Growth, Biomass Production and Root Development of Chinese fir in Relation to Initial Planting Density

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
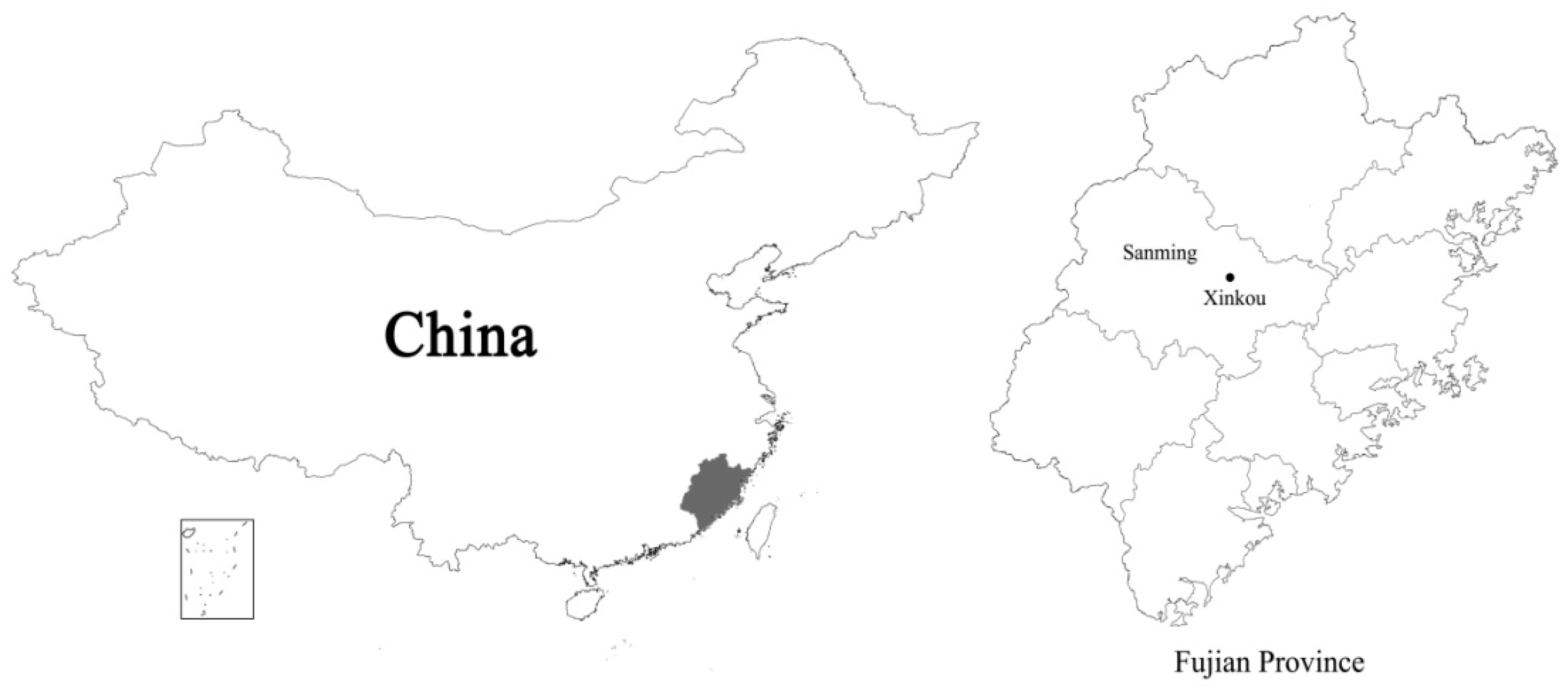
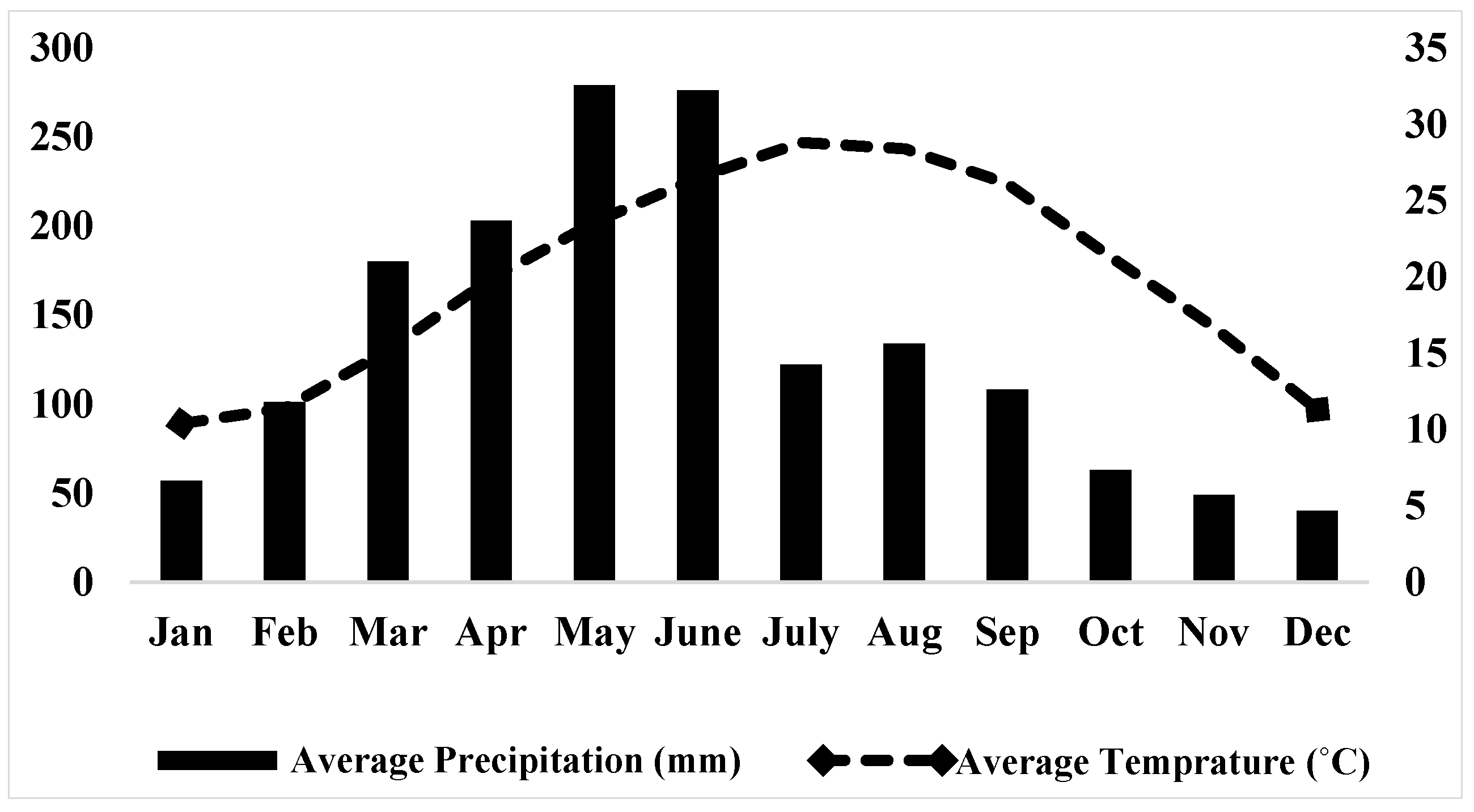
2.1. Study Site
2.2. Determination of Biomass Production
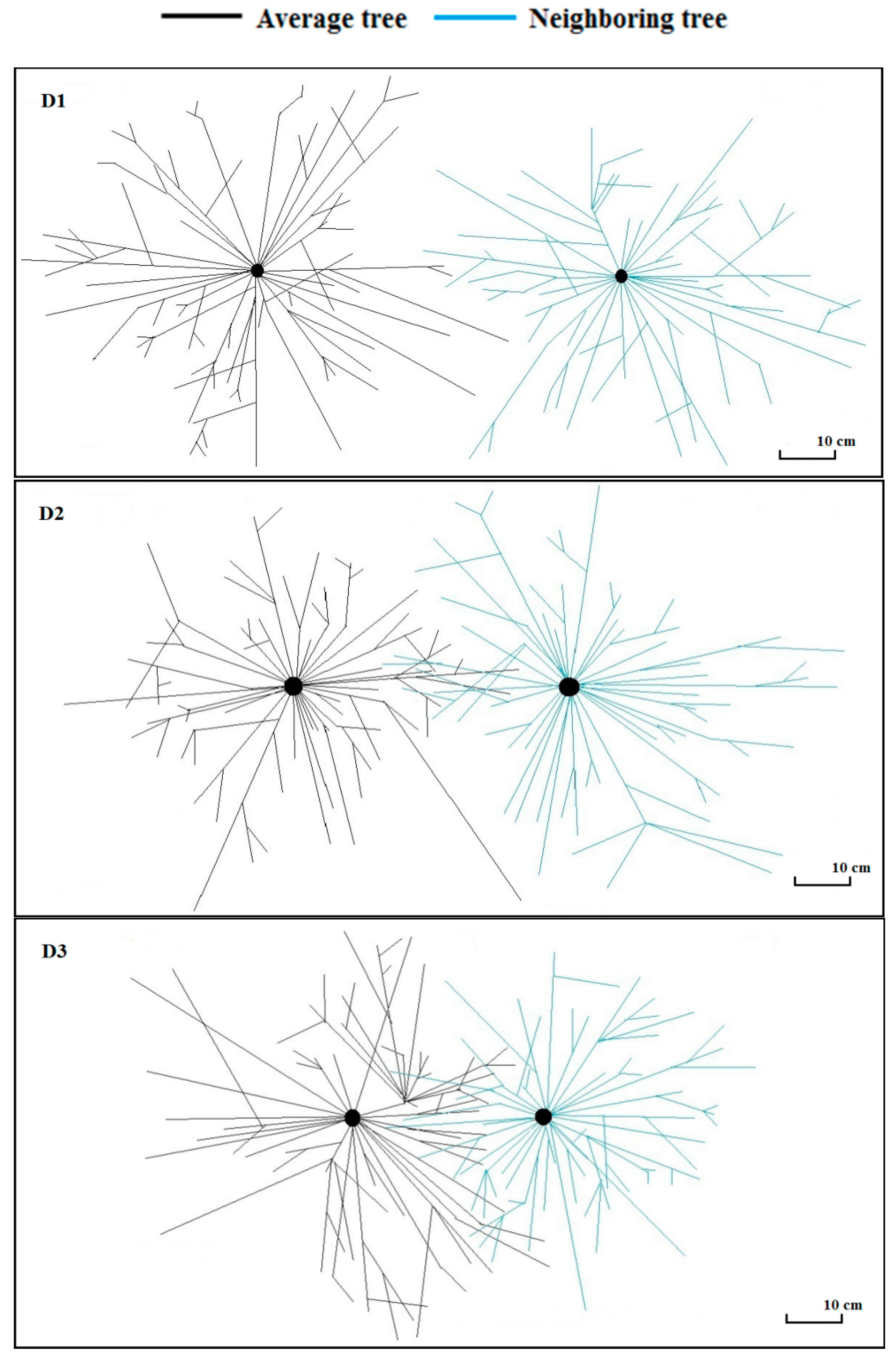
2.3. Spatial Distribution of Root Morphological Traits
2.4. Data Analysis
3. Results
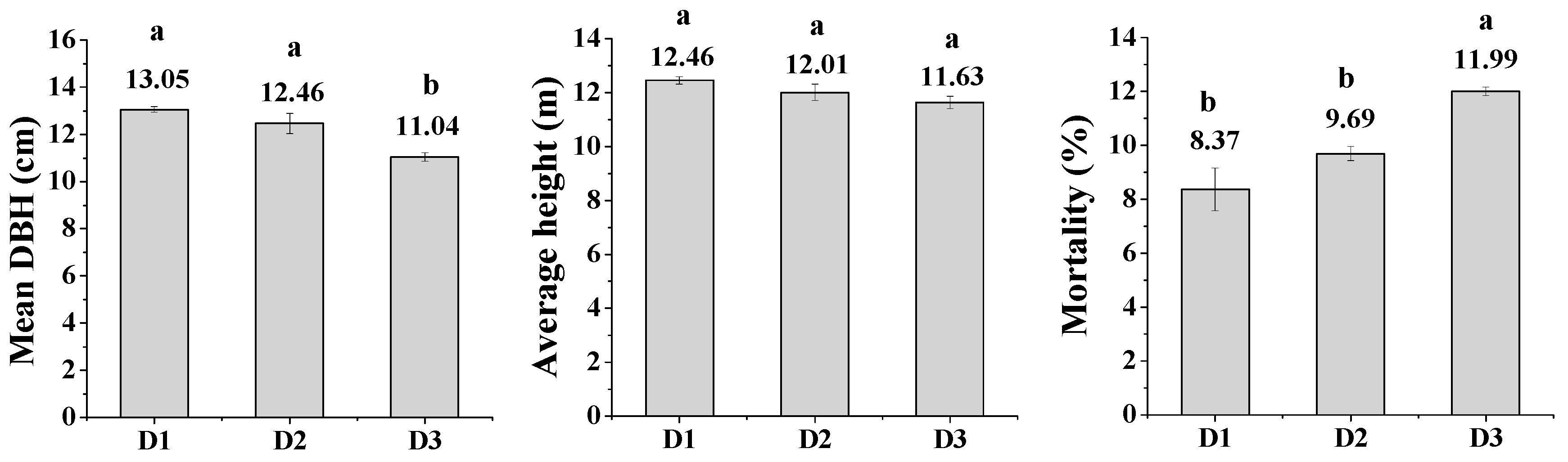
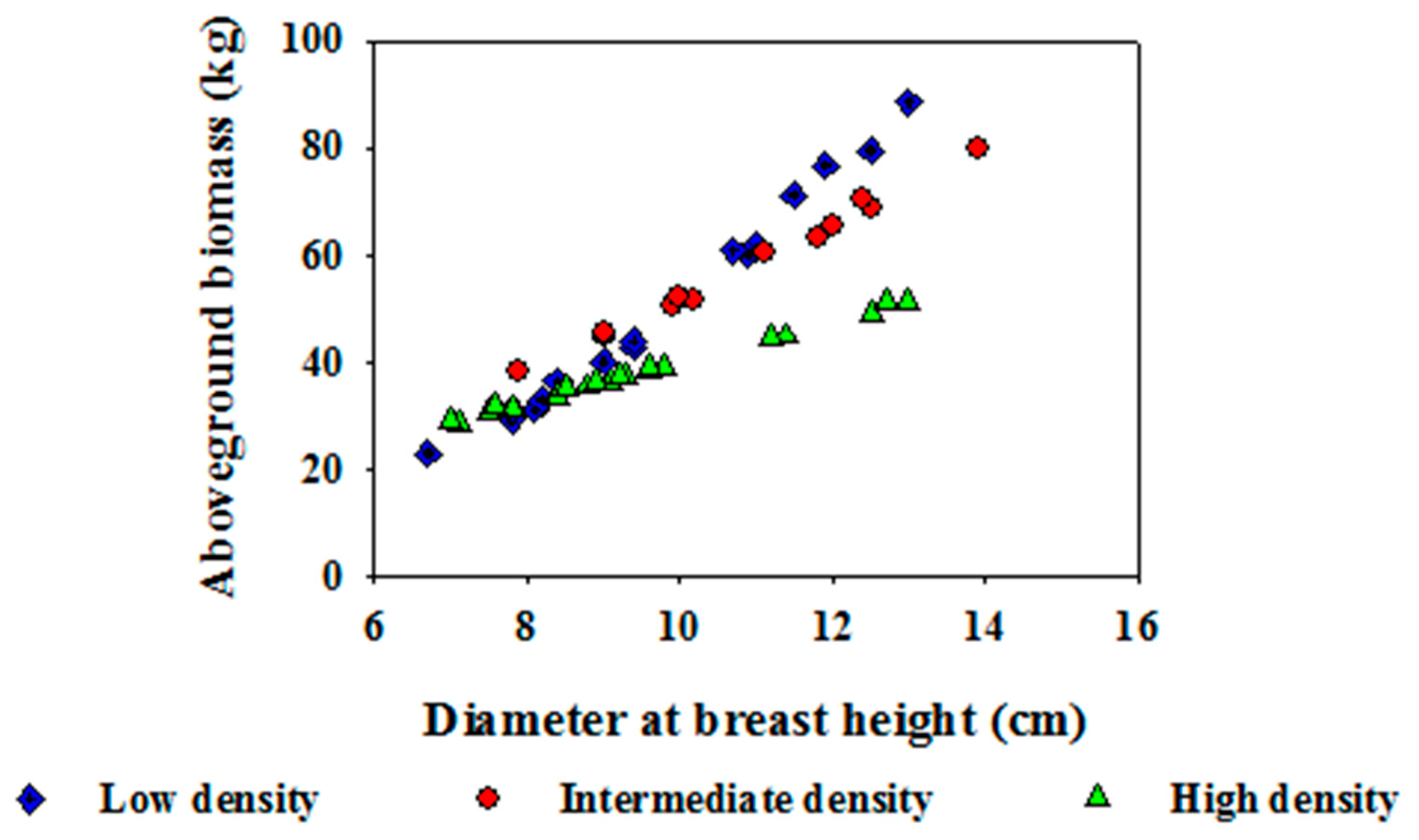
3.1. Tree Growth and Biomass Production
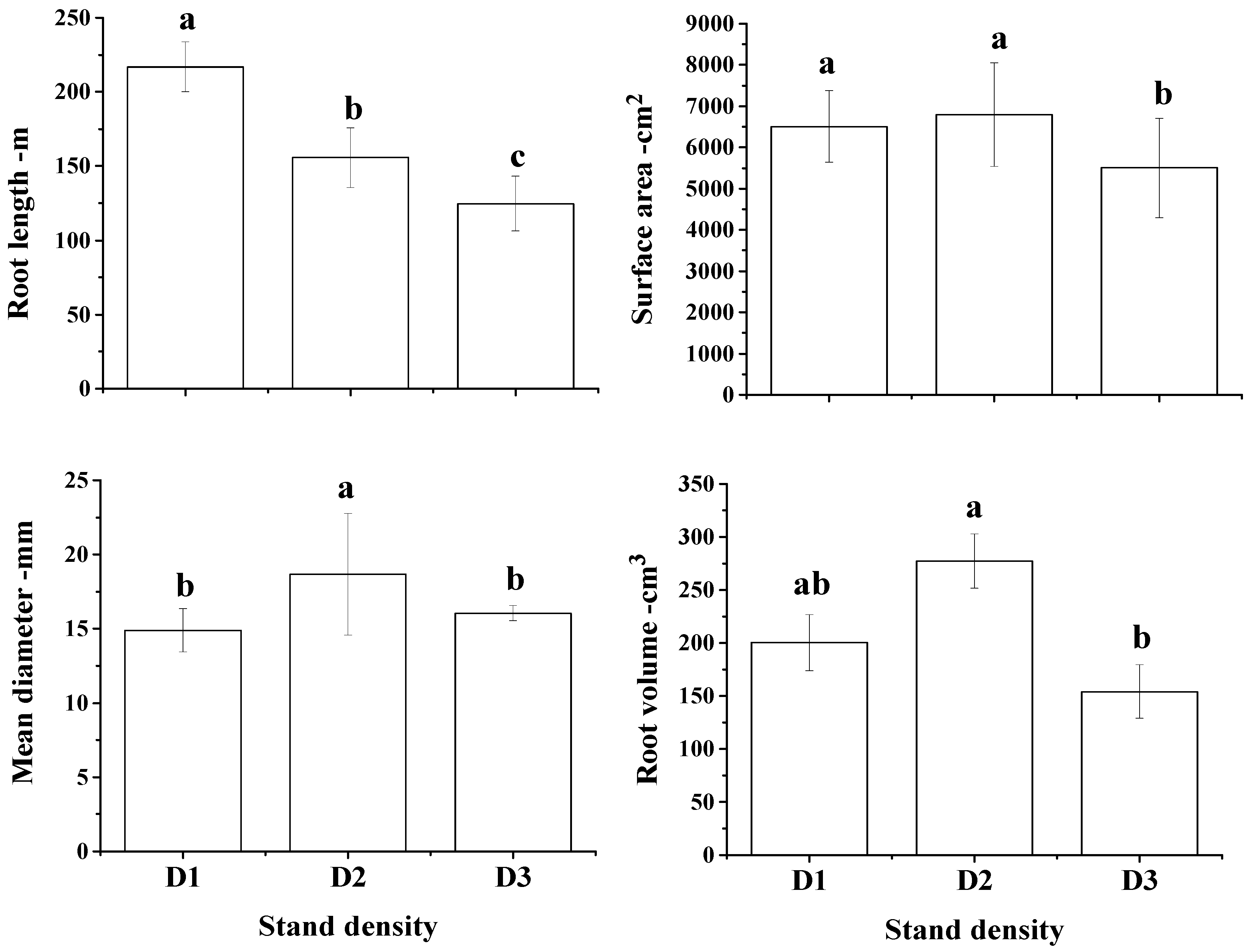
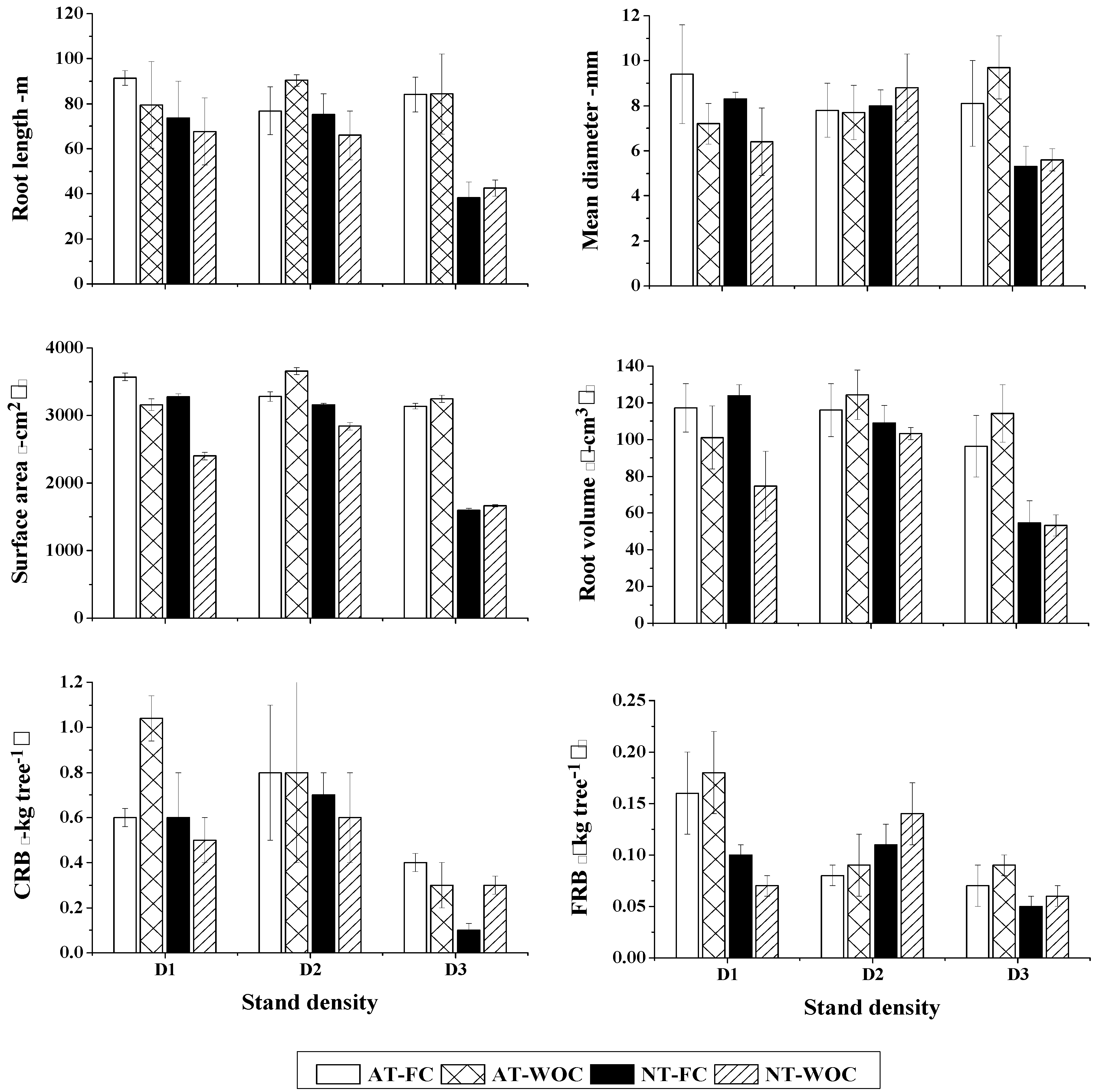
3.2. Root Morphological Traits and Their Spatial Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wu, Z.L. Chinese Fir; Forestry of China Publications: Beijing, China, 1984. [Google Scholar]
- Yu, X. Silviculture of Chinese-Fir; Science and Technology Press of Fujian: Fuzhou, China, 1997. [Google Scholar]
- Yao, L.; Kang, W.; Zhao, Z.; He, J. Carbon fixed characteristics of plant of Chinese fir (Cunninghamia lanceolata) plantation at different growth stages in Huitong. Acta Ecol. Sin. 2015, 35, 1–16. [Google Scholar]
- FAO. State of the World’s Forests; Food and Agriculture Organization of the United Nations: Rome, Italy, 2007; pp. 88–90. [Google Scholar]
- SFA (State Forestry Administration of Forest Resources Management). The 7th national forest resources inventory and the status of forest resources. For. Resour. Manag. 2010, 1, 1–8. [Google Scholar]
- FAO. Global Forest Resources Assessment. Rome 2015. Available online: http://www.fao.org/forest-resources-assessment/en/ (accessed on 20 December 2018).
- Ma, X.; Liu, C.; Hannu, I.; Westman, C.J.; Liu, A. Biomass, litterfall and the nutrient fluxes in Chinese fir stands of different age in subtropical China. J. For. Res. 2002, 13, 165–170. [Google Scholar] [CrossRef]
- Ma, X. Advance in researches on productivity decline of replanting Chinese-fir forests. J. Fujian For. Coll. 2001, 21, 380–384. [Google Scholar]
- Ding, Y.; Tian, Y.; Qi, L. A testing simulation with FORECAST on long-term productivity of Chinese-fir plantations. For. Stud. China 1999, 1, 34–38. [Google Scholar]
- Wu, P.; Wang, G.; Farooq, T.H.; Li, Q.; Zou, X.; Ma, X. Low phosphorus and competition affect Chinese fir cutting growth and root organic acid content: Does neighboring root activity aggravate P nutrient deficiency? J. Soils Sediments 2017, 17, 2775–2785. [Google Scholar] [CrossRef]
- Yang, Y.; Cheng, G.; Huang, B. Variation in soil water and nutrients between different rotation stands of Chinese fir. J. Nanjing For. Univ. 2000, 24, 25–28. [Google Scholar]
- Dong, T.; Zhang, Y.; Zhang, Y.; Zhang, S. Continuous planting under a high density enhances the competition for nutrients among young Cunninghamia lanceolata saplings. Ann. For. Sci. 2015, 73, 331–339. [Google Scholar] [CrossRef]
- Tian, D. A long-term evaluation of biomass production in first and second rotations of Chinese fir plantations at the same site. Forestry 2011, 84, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Xiang, W.; Tian, D.; Deng, X.; Huang, Z.; Zhou, X.; Peng, C. Effects of increased nitrogen deposition and rotation length on long-term productivity of Cunninghamia lanceolata plantation in southern China. PLoS ONE 2013, 8, e55376. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Heal, K.V.; Liu, A.; Jarvis, P.G. Nutrient cycling and distribution in different-aged plantations of Chinese fir in southern China. For. Ecol. Manag. 2007, 243, 61–74. [Google Scholar] [CrossRef]
- Tun, T.N.; Guo, J.; Fang, S.; Tian, Y. Planting spacing affects canopy structure, biomass production and stem roundness in poplar plantations. Scand. J. For. Res. 2018, 33, 464–474. [Google Scholar] [CrossRef]
- Liziniewicz, M.; Ekö, P.M.; Agestam, E. Effect of spacing on 23-year-old lodgepole pine (Pinus contorta Dougl. var. latifolia) in southern Sweden. Scand. J. For. Res. 2010, 27, 361–371. [Google Scholar] [CrossRef]
- Nagar, B.; Rawat, S.; Rathiesh, P.; Sekar, I. Impact of initial spacing on growth and yield of Eucalyptus camaldulensis in arid region of India. World Appl. Sci. J. 2015, 33, 1362–1368. [Google Scholar]
- Ciancio, O.; Garfì, V.; Marziliano, P.A.; Menguzzato, G.; Pelle, L. Effects of plant ing densitiy on douglas fir stands. Ital. J. For. Mt. Environ. 2008, 59, 519–534. [Google Scholar]
- La Marca, O.; Marziliano, P.A.; Moretti, N. Experimental research in ageing holm oak (Quercus ilex L.) coppices: Preliminary results. Ann. For. Sci. 1998, 4, 461–476. [Google Scholar] [CrossRef]
- Harrington, T.B.; Harrington, C.A.; DeBell, D.S. Effects of planting spacing and site quality on 25-year growth and mortality relationships of Douglas-fir (Pseudotsuga menziesiivar.menziesii). For. Ecol. Manag. 2009, 258, 18–25. [Google Scholar] [CrossRef]
- Zeide, B. How to measure stand density. Trees 2005, 19, 1–14. [Google Scholar] [CrossRef]
- Xue, L.; Pan, L.; Zhang, R.; Xu, P.B. Density effects on the growth of self-thinning Eucalyptus urophylla stands. Trees 2011, 25, 1021–1031. [Google Scholar] [CrossRef]
- Sprintsin, M.; Karnieli, A.; Sprintsin, S.; Cohen, S.; Berliner, P. Relationships between stand density and canopy structure in a dryland forest as estimated by ground-based measurements and multi-spectral spaceborne images. J. Arid Environ. 2009, 73, 955–962. [Google Scholar] [CrossRef]
- Weiner, J. Asymmetric competition in plant populations. Tree 1990, 5, 360–364. [Google Scholar] [CrossRef]
- Manning, P.; Houston, K.; Evans, T. Shifts in seed size across experimental nitrogen enrichment and plant density gradients. Basic Appl. Ecol. 2009, 10, 300–308. [Google Scholar] [CrossRef]
- Berger, U.; Piou, C.; Schiffers, K.; Grimm, V. Competition among plants: Concepts, individual-based modeling approaches, and a proposal for a future research strategy. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 121–135. [Google Scholar] [CrossRef]
- Hunt, M.A.; Battaglia, M.; Davidson, N.J.; Unwin, G.L. Competition between plantation Eucalyptus nitens and Acacia dealbata weeds in northeastern Tasmania. For. Ecol. Manag. 2006, 233, 260–274. [Google Scholar] [CrossRef]
- Fetene, M. Intra- and inter-specific competition between seedlings of Acacia etbaicaand a perennial grass (Hyparrenia hirta). J. Arid Environ. 2003, 55, 441–451. [Google Scholar] [CrossRef]
- Zhou, L.; Shalom, A.-D.D.; Wu, P.; He, Z.; Liu, C.; Ma, X. Biomass production, nutrient cycling and distribution in age-sequence Chinese fir (Cunninghamia lanceolata) plantations in subtropical China. J. For. Res. 2016, 27, 357–368. [Google Scholar] [CrossRef]
- Zhao, M.; Zhou, G.-S. Estimation of biomass and net primary productivity of major planted forests in China based on forest inventory data. For. Ecol. Manag. 2005, 207, 295–313. [Google Scholar] [CrossRef]
- Wu, P.; Tigabu, M.; Ma, X.; Odén, P.C.; He, Y.; Yu, X.; He, Z. Variations in biomass, nutrient contents and nutrient use efficiency among Chinese fir provenances. Silvae Genet. 2011, 60, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, C.; Webster, A. The influence of the development of temperate fruit tree species on the potential for their uptake of radionuclides. J. Environ. Radioact. 2001, 52, 131–146. [Google Scholar] [CrossRef]
- Marziliano, P.A.; Coletta, V.; Menguzzato, G.; Nicolaci, A.; Pellicone, G.; Veltri, A. Effects of planting density on the distribution of biomass in a douglas-fir plantation in southern Italy. Iforest 2015, 8, 368. [Google Scholar] [CrossRef]
- USDA Soil Survey Staff. Soil Taxonomy; Agriculture Handbook 436; USDA: Washington, DC, USA, 1999.
- Sanesi, G.; Lafortezza, R.; Marziliano, P.A.; Ragazzi, A.; Mariani, L. Assessing the current status of urban forest resources in the context of Parco Nord, Milan, Italy. Landsc. Ecol. Eng. 2007, 3, 187–198. [Google Scholar] [CrossRef]
- Corona, P.; Marziliano, P.A.; Scotti, R. Top-down growth modelling: A prototype for poplar plantations in Italy. For. Ecol. Manag. 2002, 161, 65–73. [Google Scholar] [CrossRef]
- Nissen, T.; Midmore, D.; Keeler, A. Biophysical and economic tradeoffs of intercropping timber with food crops in the Philippine uplands. Agric. Syst. 2001, 67, 49–69. [Google Scholar] [CrossRef]
- Aref, I.; El-Juhany, L.; Nasroon, T. Pattern of above-ground biomass production and allocation in Leucaena leucocephala trees when planted at different spacing. Saudi J. Biol. Sci. 1999, 6, 27–34. [Google Scholar]
- McEvoy, T.J. Positive Impact Forestry: A Sustainable Approach to Managing Woodlands; Island Press: Washington, DC, USA, 2012; pp. 1–268. [Google Scholar]
- Cardoso, D.J.; Lacerda, A.E.B.; Rosot, M.A.D.; Garrastazú, M.C.; Lima, R.T. Influence of spacing regimes on the development of loblolly pine (Pinus taeda L.) in Southern Brazil. For. Ecol. Manag. 2013, 310, 761–769. [Google Scholar] [CrossRef]
- Bernardo, A.L.; Reis, M.G.F.; Reis, G.G.; Harrison, R.B.; Firme, D.J. Effect of spacing on growth and biomass distribution in Eucalyptus camaldulensis, E. pellitaand E. urophylla plantations in southeastern Brazil. For. Ecol. Manag. 1998, 104, 1–13. [Google Scholar] [CrossRef]
- Pinkard, E.; Neilsen, W. Crown and stand characteristics of Eucalyptus nitens in response to initial spacing: Implications for thinning. For. Ecol. Manag. 2003, 172, 215–227. [Google Scholar] [CrossRef]
- Alcorn, P.J.; Pyttel, P.; Bauhus, J.; Smith, R.G.B.; Thomas, D.; James, R.; Nicotra, A. Effects of initial planting density on branch development in 4-year-old plantation grown Eucalyptus pilularis and Eucalyptus cloeziana trees. For. Ecol. Manag. 2007, 252, 41–51. [Google Scholar] [CrossRef]
- Wang, C. Effect of planting density on knot attributes and branch occlusion of Betula alnoides under natural pruning in southern China. Forests 2015, 6, 1343–1361. [Google Scholar] [CrossRef]
- Neilsen, W.A.; Gerrand, A.M. Growth and branching habit of Eucalyptus nitens at different spacing and the effect on final crop selection. For. Ecol. Manag. 1999, 123, 217–229. [Google Scholar] [CrossRef]
- Kearney, D. Characterisation of Branching Patterns, Changes Caused by Variations in Initial Stocking and Implications for Silviculture, for E. grandis and E. pilularis Plantations in the North Coast Region of NSW. Honours Thesis, Department of Forestry, The Australian National University, Canberra, Australia, 1999; p. 89. [Google Scholar]
- Khan, G.; Chaudhry, A.K. Effect of spacing and plant density on the growth of poplar (Populus deltoides) trees under agro-forestry system. Pak. J. Agric. Sci. 2007, 44, 321–327. [Google Scholar]
- Lanner, R.M. On the insensitivity of height growth to spacing. For. Ecol. Manag. 1985, 13, 143–148. [Google Scholar] [CrossRef]
- Seidel, K.W. A Western Larch-Engelmann Spruce Spacing Study in Eastern Oregon: Results after 10 Years; USDA For Serv Res Note PNW-409; US Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1984.
- Ritchie, G.A. Evidence for red: Far red signaling and photomorphogenic response in Douglas-fir (Pseudotsuga menziesii) seedlings. Tree Physiol. 1997, 17, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Knowe, S.A.; Hibbs, D.E. Stand structure of young red alder as affected by plant density. For. Ecol. Manag. 1996, 82, 69–85. [Google Scholar] [CrossRef]
- Gilbert, I.R.; Seavers, G.P.; Jarvis, P.G.; Smith, H. Photomorphogenesis and canopy dynamics. Phytochrome-mediated proximity perception accounts for the growth dynamics of canopies of Populus trichocarpa × deltoids ‘Beaupre’. Plant Cell Environ. 1995, 18, 475–497. [Google Scholar] [CrossRef]
- Giordano, P.A.; Hibbs, D.E. Morphological response to competition in red alder: The role of water. Funct. Ecol. 1993, 7, 462–468. [Google Scholar] [CrossRef]
- Pienaar, L.V.; Shiver, B.D. Early results from an old-field loblolly pine spacing study in the Georgia Piedmont with competition control. South J. Appl. For. 1993, 17, 193–196. [Google Scholar]
- Toillon, J.; Fichot, R.; Dallé, E.; Berthelot, A.; Brignolas, F.; Marron, N. Planting density affects growth and water-use efficiency depending on site in Populus deltoides × P. nigra. For. Ecol. Manag. 2013, 304, 345–354. [Google Scholar] [CrossRef]
- Dickens, E.D.; Barnett, J.P.; Hubbard, W.G.; Jokela, E.J. Slash Pine: Still Growing and Growing! Gen. Tech. Rep. SRS-76; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2004; p. 148.
- Zhang, J.; Ritchie, M.W.; Maguire, D.A.; Oliver, W.W. Thinning ponderosa pine (Pinus ponderosa) stands reduces mortality while maintaining stand productivity. Can. J. For. Res. 2013, 43, 311–320. [Google Scholar] [CrossRef]
- Newton, P.; Jolliffe, P. Aboveground modular component responses to intraspecific competition within density-stressed black spruce stands. Can. J. For. Res. 1998, 28, 1587–1610. [Google Scholar] [CrossRef]
- Stokes, A.; Atger, C.; Bengough, A.G.; Fourcaud, T.; Sidle, R.C. Desirable plant root traits for protecting natural and engineered slopes against landslides. Plant Soil 2009, 324, 1–30. [Google Scholar] [CrossRef]
- Mandal, K.; Hati, K.; Misra, A.; Bandyopadhyay, K. Root biomass, crop response and water-yield relationship of mustard (Brassica juncea L.) grown under combinations of irrigation and nutrient application. Irrig. Sci. 2010, 28, 271–280. [Google Scholar] [CrossRef]
- Douglas, G.B.; McIvor, I.R.; Potter, J.F.; Foote, L.G. Root distribution of poplar at varying densities on pastoral hill country. Plant Soil 2010, 333, 147–161. [Google Scholar] [CrossRef]
- Liu, L.; Gan, Y.; Bueckert, R.; Van Rees, K. Rooting systems of oilseed and pulse crops. II: Vertical distribution patterns across the soil profile. Field Crops Res. 2010, 122, 248–255. [Google Scholar] [CrossRef]
- Loades, K.W.; Bengough, A.G.; Bransby, M.F.; Hallett, P.D. Planting density influence on fibrous root reinforcement of soils. Ecol. Eng. 2010, 36, 276–284. [Google Scholar] [CrossRef]
- Van Noordwijk, M.; Lawson, G.; Hairiah, K.; Wilson, J. Root Distribution of Trees and Crops: Competition and/or Complementarity Tree–Crop Interactions: Agroforestry in a Changing Climate; CABI: Wallingford, UK, 2015; pp. 221–257. [Google Scholar]
- Litton, C.M.; Ryan, M.G.; Tinker, D.B.; Knight, D.H. Belowground and aboveground biomass in young postfire lodgepole pine forests of contrasting tree density. Can. J. For. Res. 2003, 33, 351–363. [Google Scholar] [CrossRef]
- Chang, R.; Fu, B.; Liu, G.; Yao, X.; Wang, S. Effects of soil physicochemical properties and stand age on fine root biomass and vertical distribution of plantation forests in the Loess Plateau of China. Ecol. Res. 2012, 27, 827–836. [Google Scholar] [CrossRef] [Green Version]
- McIvor, I.; Douglas, G.; Hurst, S.; Hussain, Z.; Foote, A. Structural root growth of young Veronese poplars on erodible slopes in the southern North Island, New Zealand. Agrofor. Syst. 2008, 72, 75–86. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Density | Tree Components | Biomass Equations | R2 |
|---|---|---|---|
| Low | Live Branches | W = 0.0087 (D2H)0.556 | 0.972 |
| Live Needles | W = 0.0021 (D2H)1.324 | 0.911 | |
| Bark | W = 0.0986 (D2H)0.425 | 0.836 | |
| Stem | W = 0.0242 (D2H)0.983 | 0.879 | |
| Roots | W = 0.0049 (D2H)0.793 | 0.982 | |
| Dead Branches | W = 0.0025 (D2H) − 0.003 | 0.297 | |
| Dead Needles | W = 0.0004 (D2H) + 1.552 | 0.301 | |
| Intermediate | Live Branches | W = 0.0652 (D2H)0.513 | 0.923 |
| Live Needles | W = 0.0263 (D2H)0.303 | 0.897 | |
| Bark | W = 0.0269 (D2H)0.331 | 0.898 | |
| Stem | W = 0.0496 (D2H)0.918 | 0.995 | |
| Roots | W = 0.0846 (D2H)0.035 | 0.932 | |
| Dead Branches | W = 0.0026 (D2H) + 7.208 | 0.347 | |
| Dead Needles | W = 0.0024 (D2H) + 6.551 | 0.350 | |
| High | Live Branches | W = 0.015 (D2H)0.192 | 0.896 |
| Live Needles | W = 0.019 (D2H)0.179 | 0.938 | |
| Bark | W = 0.1442 (D2H)0.487 | 0.997 | |
| Stem | W = 1.4003 (D2H)0.450 | 0.915 | |
| Roots | W = 0.0916 (D2H)0.013 | 0.996 | |
| Dead Branches | W = 0.0015 (D2H) + 0.678 | 0.463 | |
| Dead Needles | W = 0.0009 (D2H) + 0.384 | 0.368 |
| Tree Components | Individual Tree Biomass (kg tree−1) | Stand Level Biomass (t ha−1) | ||||
|---|---|---|---|---|---|---|
| D1 | D2 | D3 | D1 | D2 | D3 | |
| Branches | 2.55 ± 0.28 A | 2.59 ± 0.30 A | 2.53 ± 0.31 A | 1.68 ± 0.31 b | 2.85 ± 0.33 b | 4.47 ± 0.56 a |
| Needles | 3.82 ± 0.49 B | 6.11 ± 0.44 A | 3.34 ± 0.29 B | 2.52 ± 0.46 b | 6.74 ± 0.49 a | 5.90 ± 0.51 a |
| Dead branches | 0.76 ± 0.22 AB | 1.07 ± 0.59 A | 0.39 ± 0.23 B | 0.04 ± 0.01 b | 1.34 ± 0.32 a | 0.94 ± 0.19 a |
| Dead needles | 1.22 ± 0.26 B | 1.89 ± 0.31 A | 0.79 ± 0.18 C | 0.02 ± 0.05 c | 1.93 ± 0.35 a | 1.14 ± 0.21 b |
| Bark | 3.15 ± 0.25 AB | 4.38 ± 0.34 A | 2.29 ± 0.16 B | 2.08 ± 0.30 b | 4.83 ± 0.38 a | 4.05 ± 0.28 a |
| Stem wood | 24.84 ± 2.51 B | 35.43 ± 2.32 A | 17.19 ± 1.67 C | 16.43 ± 1.66 b | 39.05 ± 2.66 a | 30.32 ± 1.19 a |
| Total agb * | 36.34 ± 3.75 B | 51.48 ± 3.99 A | 26.53 ± 2.66 C | 22.73 ± 2.74 b | 56.74 ± 4.51 a | 46.82 ± 2.94 a |
| Stump root | 6.78 ± 1.0 AB | 8.19 ± 0.61 A | 5.10 ± 0.22 B | 7.48 ± 0.66 b | 9.03 ± 0.21 a | 8.99 ± 0.39 a |
| Coarse root | 1.38 ± 0.19 A | 1.47 ± 0.23 A | 0.57 ± 0.08 B | 0.03 ± 0.01 b | 1.63 ± 0.37 a | 1.01 ± 0.15 a |
| Fine root | 0.22 ± 0.02 A | 0.21 ± 0.04 A | 0.15 ± 0.02 A | 0.15 ± 0.01 b | 0.23 ± 0.05 a | 0.26 ± 0.04 a |
| Total bgb ** | 8.38 ± 1.21 AB | 9.87 ± 0.88 A | 5.82 ± 0.31 B | 7.66 ± 0.68 b | 10.89 ± 0.63 a | 10.26 ± 0.19 a |
| Total biomass | 44.72 ± 4.96 B | 61.35 ± 4.78 A | 32.35 ± 2.98 C | 27.39 ± 3.42 b | 67.63 ± 4.14 a | 57.08 ± 3.13 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq, T.H.; Wu, W.; Tigabu, M.; Ma, X.; He, Z.; Rashid, M.H.U.; Gilani, M.M.; Wu, P. Growth, Biomass Production and Root Development of Chinese fir in Relation to Initial Planting Density. Forests 2019, 10, 236. https://doi.org/10.3390/f10030236
Farooq TH, Wu W, Tigabu M, Ma X, He Z, Rashid MHU, Gilani MM, Wu P. Growth, Biomass Production and Root Development of Chinese fir in Relation to Initial Planting Density. Forests. 2019; 10(3):236. https://doi.org/10.3390/f10030236
Chicago/Turabian StyleFarooq, Taimoor Hassan, Wenjing Wu, Mulualem Tigabu, Xiangqing Ma, Zongming He, Muhammad Haroon U. Rashid, Matoor Mohsin Gilani, and Pengfei Wu. 2019. "Growth, Biomass Production and Root Development of Chinese fir in Relation to Initial Planting Density" Forests 10, no. 3: 236. https://doi.org/10.3390/f10030236