar] [CrossRef]
- Meyer, M.D.; Bulaon, B.; MacKenzie, M.; Safford, H.D. Mortality, structure, and regeneration in whitebark pine stands impacted by mountain pine beetle in the southern Sierra Nevada. Can. J. For. Res. 2016, 46, 572–581. [Google Scholar] [CrossRef]
- Coutts, S.R.; Salguero-Gomez, R.; Csergo, A.M.; Buckley, Y.M. Extrapolating demography with climate, proximity and phylogeny: Approach with caution. Ecol. Lett. 2016, 19, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Purves, D.W. The demography of range boundaries versus range cores in eastern US tree species. Proc. Biol. Sci. 2009, 276, 1477–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadenasso, M.L.; Pickett, S.T.A. Linking forest edge structure to edge function: Mediation of herbivore damage. J. Ecol. 2000, 88, 31–44. [Google Scholar] [CrossRef]
- Millar, C.I.; Westfall, R.D.; Delany, D.L.; Bokach, M.J.; Flint, A.L.; Flint, L.E. Forest mortality in high-elevation whitebark pine (Pinus albicaulis) forests of eastern California, USA; influence of environmental context, bark beetles, climatic water deficit, and warming. Can. J. For. Res. 2012, 42, 749–765. [Google Scholar] [CrossRef]
- Amman, G.; McGregor, M.; Cahill, D.; Klein, W. Guidelines for Reducing Losses of Lodgepole Pine to the Mountain Pine Beetle in Unmanaged Stands in the Rocky Mountains; General Technical Report INT-36; USDA Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1977.
- Bentz, B.J.; Regniere, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negron, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. Bioscience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Gelderman, M.S.; Macdonald, S.E.; Gould, A.J. Regeneration niche of whitebark pine in the Canadian Rocky Mountains: The basis to restoring an endangered species. Arc. Antarct. Alp. Res. 2016, 48, 279–292. [Google Scholar] [CrossRef]
- Pansing, E.R.; Tomback, D.F.; Wunder, M.B.; French, J.P.; Wagner, A.C. Microsite and elevation zone effects on seed pilferage, germination, and seedling survival during early whitebark pine recruitment. Ecol. Evol. 2017, 7, 9027–9040. [Google Scholar] [CrossRef] [PubMed]
- Larson, E.R.; Kipfmueller, K.F. Ecological disaster or the limits of observation? Reconciling modern declines with the long-term dynamics of whitebark pine communities. Geogr. Comp. 2012, 6, 189–214. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing, 3.1 ed.; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Flint, L.E.; Flint, A.L. California Basin Characterization Model: A Dataset of Historical and Future Hydrologic Response to Climate Change; BCMv52; U.S. Geological Survey Data Release: Sacramento, CA, USA, 2014.
- Zeileis, A. Lmtest v-0.9-36. R Package. Available online: https://cran.r-project.org/web/packages/ lmtest/index.html (accessed on 23 May 2019).
- Gross, J.; Ligges, U. Nortest: Tests for Normality, R package version 1.0-3; Available online: https://cran.r-project.org/web/packages/ nortest/index.html (accessed on 23 May 2019).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Lenth, R. R Package, Estimated Marginal Means (EMMEANS). R package. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 23 May 2019).
- Gibson, K.; Skov, K.; Kegley, S.; Jorgensen, C.; Smith, S.; Witcosky, J. Mountain Pine Beetle Impacts in High-Elevation Five-Needle Pines: Current Trends and Challenges; R1-08-020; USDA Forest Service Forest Health and Protection: Missoula, MT, USA, 2008.
- Furniss, M.M.; Renkin, R. Forest entomology in Yellowstone National Park, 1923–1957: A time of Discovery and learning to let live. Am. Entomol. 2003, 49, 198–209. [Google Scholar] [CrossRef]
- Arno, S.F.; Hoff, R.J. Silvics of Whitebark Pine (Pinus albicualis); General Technical Report INT-253; US Forest Service, Intermountain Research Station: Ogden, UT, USA, 1989.
- Retzlaff, M.L.; Keane, R.E.; Affleck, D.L.; Hood, S.M. Growth response of whitebark pine (Pinus albicaulis Engelm) regeneration to thinning and prescribed burn treatments. Forests 2018, 9, 311. [Google Scholar] [CrossRef]
- Maloney, P.E. The multivariate underpinnings of recruitment for three Pinus species in montane forests of the Sierra Nevada, USA. Plant Ecol. 2014, 215, 261–274. [Google Scholar] [CrossRef]
- Leirfallom, S.B.; Keane, R.E.; Tomback, D.F.; Dobrowski, S.Z. The effects of seed source health on whitebark pine (Pinus albicaulis) regeneration density after wildfire. Can. J. For. Res. 2015, 45, 1597–1606. [Google Scholar] [CrossRef]
- Amberson, J.T.; Keville, M.P.; Nelson, C.R. Effects of disturbance on tree community dynamics in whitebark pine (Pinus albicaulis Engelm.) Ecosystems. Forests 2018, 9, 566. [Google Scholar] [CrossRef]
- Tomback, D.F. Dispersal of whitebark pine seeds by Clark nutcracker—A mutualism hypothesis. J. Anim. Ecol. 1982, 51, 451–467. [Google Scholar] [CrossRef]
- Maher, E.L.; Germino, M.J.; Hasselquist, N.J. Interactive effects of tree and herb cover on survivorship, physiology, and microclimate of conifer seedlings at the alpine tree-line ecotone. Can. J. For. Res. 2005, 35, 567–574. [Google Scholar] [CrossRef]
- Maher, E.L.; Germino, M.J. Microsite differentiation among conifer species during seedling establishment at alpine treeline. Ecoscience 2006, 13, 334–341. [Google Scholar] [CrossRef]
- Lorenz, T.J.; Aubry, C.; Shoal, R. A Review of the Literature on Seed Fate in Whitebark Pine and the Life History Traits of Clark’s Nutcracker and Pine Squirrels; General Technical Report PNW-GTR-742; US Forest Service Pacific Northwest Research Station: Portland, OR, USA, 2008; p. 62.
- Wiens, D.; Slaton, M.R. The mechanism of background extinction. Biol. J. Linn. Soc. 2012, 105, 255–268. [Google Scholar] [CrossRef]
- Keane, R.E.; Parsons, R.A. A Management Guide to Ecosystem Restoration Treatments: Whitebark Pine Forests of the Northern Rocky Mountains; General Technical Report RMRS-GTR-232; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2010.
- Clason, A.J.; Macdonald, S.E.; Haeussler, S. Forest response to cumulative disturbance and stress: Two decades of change in whitebark pine ecosystems of west-central British Columbia. Ecoscience 2014, 21, 174–185. [Google Scholar] [CrossRef]
- Campbell, E.M.; Antos, J.A. Postfire succession in Pinus albicaulis—Abies lasiocarpa forests of southern British Columbia. Can. J. Bot. 2003, 81, 383–397. [Google Scholar] [CrossRef]
- Kremer, A.; Potts, B.M.; Delzon, S. Genetic divergence in forest trees: Understanding the consequences of climate change. Funct. Ecol. 2014, 28, 22–36. [Google Scholar] [CrossRef]
- Rehm, E.M.; Olivas, P.; Stroud, J.; Feeley, K.J. Losing your edge: Climate change and the conservation value of range-edge populations. Ecol. Evol. 2015, 5, 4315–4326. [Google Scholar] [CrossRef]
- Maloney, P.E.; Vogler, R.; Jensen, C.E.; Mix, A.D. Ecology of whitebark pine populations in relation to white pine blister rust infection in subalpine forests of the Lake Tahoe Basin, USA: Implications for restoration. For. Ecol. Manag. 2012, 280, 166–175. [Google Scholar] [CrossRef]
- Slaton, M.R. The roles of disturbance, topography and climate in determining the leading and rear edges of population range limits. J. Biogeogr. 2015, 42, 255–266. [Google Scholar] [CrossRef]
- Sanchez-Salguero, R.; Camarero, J.J.; Gutierrez, E.; Rouco, F.G.; Gazol, A.; Sanguesa-Barreda, G.; Andreu-Hayles, L.; Linares, J.C.; Seftigen, K. Assessing forest vulnerability to climate warming using a process-based model of tree growth: Bad prospects for rear-edges. Glob. Chang. Biol. 2017, 23, 2705–2719. [Google Scholar] [CrossRef] [PubMed]
- Thom, D.; Rammer, W.; Seidl, R. Disturbances catalyze the adaptation of forest ecosystems to changing climate conditions. Glob. Chang. Biol. 2017, 23, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Thom, D.; Rammer, W.; Dirnbock, T.; Muller, J.; Kobler, J.; Katzensteiner, K.; Helm, N.; Seidl, R. The impacts of climate change and disturbance on spatio-temporal trajectories of biodiversity in a temperate forest landscape. J. Appl. Ecol. 2017, 54, 28–38. [Google Scholar] [CrossRef]
- Warwell, M.V.; Rehfeldt, G.E.; Crookston, N.L. Modeling contemporary climate profiles of whitebark pine (Pinus albicaulis) and predicting responses to global warming. In Proceedings of the Conference Whitebark Pine: A Pacific Coast Perspective, Ashland, OR, USA, 27–31 August 2006; USDA Forest Service, Pacific Northwest Region: Ashland, OR, USA, 2007; Volume R6-NR-FHP-2007-01, pp. 139–142. [Google Scholar]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Whitebark Seedling Density | Whitebark Seedling Proportion (%) | |
|---|---|---|
| Disturbance severity | + ** | + * |
| Whitebark main canopy cover | + *** | + *** |
| Elevation | + * | + *** |
| Elevation^2 | * | − |
| Latitude | + * | ns |
| Latitude^2 | * | − |
| Slope | − * | ns |
| Aspect | ns | ns |
| R-squared | 0.32 | 0.47 |
| Breusch-Pagan Test | 0.009 | 0.08 |
| Anderson Darling Test | 0.20 | 0.71 |
| Factor | Coefficient | SE | p(Wald) | Variance of Random Effects | AIC | p(χ2) Comparison to Model C |
|---|---|---|---|---|---|---|
| Model A: seedling.density ~ range + offset(log(area)) + (1 | site) | ||||||
| 1.116 | 239.2 | 0.001 | ||||
| Intercept | 3.94477 | 0.5435 | <0.00001 | |||
| range.edge | −0.62307 | 0.09623 | <0.00001 | |||
| Model B: seedling.density ~ disturb + offset(log(area)) + (1 | site) | ||||||
| 1.592 | 266.4 | <0.00001 | ||||
| Intercept | 3.57984 | 0.64374 | <0.00001 | |||
| disturb.mod | −0.26221 | 0.09626 | 0.00645 | |||
| disturb.severe | 0.12024 | 0.0976 | 0.21979 | |||
| Model C: seedling.density ~ disturb + range + offset(log(area)) + (1 | site) | ||||||
| 1.156 | 229.8 | na | ||||
| Intercept | 3.98644 | 0.55606 | <0.00001 | |||
| range.edge | −0.59628 | 0.09639 | <0.00001 | |||
| disturb.mod | −0.25254 | 0.09774 | 0.0088 | |||
| disturb.severe | 0.07745 | 0.09654 | 0.4281 | |||
| Model D: seedling.density ~ disturb + range + (disturb × range) + offset(log(area)) + (1 | site) | ||||||
| 0.9956 | 158.2 | <0.00001 | ||||
| Intercept | 3.6453 | 0.5242 | <0.00001 | |||
| range.edge | 0.2514 | 0.1451 | 0.0831 | |||
| disturb.mod | 0.2964 | 0.1367 | 0.0301 | |||
| disturb.severe | 0.7172 | 0.1321 | <0.00001 | |||
| disturb.mod:range.edge | −1.1884 | 0.2038 | <0.00001 | |||
| disturb.severe:range.edge | −1.9335 | 0.2545 | <0.00001 | |||
| Factor | Coefficient | SE | p (Wald) | Variance of Random Effects | AIC | p(χ2) Comparison to Model C |
|---|---|---|---|---|---|---|
| Model A: % seedlings ~ range + (1 | site) | ||||||
| 0.03241 | 23.9 | 0.0042 | ||||
| Intercept | 0.479 | 0.1724 | 0.0277 | |||
| range.edge | −0.1611 | 0.1741 | 0.3689 | |||
| Model B: % seedlings ~ disturb + (1 | site) | ||||||
| 0.05837 | 18.9 | 0.3032 | ||||
| Intercept | 0.6339 | 0.1537 | 0.00845 | |||
| disturb.mod | −0.4497 | 0.1322 | 0.00522 | |||
| disturb.severe | −0.4097 | 0.1322 | 0.00916 | |||
| Model C: % seedlings ~ disturb + range + (1 | site) | ||||||
| 0.04517 | 22.4 | na | ||||
| Intercept | 0.7265 | 0.1736 | 0.00268 | |||
| range.edge | −0.1221 | 0.1315 | 0.3709 | |||
| disturb.mod | −0.4497 | 0.1366 | 0.0072 | |||
| disturb.severe | −0.4097 | 0.1366 | 0.01213 | |||
| Model D: % seedlings ~ disturb + range + (disturb × range) + (1 | site) | ||||||
| 0.0507 | 22 | 0.01197 | ||||
| Intercept | 0.53301 | 0.18226 | 0.016 | |||
| range.edge | 0.15943 | 0.17205 | 0.3775 | |||
| disturb.mod | −0.34372 | −0.34372 | 0.1044 | |||
| disturb.severe | 0.01277 | 0.01277 | 0.948 | |||
| disturb.mod:range.edge | −0.15894 | 0.23308 | 0.5125 | |||
| disturb.severe:range.edge | −0.63366 | 0.23308 | 0.0237 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slaton, M.R.; MacKenzie, M.; Kohler, T.; Ramirez, C.M. Whitebark Pine Recruitment in Sierra Nevada Driven by Range Position and Disturbance History. Forests 2019, 10, 455. https://doi.org/10.3390/f10050455
Slaton MR, MacKenzie M, Kohler T, Ramirez CM. Whitebark Pine Recruitment in Sierra Nevada Driven by Range Position and Disturbance History. Forests. 2019; 10(5):455. https://doi.org/10.3390/f10050455
Chicago/Turabian StyleSlaton, Michèle R., Martin MacKenzie, Tanya Kohler, and Carlos M. Ramirez. 2019. "Whitebark Pine Recruitment in Sierra Nevada Driven by Range Position and Disturbance History" Forests 10, no. 5: 455. https://doi.org/10.3390/f10050455