What Makes the Wood? Exploring the Molecular Mechanisms of Xylem Acclimation in Hardwoods to an Ever-Changing Environment

 ,
,
Abstract
:
1. Introduction: The Xylem Keeps the Stream of Life Flowing
2. Anatomy and Molecular Biology of Wood Formation
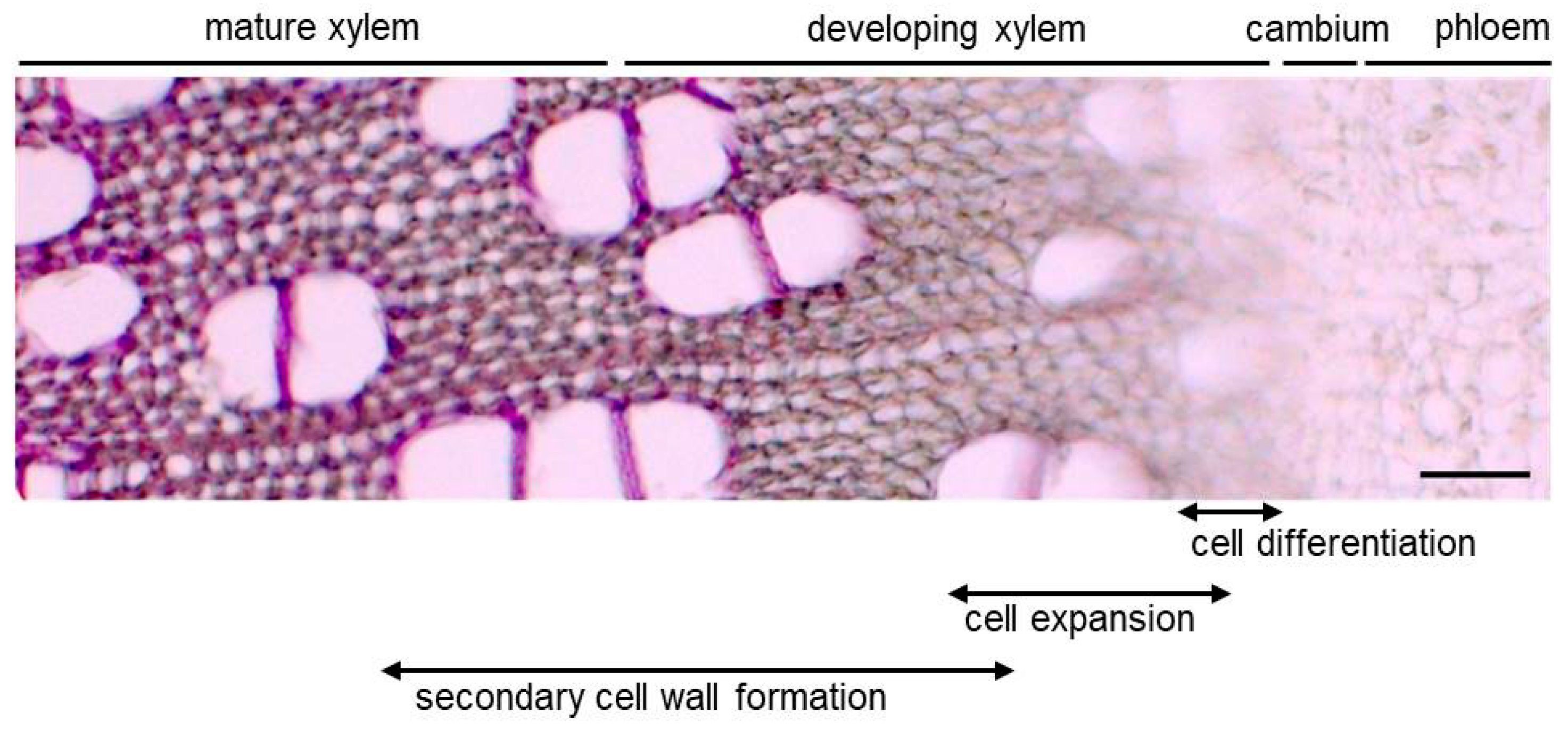
2.1. What Makes the Stem?
2.2. Molecular Mechanism of Wood Formation
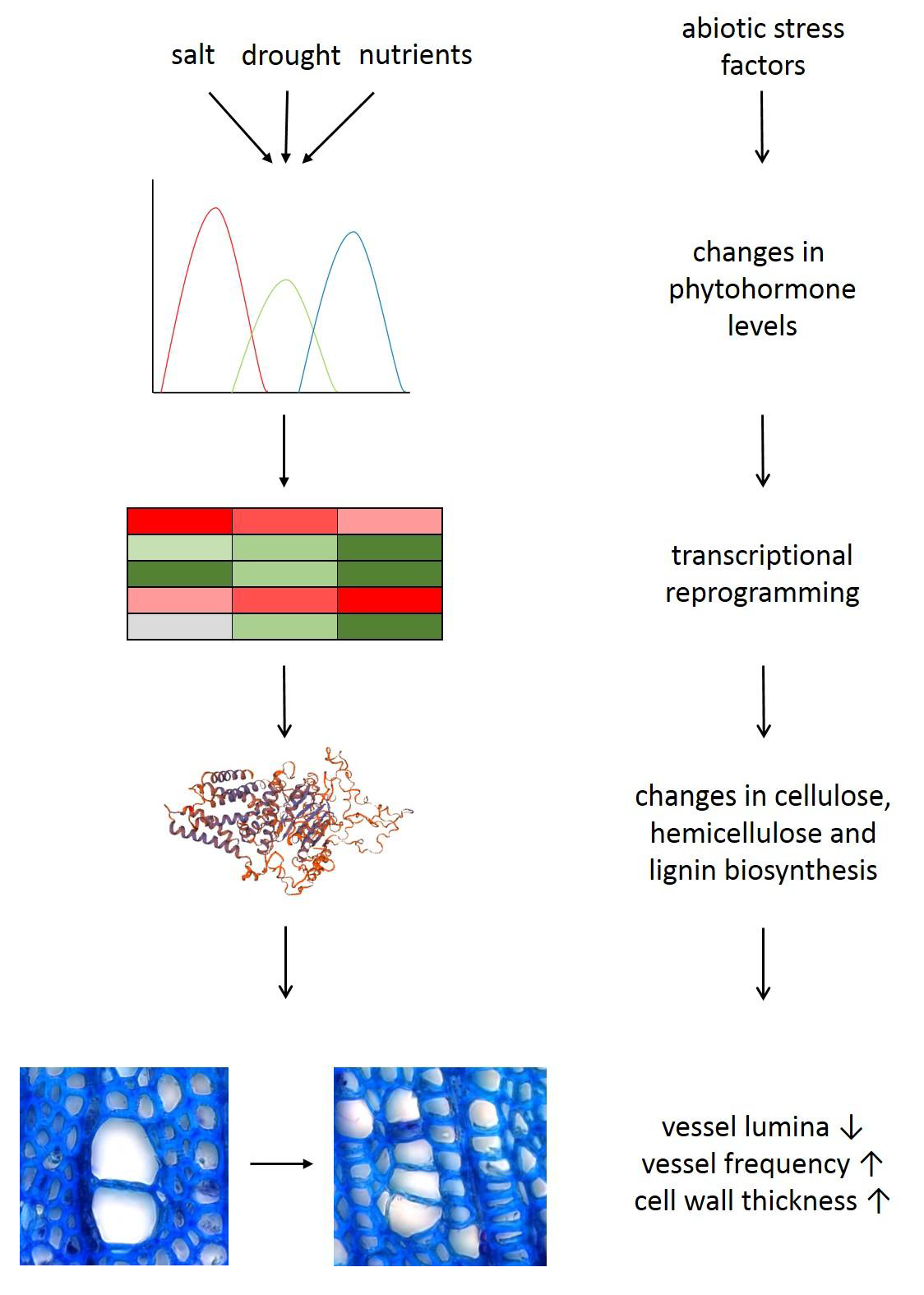
3. Abiotic Stresses Affecting Wood Formation
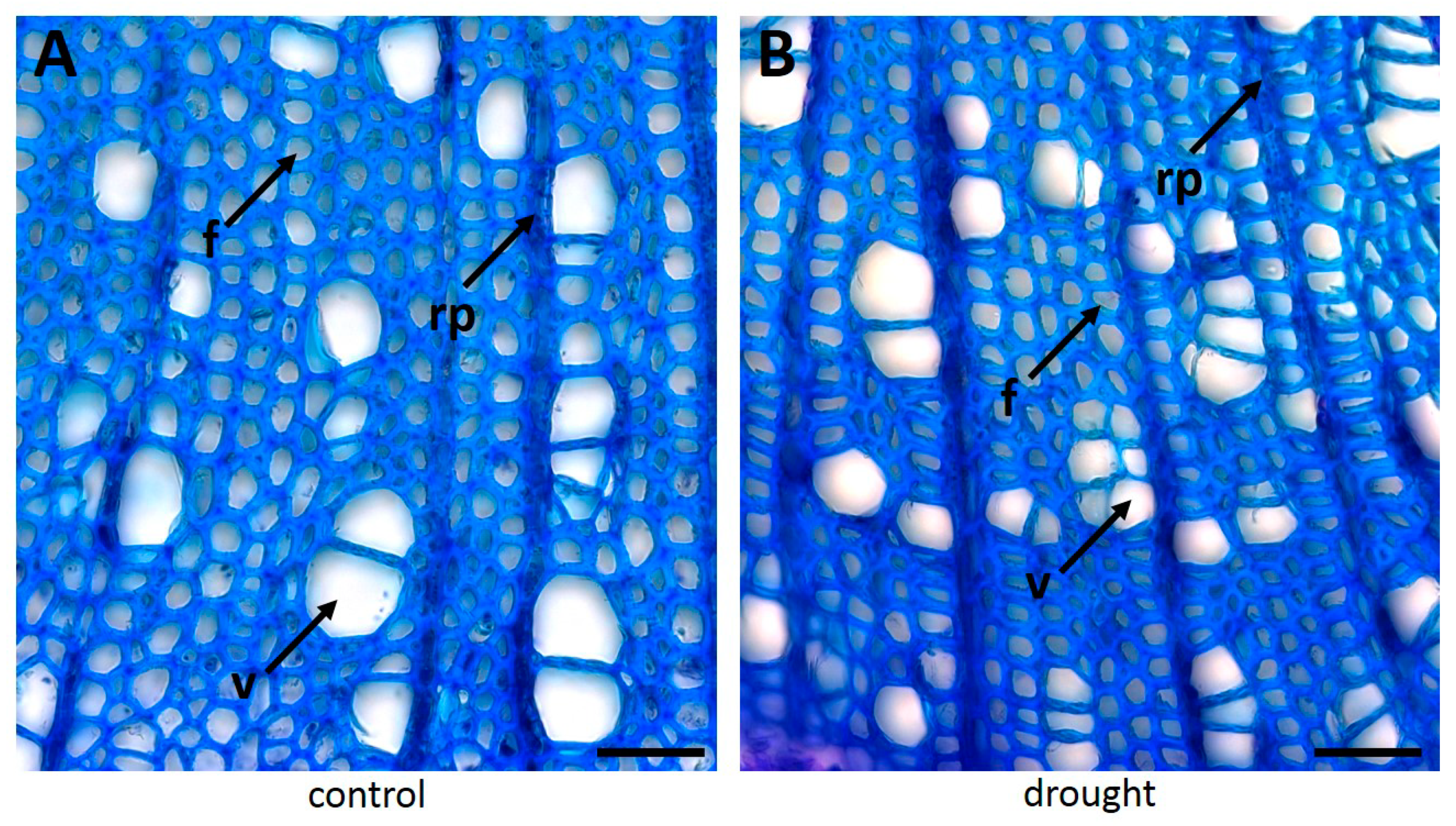
3.1. Drought Severely Changes Xylem Anatomy
3.2. Drought Leads to Major Transcriptional Remodelling
3.3. Phytohormones Mediate Xylem Changes under Drought
3.4. Salt Severely Affects Wood Formation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boer, A.H.D.; Volkov, V. Logistics of water and salt transport through the plant: Structure and functioning of the xylem. Plant Cell Environ. 2003, 26, 87–101. [Google Scholar] [CrossRef]
- Myburg, A.A.; Lev-Yadun, S.; Sederoff, R.R. Xylem Structure and Function. In Encycolpedia of Life Science; American Cancer Society: London, UK, 2013; ISBN 978-0-470-01590-2. [Google Scholar]
- Tyree, M.T.; Sperry, J.S. Vulnerability of Xylem to Cavitation and Embolism. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1989, 40, 19–36. [Google Scholar] [CrossRef]
- Sperry, J.S.; Hacke, U.G.; Pittermann, J. Size and function in conifer tracheids and angiosperm vessels. Am. J. Bot. 2006, 93, 1490–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alder, N.N.; Sperry, J.S.; Pockman, W.T. Root and stem xylem embolism, stomatal conductance, and leaf turgor in Acer grandidentatum populations along a soil moisture gradient. Oecologia 1996, 105, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Cobb, A.R.; Jansen, S. Structure and function of bordered pits: New discoveries and impacts on whole-plant hydraulic function. New Phytol. 2008, 177, 608–626. [Google Scholar] [CrossRef]
- Zwieniecki, M.A.; Holbrook, N.M. Confronting Maxwell’s demon: Biophysics of xylem embolism repair. Trends Plant Sci. 2009, 14, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Awad, H.; Barigah, T.; Badel, E.; Cochard, H.; Herbette, S. Poplar vulnerability to xylem cavitation acclimates to drier soil conditions. Physiologia Plantarum 2010, 139, 208–288. [Google Scholar] [CrossRef]
- Rita, A.; Cherubini, P.; Leonardi, S.; Todaro, L.; Borghetti, M. Functional adjustments of xylem anatomy to climatic variability: Insights from long-term Ilex aquifolium tree-ring series. Tree Physiol. 2015, 35, 817–828. [Google Scholar] [CrossRef]
- Wildhagen, H.; Paul, S.; Allwright, M.; Smith, H.K.; Malinowska, M.; Schnabel, S.K.; Paulo, M.J.; Cattonaro, F.; Vendramin, V.; Scalabrin, S.; et al. Genes and gene clusters related to genotype and drought-induced variation in saccharification potential, lignin content and wood anatomical traits in Populus nigra. Tree Physiol 2018, 38, 320–339. [Google Scholar] [CrossRef] [PubMed]
- Bogeat-Triboulot, M.-B.; Brosché, M.; Renaut, J.; Jouve, L.; Thiec, D.L.; Fayyaz, P.; Vinocur, B.; Witters, E.; Laukens, K.; Teichmann, T.; et al. Gradual soil water depletion results in reversible changes of gene expression, protein profiles, ecophysiology, and growth performance in Populus euphratica, a poplar growing in arid regions. Plant Physiol. 2007, 143, 876–892. [Google Scholar] [CrossRef] [PubMed]
- Aichinger, E.; Kornet, N.; Friedrich, T.; Laux, T. Plant Stem Cell Niches. Annu. Rev. Plant Biol. 2012, 63, 615–636. [Google Scholar] [CrossRef] [PubMed]
- Plomion, C.; Leprovost, G.; Stokes, A. Wood Formation in Trees. Plant Physiol. 2001, 127, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Furuta, K.M.; Hellmann, E.; Helariutta, Y. Molecular control of cell specification and cell differentiation during procambial development. Annu. Rev. Plant Biol. 2014, 65, 607–638. [Google Scholar] [CrossRef] [PubMed]
- Słupianek, A.; Kasprowicz-Maluśki, A.; Myśkow, E.; Turzańska, M.; Sokołowska, K. Endocytosis acts as transport pathway in wood. New Phytol. 2018. [Google Scholar] [CrossRef]
- Růžička, K.; Ursache, R.; Hejátko, J.; Helariutta, Y. Xylem development – from the cradle to the grave. New Phytol. 2015, 207, 519–535. [Google Scholar] [CrossRef]
- Déjardin, A.; Laurans, F.; Arnaud, D.; Breton, C.; Pilate, G.; Leplé, J.-C. Wood formation in Angiosperms. Comptes Rendus Biologies 2010, 333, 325–334. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, H.E.; Döring, A.; Persson, S. The cell biology of cellulose synthesis. Annu. Rev. Plant Biol. 2014, 65, 69–94. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Gille, S.; Liu, L.; Mansoori, N.; de Souza, A.; Schultink, A.; Xiong, G. Hemicellulose biosynthesis. Planta 2013, 238, 627–642. [Google Scholar] [CrossRef]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- Voxeur, A.; Wang, Y.; Sibout, R. Lignification: Different mechanisms for a versatile polymer. Curr. Opin. Plant Biol. 2015, 23, 83–90. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and Biological Functions in Plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, E.; Bokelman, G.H.; Lewis, N.G. Phenylpropanoid Metabolism in Cell Walls. In Plant Cell Wall Polymers; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1989; Volume 399, pp. 68–88. ISBN 978-0-8412-1658-7. [Google Scholar]
- Zhao, Q. Lignification: Flexibility, biosynthesis and regulation. Trends Plant Sci. 2016, 21, 713–721. [Google Scholar] [CrossRef]
- Barros, J.; Serk, H.; Granlund, I.; Pesquet, E. The cell biology of lignification in higher plants. Ann. Bot. 2015, 115, 1053–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuetz, M.; Smith, R.; Ellis, B. Xylem tissue specification, patterning, and differentiation mechanisms. J. Exp. Bot. 2013, 64, 11–31. [Google Scholar] [CrossRef]
- Ye, Z.-H.; Zhong, R. Molecular control of wood formation in trees. J. Exp. Bot. 2015, 66, 4119–4131. [Google Scholar] [CrossRef] [Green Version]
- Kubo, M.; Udagawa, M.; Nishikubo, N.; Horiguchi, G.; Yamaguchi, M.; Ito, J.; Mimura, T.; Fukuda, H.; Demura, T. Transcription switches for protoxylem and metaxylem vessel formation. Genes Dev. 2005, 19, 1855–1860. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Mitsuda, N.; Ohtani, M.; Ohme-Takagi, M.; Kato, K.; Demura, T. Vascular-related NAC-domain 7 directly regulates a broad range of genes for xylem vessel differentiation. BMC Proc. 2011, 5, O37. [Google Scholar] [CrossRef]
- Zhong, R.; Lee, C.; Ye, Z.-H. Functional characterization of poplar wood-associated NAC domain transcription factors. Plant Physiol. 2010, 152, 1044–1055. [Google Scholar] [CrossRef] [PubMed]
- Mariconti, L.; Pellegrini, B.; Cantoni, R.; Stevens, R.; Bergounioux, C.; Cella, R.; Albani, D. The E2F family of transcription factors from Arabidopsis thaliana. J. Biol. Chem. 2002, 277, 9911–9919. [Google Scholar] [CrossRef]
- Du, J.; Miura, E.; Robischon, M.; Martinez, C.; Groover, A. The Populus class III HD ZIP transcription factor POPCORONA affects cell differentiation during secondary growth of woody stems. PLoS ONE 2011, 6, e17458. [Google Scholar] [CrossRef]
- McCarthy, R.L.; Zhong, R.; Fowler, S.; Lyskowski, D.; Piyasena, H.; Carleton, K.; Spicer, C.; Ye, Z.-H. The poplar MYB transcription factors, PtrMYB3 and PtrMYB20, are involved in the regulation of secondary wall biosynthesis. Plant Cell Physiol. 2010, 51, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; McCarthy, R.L.; Haghighat, M.; Ye, Z.-H. The poplar MYB master switches bind to the SMRE site and activate the secondary wall biosynthetic program during wood formation. PLoS ONE 2013, 8, e69219. [Google Scholar] [CrossRef] [PubMed]
- Pear, J.R.; Kawagoe, Y.; Schreckengost, W.E.; Delmer, D.P.; Stalker, D.M. Higher plants contain homologs of the bacterial celA genes encoding the catalytic subunit of cellulose synthase. Proc. Natl. Acad. Sci. USA 1996, 93, 12637–12642. [Google Scholar] [CrossRef] [PubMed]
- Djerbi, S.; Lindskog, M.; Arvestad, L.; Sterky, F.; Teeri, T.T. The genome sequence of black cottonwood (Populus trichocarpa) reveals 18 conserved cellulose synthase (CesA) genes. Planta 2005, 221, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Li, L.; Sun, Y.-H.; Chiang, V.L. The Cellulose Synthase Gene Superfamily and Biochemical Functions of Xylem-Specific Cellulose Synthase-Like Genes in Populus trichocarpa. Plant Physiol. 2006, 142, 1233–1245. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Schneider, R.; Barkwill, S.; Gonzales-Vigil, E.; Hill, J.L., Jr.; Samuels, A.L.; Persson, S.; Mansfield, S.D. Cellulose synthase complexes display distinct dynamic behaviors during xylem transdifferentiation. Proc. Natl. Acad. Sci. USA 2018, 115, E6366–E6374. [Google Scholar] [CrossRef] [PubMed]
- Polle, A.; Janz, D.; Teichmann, T.; Lipka, V. Poplar genetic engineering: Promoting desirable wood characteristics and pest resistance. Appl. Microbiol. Biotechnol. 2013, 97, 5669–5679. [Google Scholar] [CrossRef]
- Liu, C.-J.; Miao, Y.-C.; Zhang, K.-W. Sequestration and transport of lignin monomeric precursors. Molecules 2011, 16, 710–727. [Google Scholar] [CrossRef]
- Alejandro, S.; Lee, Y.; Tohge, T.; Sudre, D.; Osorio, S.; Park, J.; Bovet, L.; Lee, Y.; Geldner, N.; Fernie, A.R.; Martinoia, E. AtABCG29 Is a Monolignol Transporter Involved in Lignin Biosynthesis. Curr. Biol. 2012, 22, 1207–1212. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.-C.; Liu, C.-J. ATP-binding cassette-like transporters are involved in the transport of lignin precursors across plasma and vacuolar membranes. Proc. Natl. Acad. Sci. USA 2010, 107, 22728–22733. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, A.P.S.; Randhawa, G.S.; Dhugga, K.S. Plant cell wall matrix polysaccharide biosynthesis. Mol. Plant 2009, 2, 840–850. [Google Scholar] [CrossRef]
- Savi, T.; Bertuzzi, S.; Branca, S.; Tretiach, M.; Nardini, A. Drought-induced xylem cavitation and hydraulic deterioration: Risk factors for urban trees under climate change? New Phytol. 2015, 205, 1106–1116. [Google Scholar] [CrossRef]
- Nardini, A.; Savi, T.; Trifilò, P.; Gullo, M.A.L. Drought stress and the recovery from xylem embolism in woody plants. In Progress in Botany; Progress in Botany; Springer: Cham, Switzerland, 2017; Volume 79, pp. 197–231. ISBN 978-3-319-71412-7. [Google Scholar]
- Fischer, U.; Polle, A. Populus responses to abiotic stress. In Genetics and Genomics of Populus; Plant Genetics and Genomics: Crops and Models; Springer: New York, NY, USA, 2010; pp. 225–246. ISBN 978-1-4419-1540-5. [Google Scholar]
- Polle, A.; Chen, S.L.; Eckert, C.; Harfouche, A. Engineering drought resistance in forest trees. Front. Plant Sci. 2019, 9. [Google Scholar] [CrossRef]
- Arend, M.; Fromm, J. Seasonal change in the drought response of wood cell development in poplar. Tree Physiol. 2007, 27, 985. [Google Scholar] [CrossRef] [PubMed]
- Beniwal, R.S.; Langenfeld-Heyser, R.; Polle, A. Ectomycorrhiza and hydrogel protect hybrid poplar from water deficit and unravel plastic responses of xylem anatomy. Environ. Exp. Bot. 2010, 69, 189–197. [Google Scholar] [CrossRef]
- Schreiber, S.G.; Hacke, U.G.; Chamberland, S.; Lowe, C.W.; Kamelchuk, D.; Bräutigam, K.; Campbell, M.M.; Thomas, B.R. Leaf size serves as a proxy for xylem vulnerability to cavitation in plantation trees. Plant Cell Environ. 2016, 39, 272–281. [Google Scholar] [CrossRef]
- Fonti, P.; Heller, O.; Cherubini, P.; Rigling, A.; Arend, M. Wood anatomical responses of oak saplings exposed to air warming and soil drought. Plant Biol. 2013, 15, 210–219. [Google Scholar] [CrossRef]
- Colangelo, M.; Camarero, J.J.; Borghetti, M.; Gazol, A.; Gentilesca, T.; Ripullone, F. Size matters a lot: Drought-affected italian oaks are smaller and show lower growth prior to tree death. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Fichot, R.; Barigah, T.S.; Chamaillard, S.; Le Thiec, D.; Laurans, F.; Cochard, H.; Brignolas, F. Common trade-offs between xylem resistance to cavitation and other physiological traits do not hold among unrelated Populus deltoides x Populus nigra hybrids: Xylem resistance to cavitation and water relations in poplar. Plant Cell Environ. 2010, 33, 1553–1568. [Google Scholar] [CrossRef] [PubMed]
- Rathgeber, C.B.K.; Cuny, H.E.; Fonti, P. Biological Basis of Tree-Ring Formation: A Crash Course. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Langer, K.; Ache, P.; Geiger, D.; Stinzing, A.; Arend, M.; Wind, C.; Regan, S.; Fromm, J.; Hedrich, R. Poplar potassium transporters capable of controlling K+ homeostasis and K+-dependent xylogenesis. Plant J. 2002, 32, 997–1009. [Google Scholar] [CrossRef]
- Wind, C.; Arend, M.; Fromm, J. Potassium-Dependent Cambial Growth in Poplar. Plant Biol. 2004, 7, 30–37. [Google Scholar] [CrossRef]
- Larisch, C.; Dittrich, M.; Wildhagen, H.; Lautner, S.; Fromm, J.; Polle, A.; Hedrich, R.; Rennenberg, H.; Müller, T.; Ache, P. Poplar wood rays are involved in seasonal remodeling of tree physiology. Plant Physiol. 2012, 160, 1515–1529. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, L.; Polle, A. Poplar nutrition under drought as affected by ectomycorrhizal colonization. Environ. Exp. Bot. 2014, 108, 89–98. [Google Scholar] [CrossRef]
- Giovannelli, A.; Emiliani, G.; Traversi, M.L.; Deslauriers, A.; Rossi, S. Sampling cambial region and mature xylem for non structural carbohydrates and starch analyses. Dendrochronologia 2011, 29, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Xue, L.-J.; Frost, C.J.; Tsai, C.-J.; Harding, S.A. Drought response transcriptomes are altered in poplar with reduced tonoplast sucrose transporter expression. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Dong, Y.; Liang, D.; Zhang, Z.; Ye, C.-Y.; Shuai, P.; Han, X.; Zhao, Y.; Yin, W.; Xia, X. Analysis of the drought stress-responsive transcriptome of black cottonwood (Populus trichocarpa) using deep RNA sequencing. Plant Mol. Biol. Rep. 2015, 33, 424–438. [Google Scholar] [CrossRef]
- Deslauriers, A.; Giovannelli, A.; Rossi, S.; Castro, G.; Fragnelli, G.; Traversi, L. Intra-annual cambial activity and carbon availability in stem of poplar. Tree Physiol. 2009, 29, 1223–1235. [Google Scholar] [CrossRef] [Green Version]
- Paux, E.; Tamasloukht, M.; Ladouce, N.; Sivadon, P.; Grima-Pettenati, J. Identification of genes preferentially expressed during wood formation in Eucalyptus. Plant Mol. Biol. 2004, 55, 263–280. [Google Scholar] [CrossRef]
- Druart, N.; Johansson, A.; Baba, K.; Schrader, J.; Sjödin, A.; Bhalerao, R.R.; Resman, L.; Trygg, J.; Moritz, T.; Bhalerao, R.P. Environmental and hormonal regulation of the activity-dormancy cycle in the cambial meristem involves stage-specific modulation of transcriptional and metabolic networks. Plant J. 2007, 50, 557–573. [Google Scholar] [CrossRef] [Green Version]
- Doblin, M.S.; Kurek, I.; Jacob-Wilk, D.; Delmer, D.P. Cellulose biosynthesis in plants: From genes to rosettes. Plant Cell Physiol. 2002, 43, 1407–1420. [Google Scholar] [CrossRef]
- Peng, L.; Kawagoe, Y.; Hogan, P.; Delmer, D. Sitosterol-β-glucoside as primer for cellulose synthesis in plants. Science 2002, 295, 147–150. [Google Scholar] [CrossRef]
- Leung, A.K.L.; Sharp, P.A. MicroRNA functions in stress responses. Mol. Cell 2010, 40, 205–215. [Google Scholar] [CrossRef]
- Sunkar, R.; Li, Y.-F.; Jagadeeswaran, G. Functions of microRNAs in plant stress responses. Trends Plant Sci. 2012, 17, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Shuai, P.; Liang, D.; Zhang, Z.; Yin, W.; Xia, X. Identification of drought-responsive and novel Populus trichocarpa microRNAs by high-throughput sequencing and their targets using degradome analysis. BMC Genomics 2013, 14, 233. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; Leijon, F.; Bulone, V. Radiometric and spectrophotometric in vitro assays of glycosyltransferases involved in plant cell wall carbohydrate biosynthesis. Nat. Protoc. 2012, 7, 1634–1650. [Google Scholar] [CrossRef]
- Secchi, F.; Pagliarani, C.; Zwieniecki, M.A. The functional role of xylem parenchyma cells and aquaporins during recovery from severe water stress. Plant Cell Environ. 2017, 40, 858–871. [Google Scholar] [CrossRef] [PubMed]
- Secchi, F.; Zwieniecki, M.A. Sensing embolism in xylem vessels: The role of sucrose as a trigger for refilling. Plant Cell Environ. 2011, 34, 514–524. [Google Scholar] [CrossRef] [PubMed]
- Secchi, F.; Maciver, B.; Zeidel, M.L.; Zwieniecki, M.A. Functional analysis of putative genes encoding the PIP2 water channel subfamily in Populus trichocarpa. Tree Physiol. 2009, 29, 1467–1477. [Google Scholar] [CrossRef]
- Secchi, F.; Zwieniecki, M.A. Patterns of PIP gene expression in Populus trichocarpa during recovery from xylem embolism suggest a major role for the PIP1 aquaporin subfamily as moderators of refilling process. Plant Cell Environ. 2010, 33, 1285–1297. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Rodriguez, A.M.; Hacke, U.G. Cellular localization of aquaporin mRNA in hybrid poplar stems. Am. J. Bot. 2012, 99, 1249–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakr, S.; Alves, G.; Morillon, R.; Maurel, K.; Decourteix, M.; Guilliot, A.; Fleurat-Lessard, P.; Julien, J.-L.; Chrispeels, M.J. Plasma membrane aquaporins are involved in winter embolism recovery in walnut tree. Plant Physiol. 2003, 133, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Nardini, A.; Lo Gullo, M.A.; Salleo, S. Refilling embolized xylem conduits: Is it a matter of phloem unloading? Plant Sci. 2011, 180, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Fernie, A.R. Remote Control of Transpiration via ABA. Trends Plant Sci. 2018, 23, 755–758. [Google Scholar] [CrossRef] [PubMed]
- Lachaud, S. Participation of auxin and abscisic acid in the regulation of seasonal variations in cambial activity and xylogenesis. Trees-Struct. Funct. 1989, 3, 125–137. [Google Scholar] [CrossRef]
- Luisi, A.; Giovannelli, A.; Traversi, M.L.; Anichini, M.; Sorce, C. Hormonal responses to water deficit in cambial tissues of Populus alba L. J. Plant Grow. Regul. 2014, 33, 489–498. [Google Scholar] [CrossRef]
- Tylewicz, S.; Petterle, A.; Marttila, S.; Miskolczi, P.; Azeez, A.; Singh, R.K.; Immanen, J.; Mähler, N.; Hvidsten, T.R.; Eklund, D.M.; et al. Photoperiodic control of seasonal growth is mediated by ABA acting on cell-cell communication. Science 2018, 360, 212–215. [Google Scholar] [CrossRef]
- Sorce, C.; Giovannelli, A.; Sebastiani, L.; Anfodillo, T. Hormonal signals involved in the regulation of cambial activity, xylogenesis and vessel patterning in trees. Plant Cell Rep. 2013, 32, 885–898. [Google Scholar] [CrossRef] [Green Version]
- Popko, J.; Haensch, R.; Mendel, R.R.; Polle, A.; Teichmann, T. The role of abscisic acid and auxin in the response of poplar to abiotic stress. Plant Biol. 2010, 12, 242–258. [Google Scholar] [CrossRef]
- Jensen, M.K.; Kjaersgaard, T.; Nielsen, M.M.; Galberg, P.; Petersen, K.; O’Shea, C.; Skriver, K. The Arabidopsis thaliana NAC transcription factor family: Structure–function relationships and determinants of ANAC019 stress signalling. Biochem. J. 2010, 426, 183–196. [Google Scholar] [CrossRef]
- Keshishian, E.A.; Rashotte, A.M. Plant cytokinin signalling. Essays Biochem. 2015, 58, 13–27. [Google Scholar] [CrossRef]
- Nieminen, K.; Immanen, J.; Laxell, M.; Kauppinen, L.; Tarkowski, P.; Dolezal, K.; Tähtiharju, S.; Elo, A.; Decourteix, M.; Ljung, K.; et al. Cytokinin signaling regulates cambial development in poplar. PNAS 2008, 105, 20032–20037. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Wildhagen, H.; Janz, D.; Teichmann, T.; Hänsch, R.; Polle, A. Tissue- and cell-specific cytokinin activity in Populus x canescens monitored by ARR5::GUS reporter lines in summer and winter. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Paul, S.; Wildhagen, H.; Janz, D.; Polle, A. Drought effects on the tissue- and cell-specific cytokinin activity in poplar. AoB Plants 2018, 10. [Google Scholar] [CrossRef]
- Jang, G.; Chang, S.H.; Um, T.Y.; Lee, S.; Kim, J.-K.; Choi, Y.D. Antagonistic interaction between jasmonic acid and cytokinin in xylem development. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Jang, G.; Choi, Y.D. Drought stress promotes xylem differentiation by modulating the interaction between cytokinin and jasmonic acid. Plant Signal. Behav. 2018, 13. [Google Scholar] [CrossRef]
- Brenner, W.G.; Ramireddy, E.; Heyl, A.; Schmülling, T. Gene regulation by cytokinin in Arabidopsis. Front Plant Sci. 2012, 3. [Google Scholar] [CrossRef]
- Nafisi, M.; Fimognari, L.; Sakuragi, Y. Interplays between the cell wall and phytohormones in interaction between plants and necrotrophic pathogens. Phytochemistry 2015, 112, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.W.; Oh, S.-I.; Kim, Y.Y.; Yoo, K.S.; Cui, M.H.; Shin, J.S. Arabidopsis histidine-containing phosphotransfer factor 4 (AHP4) negatively regulates secondary wall thickening of the anther endothecium during flowering. Mol. Cells 2008, 25, 294–300. [Google Scholar]
- Wilkinson, S.; Kudoyarova, G.R.; Veselov, D.S.; Arkhipova, T.N.; Davies, W.J. Plant hormone interactions: Innovative targets for crop breeding and management. J. Exp. Bot. 2012, 63, 3499–3509. [Google Scholar] [CrossRef] [PubMed]
- Reguera, M.; Peleg, Z.; Abdel-Tawab, Y.M.; Tumimbang, E.B.; Delatorre, C.A.; Blumwald, E. Stress-induced cytokinin synthesis increases drought tolerance through the coordinated regulation of carbon and nitrogen assimilation in rice. Plant Physiol. 2013, 163, 1609–1622. [Google Scholar] [CrossRef] [PubMed]
- Euring, D.; Bai, H.; Janz, D.; Polle, A. Nitrogen-driven stem elongation in poplar is linked with wood modification and gene clusters for stress, photosynthesis and cell wall formation. BMC Plant Biol. 2014, 14, 391. [Google Scholar] [CrossRef] [PubMed]
- Van den Broeck, H.C.; Maliepaard, C.; Ebskamp, M.J.M.; Toonen, M.A.J.; Koops, A.J. Differential expression of genes involved in C1 metabolism and lignin biosynthesis in wooden core and bast tissues of fibre hemp (Cannabis sativa L.). Plant Sci. 2008, 174, 205–220. [Google Scholar] [CrossRef]
- Camargo, E.L.O.; Nascimento, L.C.; Soler, M.; Salazar, M.M.; Lepikson Neto, J.; Marques, W.L.; Alves, A.; Teixeira, P.J.P.L.; Mieczkowski, P.; Carazzolle, M.F.; et al. Contrasting nitrogen fertilization treatments impact xylem gene expression and secondary cell wall lignification in Eucalyptus. BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Plavcová, L.; Hacke, U.G.; Almeida-Rodriguez, A.M.; Li, E.; Douglas, C.J. Gene expression patterns underlying changes in xylem structure and function in response to increased nitrogen availability in hybrid poplar. Plant Cell Environ. 2013, 36, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Zhou, J.; Li, H.; Shi, W.; Polle, A.; Lu, M.; Sun, X.; Luo, Z.-B. Global poplar root and leaf transcriptomes reveal links between growth and stress responses under nitrogen starvation and excess. Tree Physiol. 2015, 35, 1283–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, M.L.; Myers, B.J.; Raison, R.J. Dynamics of stem growth of Pinus radiata as affected by water and nitrogen supply. For. Ecol. Manag. 1992, 52, 117–137. [Google Scholar] [CrossRef]
- Dobbertin, M. Tree growth as indicator of tree vitality and of tree reaction to environmental stress: A review. Eur. J. For. Res. 2006, 125, 89. [Google Scholar] [CrossRef]
- Novaes, E.; Osorio, L.; Drost, D.R.; Miles, B.L.; Boaventura-Novaes, C.R.D.; Benedict, C.; Dervinis, C.; Yu, Q.; Sykes, R.; Davis, M.; et al. Quantitative genetic analysis of biomass and wood chemistry of Populus under different nitrogen levels. New Phytol. 2009, 182, 878–890. [Google Scholar] [CrossRef] [PubMed]
- Galle, A.; Esper, J.; Feller, U.; Ribas-Carbo, M.; Fonti, P. Responses of wood anatomy and carbon isotope composition of Quercus pubescens saplings subjected to two consecutive years of summer drought. Ann. For. Sci. 2010, 67, 809. [Google Scholar] [CrossRef]
- Liu, Z.; Dickmann, D.I. Abscisic acid accumulation in leaves of two contrasting hybrid poplar clones affected by nitrogen fertilization plus cyclic flooding and soil drying. Tree Physiol. 1992, 11, 109–122. [Google Scholar] [CrossRef]
- Liu, Z.; Dickmann, D.I. Responses of two hybrid Populus clones to flooding, drought, and nitrogen availability. I. Morphology and growth. Can. J. Bot. 1992, 70, 2265–2270. [Google Scholar] [CrossRef]
- Ibrahim, L.; Proe, M.F.; Cameron, A.D. Interactive effects of nitrogen and water availabilities on gas exchange and whole-plant carbon allocation in poplar. Tree Physiol. 1998, 18, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Mazzoleni, S.; Dickmann, D.I. Differential physiological and morphological responses of two hybrid Populus clones to water stress. Tree Physiol. 1988, 4, 61–70. [Google Scholar] [CrossRef]
- Harvey, H.P.; Van Den Driessche, R. Nutrition, xylem cavitation and drought resistance in hybrid poplar. Tree Physiol. 1997, 17, 647–654. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.H. Hydraulic architecture of some diffuse-porous trees. Can. J. Bot. 2011, 56, 2286–2295. [Google Scholar] [CrossRef]
- Euring, D. Nitrogen responsive wood formation in poplar (Populus sp.). In Chapter 3: N-Responsive Network; Georg-August-Universität Göttingen: Göttingen, Germany, 2014. [Google Scholar]
- Tarighaleslami, M.; Zarghami, R.; Mashhadi, M.; Boojar, A.; Oveysi, M. Effects of drought stress and different nitrogen levels on morphological traits of proline in leaf and protein of corn seed (Zea mays L.). Am.-Euras. J. Agric. Environ. Sci. 2012, 12, 49–56. [Google Scholar]
- Chen, S.; Polle, A. Salinity tolerance of Populus. Plant Biol. 2010, 12, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Polle, A.; Chen, S. On the salty side of life: Molecular, physiological and anatomical adaptation and acclimation of trees to extreme habitats. Plant Cell Environ. 2015, 38, 1794–1816. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Ottow, E.A.; Brinker, M.; Teichmann, T.; Fritz, E.; Kaiser, W.; Brosché, M.; Kangasjärvi, J.; Jiang, X.; Polle, A. Populus euphratica displays apoplastic sodium accumulation, osmotic adjustment by decreases in calcium and soluble carbohydrates, and develops leaf succulence under salt stress. Plant Physiol. 2005, 139, 1762–1772. [Google Scholar] [CrossRef]
- Lautner, S. Wood formation under drought stress and salinity. In Cellular Aspects of Wood Formation; Fromm, J., Ed.; Springer: Berlin, Germany, 2013; Volume 20, pp. 187–202. ISBN 978-3-642-36490-7. [Google Scholar]
- Escalante-Pérez, M.; Lautner, S.; Nehls, U.; Selle, A.; Teuber, M.; Schnitzler, J.-P.; Teichmann, T.; Fayyaz, P.; Hartung, W.; Polle, A.; et al. Salt stress affects xylem differentiation of grey poplar (Populus x canescens). Planta 2009, 229, 299–309. [Google Scholar] [CrossRef]
- Junghans, U.; Polle, A.; Düchting, P.; Weiler, E.; Kuhlman, B.; Gruber, F.; Teichmann, T. Adaptation to high salinity in poplar involves changes in xylem anatomy and auxin physiology. Plant Cell Environ. 2006, 29, 1519–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janz, D.; Lautner, S.; Wildhagen, H.; Behnke, K.; Schnitzler, J.-P.; Rennenberg, H.; Fromm, J.; Polle, A. Salt stress induces the formation of a novel type of ‘pressure wood’ in two Populus species. New Phytol. 2012, 194, 129–141. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S. Functional and ecological xylem anatomy. Perspect. Plant Ecol. Evol. Syst. 2001, 4, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Bhalerao, R.P.; Fischer, U. Environmental and hormonal control of cambial stem cell dynamics. J. Exp. Bot. 2017, 68, 79–87. [Google Scholar] [CrossRef]
- Uggla, C.; Moritz, T.; Sandberg, G.; Sundberg, B. Auxin as a positional signal in pattern formation in plants. PNAS 1996, 93, 9282–9286. [Google Scholar] [CrossRef]
- Uggla, C.; Mellerowicz, E.J.; Sundberg, B. Indole-3-Acetic Acid Controls Cambial Growth in Scots Pine by Positional Signaling. Plant Physiol. 1998, 117, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, J.; Karlberg, A.; Antti, H.; Lopez-Vernaza, M.; Mellerowicz, E.; Perrot-Rechenmann, C.; Sandberg, G.; Bhalerao, R.P. Dissecting the Molecular Basis of the Regulation of Wood Formation by Auxin in Hybrid Aspen. Plant Cell 2008, 20, 843–855. [Google Scholar] [CrossRef]
- Sheldrake, A.R. Effects of Osmotic Stress on Polar Auxin Transport in Avena Mesocotyl Sections. Planta 1979, 145, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Teichmann, T.; Bolu-Arianto, W.H.; Olbrich, A.; Langenfeld-Heyser, R.; Gobel, C.; Grzeganek, P.; Feussner, I.; Hansch, R.; Polle, A. GH3::GUS reflects cell-specific developmental patterns and stress-induced changes in wood anatomy in the poplar stem. Tree Physiol. 2008, 28, 1305–1315. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.E.; Kwon, Y.; Kim, J.H.; Noh, H.; Hong, S.-W.; Lee, H. A dual role for MYB60 in stomatal regulation and root growth of Arabidopsis thaliana under drought stress. Plant Mol. Biol. 2011, 77, 91–103. [Google Scholar] [CrossRef]
- Sixto, H.; González-González, B.D.; Molina-Rueda, J.J.; Garrido-Aranda, A.; Sanchez, M.M.; López, G.; Gallardo, F.; Cañellas, I.; Mounet, F.; Grima-Pettenati, J.; et al. Eucalyptus spp. and Populus spp. coping with salinity stress: An approach on growth, physiological and molecular features in the context of short rotation coppice (SRC). Trees 2016, 30, 1873–1891. [Google Scholar] [CrossRef]
- Guo, H.; Wang, Y.; Wang, L.; Hu, P.; Wang, Y.; Jia, Y.; Zhang, C.; Zhang, Y.; Zhang, Y.; Wang, C.; Yang, C. Expression of the MYB transcription factor gene BplMYB46 affects abiotic stress tolerance and secondary cell wall deposition in Betula platyphylla. Plant Biotechnol. J. 2017, 15, 107–121. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Function | Potri ID | Populus Gene Name | AGI ID | Arabidopsis Gene Name |
|---|---|---|---|---|
| Fine-tuning factors | Potri.001G188800 | Ptr-ATHB.12 | At1G52150 | AtHB15/AtCNA/AtICU4 |
| Potri.003G050100 | Ptr-ATHB.11 | At1G52150 | AtHB15/AtCNA/AtICU4 | |
| Potri.001G197000 | AT3G13890 | AtMYB26 | ||
| no annotated ortholog | At3G32090 | AtWRKY12 | ||
| Potri.002G023400 | Ptr-E2FC.1/E2Fc | At1G47870 | AtE2Fc | |
| Potri.001G061200 | PtrNAC053 | AT5G13180 | AtVNI2/AtANAC083 | |
| Master regulators (NAC) | no annotated ortholog | At1G32770 | AtNST3/AtSND1/AtANAC012 | |
| Potri.001G448400 | PtrWND1B/NAC063/PtVNS11 | At2G46770 | AtANAC043/AtNST1 | |
| Potri.002G178700 | PtrWND2B/NAC061/PtVNS10 | AT2G46770 | AtANAC043/AtNST1 | |
| Potri.011G153300 | PtrWND1A/NAC068/PtVNS12 | AT2G46770 | AtANAC043/AtNST1 | |
| Potri.014G104800 | PtrWND2A/NAC065/PtVNS09 | AT2G46770 | AtANAC043/AtNST1 | |
| Potri.015G127400 | PtrWND3A/NAC050/PtVNS05 | AT1G12260 | AtANAC007/AtNAC007/ATVND4 | |
| Potri.012G126500 | PtrWND3B/NAC037/PtVNS06 | AT1G12260 | AtANAC007/AtNAC007/ATVND4 | |
| Potri.001G120000 | PtrWND4A/NAC038/PtVNS03 | AT1G12260 | AtANAC007/AtNAC007/ATVND4 | |
| Potri.003G113000 | PtrWND4B/NAC046/PtVNS04 | AT1G12260 | AtANAC007/AtNAC007/ATVND4 | |
| Potri.007G014400 | PtrWND5A/NAC025/PtVNS01 | AT2G18060 | AtANAC037/AtVND1 | |
| Potri.005G116800 | PtrWND5B/NAC039/PtVNS02 | AT2G18060 | AtANAC037/AtVND1 | |
| Potri.013G113100 | PtrWND6A/NAC055/PtVNS07 | AT1G71930 | AtANAC030/AtVND7 | |
| Potri.019G083600 | PtrWND6B/NAC060/PtVNS08 | AT1G71930 | AtANAC030/AtVND7 | |
| Potri.015G002900 | PtrNAC147 | AT1G71930 | AtANAC030/AtVND7 | |
| no annotated ortholog | At3G61910 | AtNST2 | ||
| Potri.004G107200 | At5G62380 | AtVND6 | ||
| Potri.004G107400 | At5G62380 | AtVND6 | ||
| Potri.005G082700 | PtrNAC144 | At5G62380 | AtVND6 | |
| Potri.006G231300 | At5G62380 | AtVND6 | ||
| Potri.014G163600 | At5G62380 | AtVND6 | ||
| no annotated ortholog | At4G36160 | AtVND2/AtANAC076 | ||
| no annotated ortholog | At5G66300 | AtVND3/AtANAC105 | ||
| no annotated ortholog | At1G62700 | AtVND5 | ||
| Second Level Regulators (MYB) | Potri.001G258700 | PtrMYB2 | At5G12870 | AtMYB46 |
| Potri.009G053900 | PtrMYB21 | At5G12870 | AtMYB46 | |
| Potri.001G267300 | At3G08500 | AtMYB83 | ||
| Potri.009G061500 | At3G08500 | AtMYB83 | ||
| Third Level Regulators | Potri.004G049300 | AT4G28500 | AtSND2/AtANAC073 | |
| Potri.007G135300 | AT4G28500 | AtSND2/AtANAC073 | ||
| Potri.011G058400 | AT4G28500 | AtSND2/AtANAC073 | ||
| Potri.017G016700 | AT4G28500 | AtSND2/AtANAC073 | ||
| no annotated ortholog | AT1G28470 | AtSND3 | ||
| no annotated ortholog | At1G63910 | AtMYB103 | ||
| Potri.002G073500 | At1G17950 | AtMYB52 | ||
| Potri.005G186400 | PtrMYB158/PtrMYB.50 | At1G17950 | AtMYB52 | |
| Potri.007G134500 | PtrMYB161/PtrMYB.43 | At1G17950 | AtMYB52 | |
| Potri.012G039400 | PtrMYB167/PtrMYB.41 | At1G17950 | AtMYB52 | |
| Potri.015G033600 | PtrMYB090/PtrMYB.38 | At1G17950 | AtMYB52 | |
| no annotated ortholog | At3G48920 | AtMYB45 | ||
| no annotated ortholog | At1G16490 | AtMYB58 | ||
| Potri.005G096600 | At1G79180 | AtMYB63 | ||
| Potri.007G067600 | At1G79180 | AtMYB63 | ||
| Potri.019G118900 | At1G79180 | AtMYB63 | ||
| Potri.015G129100 | At4G22680 | AtMYB85 | ||
| Potri.001G112200 | PtrKNAT7.1 | At1G62990 | AtKNAT7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eckert, C.; Sharmin, S.; Kogel, A.; Yu, D.; Kins, L.; Strijkstra, G.-J.; Polle, A. What Makes the Wood? Exploring the Molecular Mechanisms of Xylem Acclimation in Hardwoods to an Ever-Changing Environment. Forests 2019, 10, 358. https://doi.org/10.3390/f10040358
Eckert C, Sharmin S, Kogel A, Yu D, Kins L, Strijkstra G-J, Polle A. What Makes the Wood? Exploring the Molecular Mechanisms of Xylem Acclimation in Hardwoods to an Ever-Changing Environment. Forests. 2019; 10(4):358. https://doi.org/10.3390/f10040358
Chicago/Turabian StyleEckert, Christian, Shayla Sharmin, Aileen Kogel, Dade Yu, Lisa Kins, Gerrit-Jan Strijkstra, and Andrea Polle. 2019. "What Makes the Wood? Exploring the Molecular Mechanisms of Xylem Acclimation in Hardwoods to an Ever-Changing Environment" Forests 10, no. 4: 358. https://doi.org/10.3390/f10040358